
Exogenous Application of Indol-3-Acetic Acid and Salicylic Acid Improves Tolerance to Salt Stress in Olive Plantlets (Olea europaea L. Cultivar Picual) in Growth Chamber Environments



Abstract
:1. Introduction
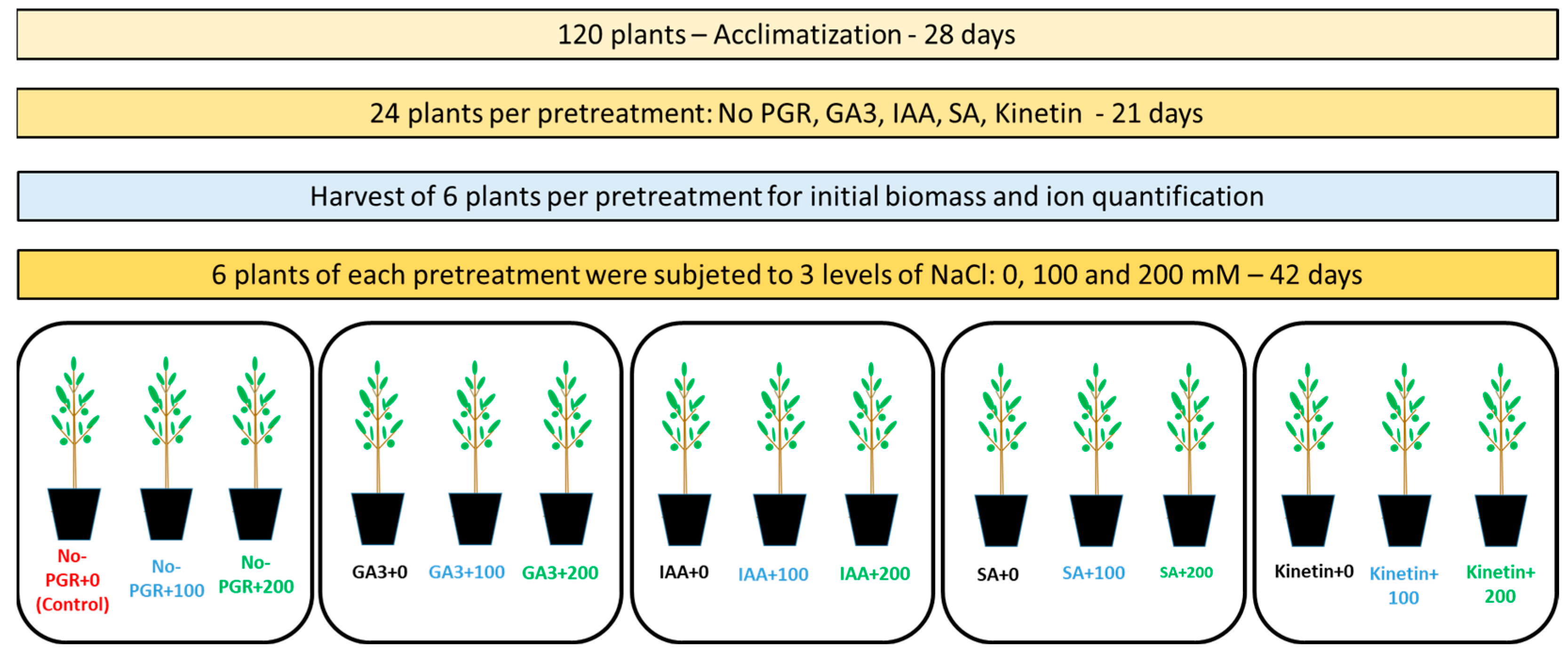
2. Materials and Methods
2.1. Plant Material and Growth Conditions
- -
- Group 1: each plant was sprayed with 25 mL of distilled water (no PGR pretreatment).
- -
- Group 2: each plant was sprayed with 25 mL of gibberellic acid (GA3) (1 µM).
- -
- Group 3: each plant was sprayed with 25 mL of indol-3-acetic acid (IAA) (1 µM).
- -
- Group 4: each plant was sprayed with 25 mL of salicylic acid (SA) (0.5 mM).
- -
- Group 5: each plant was sprayed with 25 mL of Kinetin (1 µM).
2.2. Determination of Growth Parameters and Leaf Relative Water Content
2.3. Ion Quantification and Net Translocation
2.4. Determination of Photosynthetic Pigments
2.5. Determination of Proline
2.6. Determination of Alcohol-Soluble Sugars and Sugar Alcohols
2.7. Determination of Starch
2.8. Determination of Free Polyamines
2.9. Determination of Ethylene Production
2.10. Statistical Analysis
3. Results
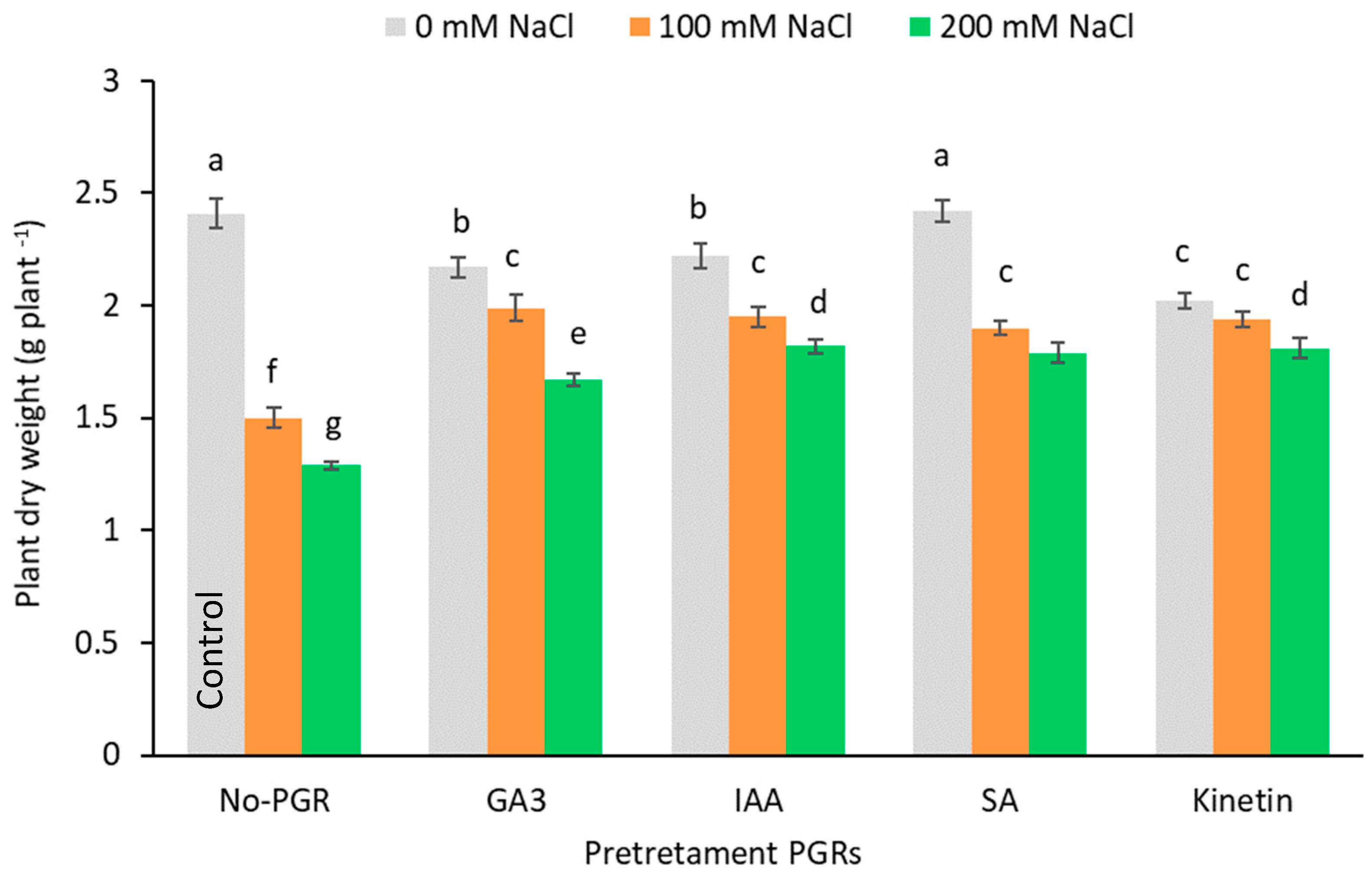
3.1. Effects of the Application of Plant Growth Regulators (PGRs) on Biomass
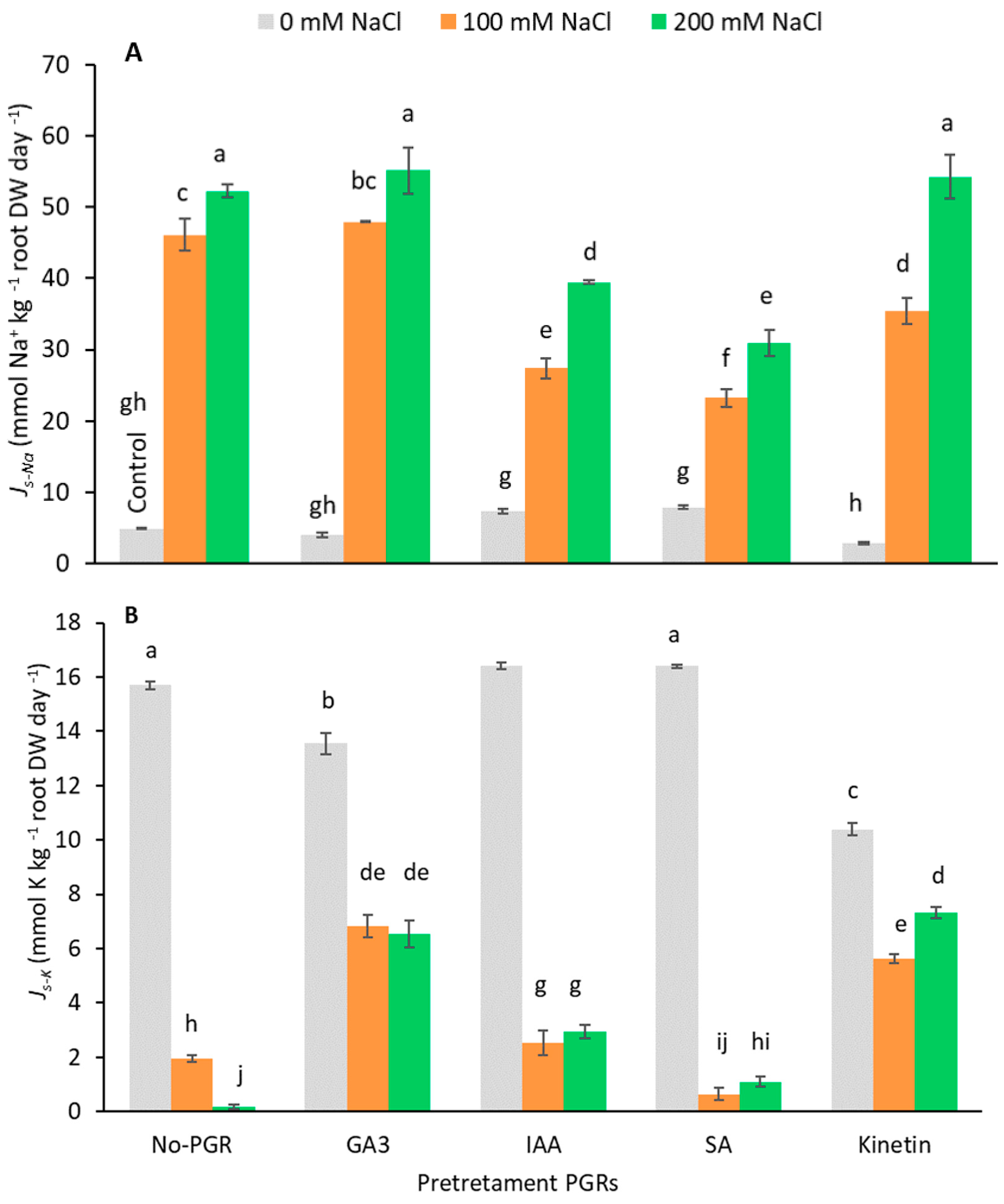
3.2. Effects of the Application of Plant Growth Regulators (PGRs) on Tissue Mineral Concentration
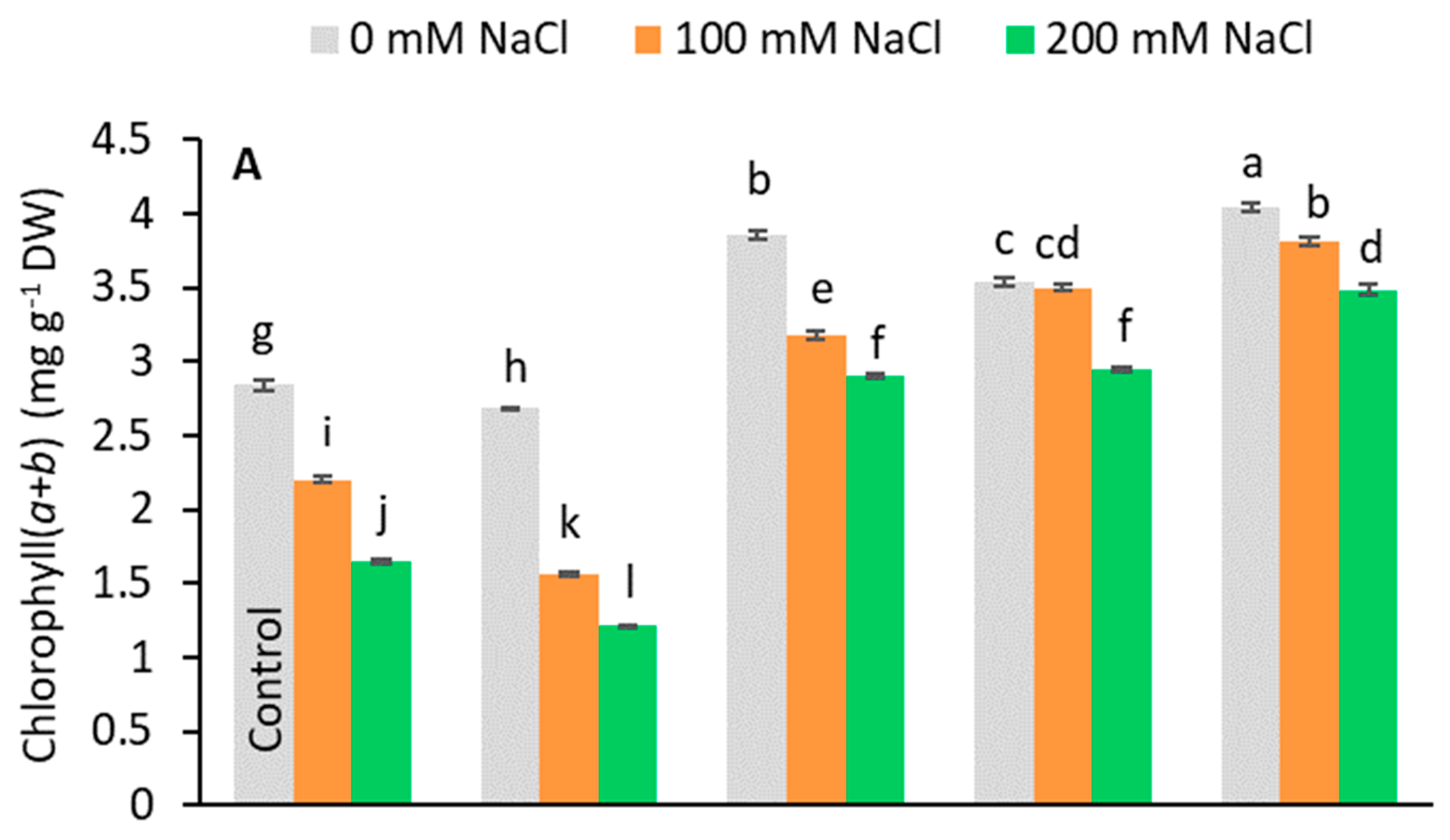
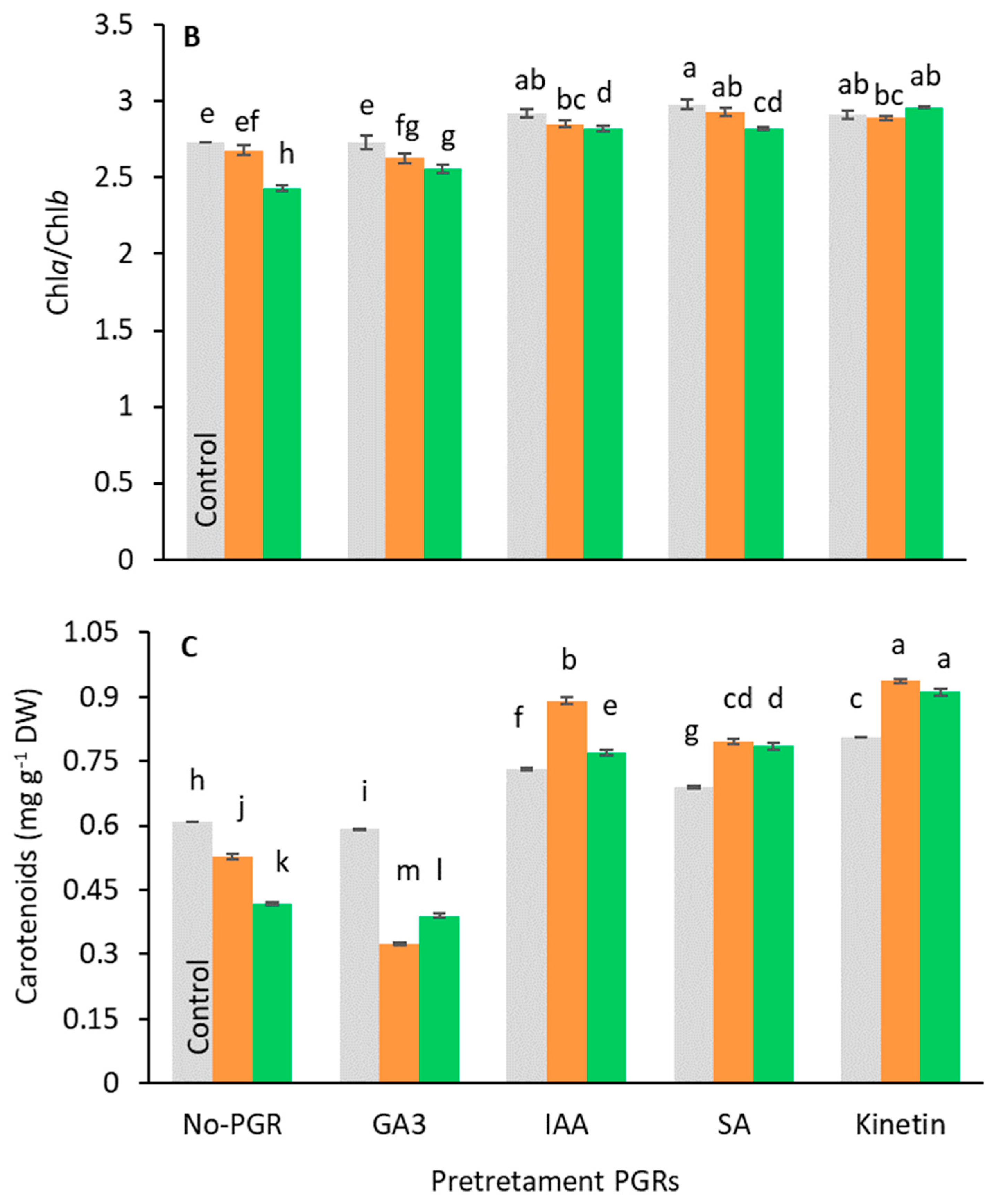
3.3. Effects of the Application Plant Growth Regulators (PGRs) on the Chlorophyll Content
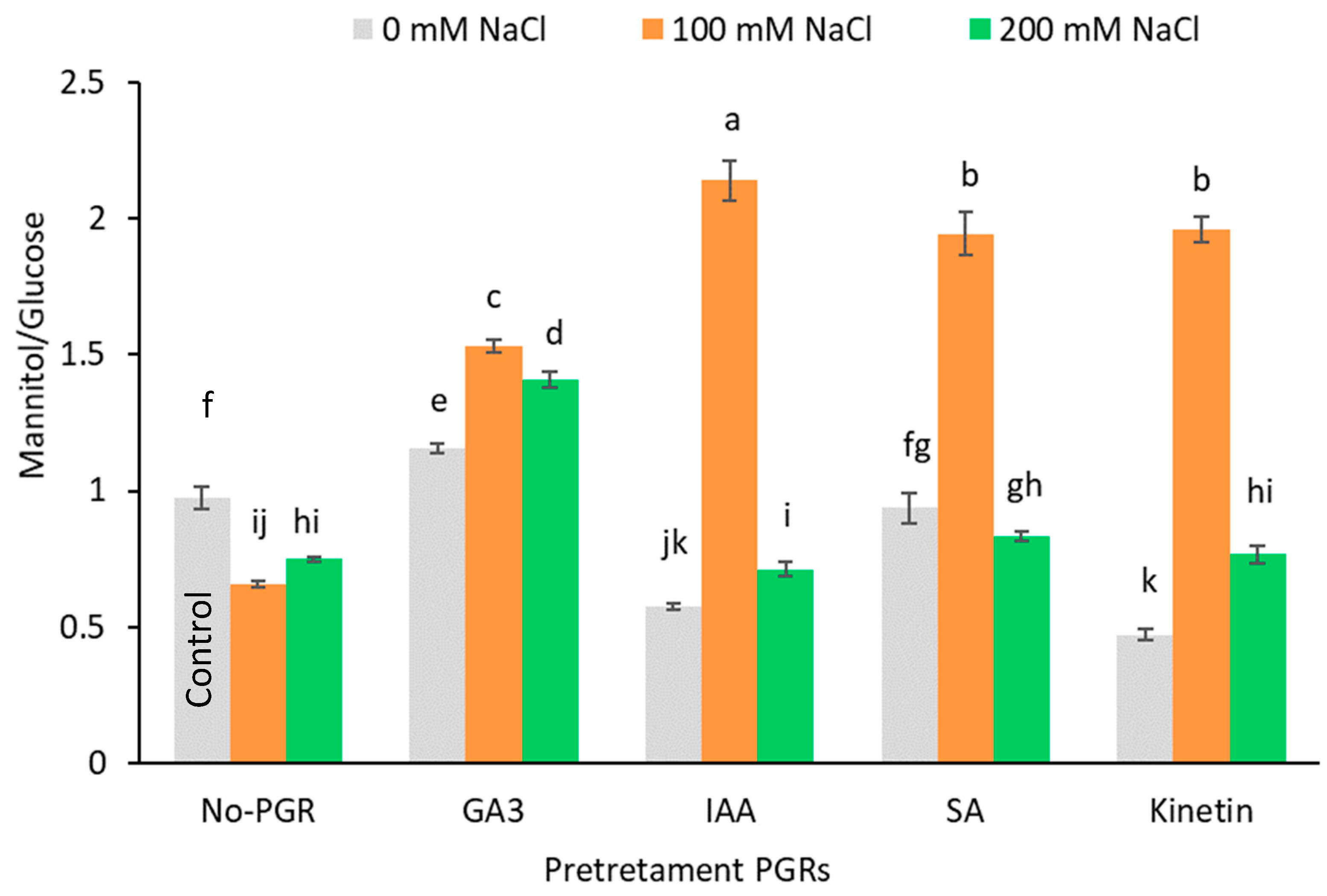
3.4. Effects of the Application of Plant Growth Regulators (PGRs) on Leaf Organic Solutes and Starch
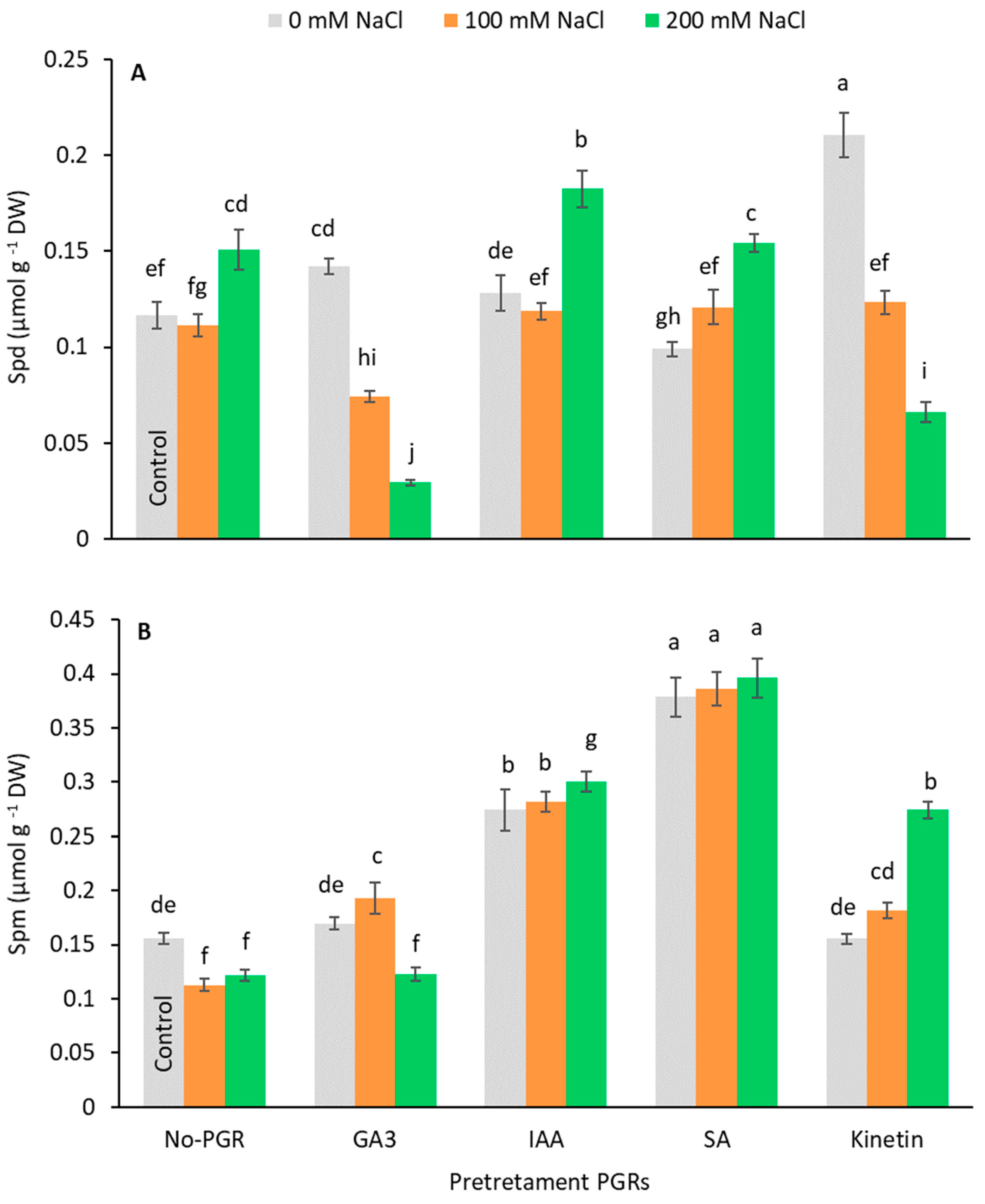
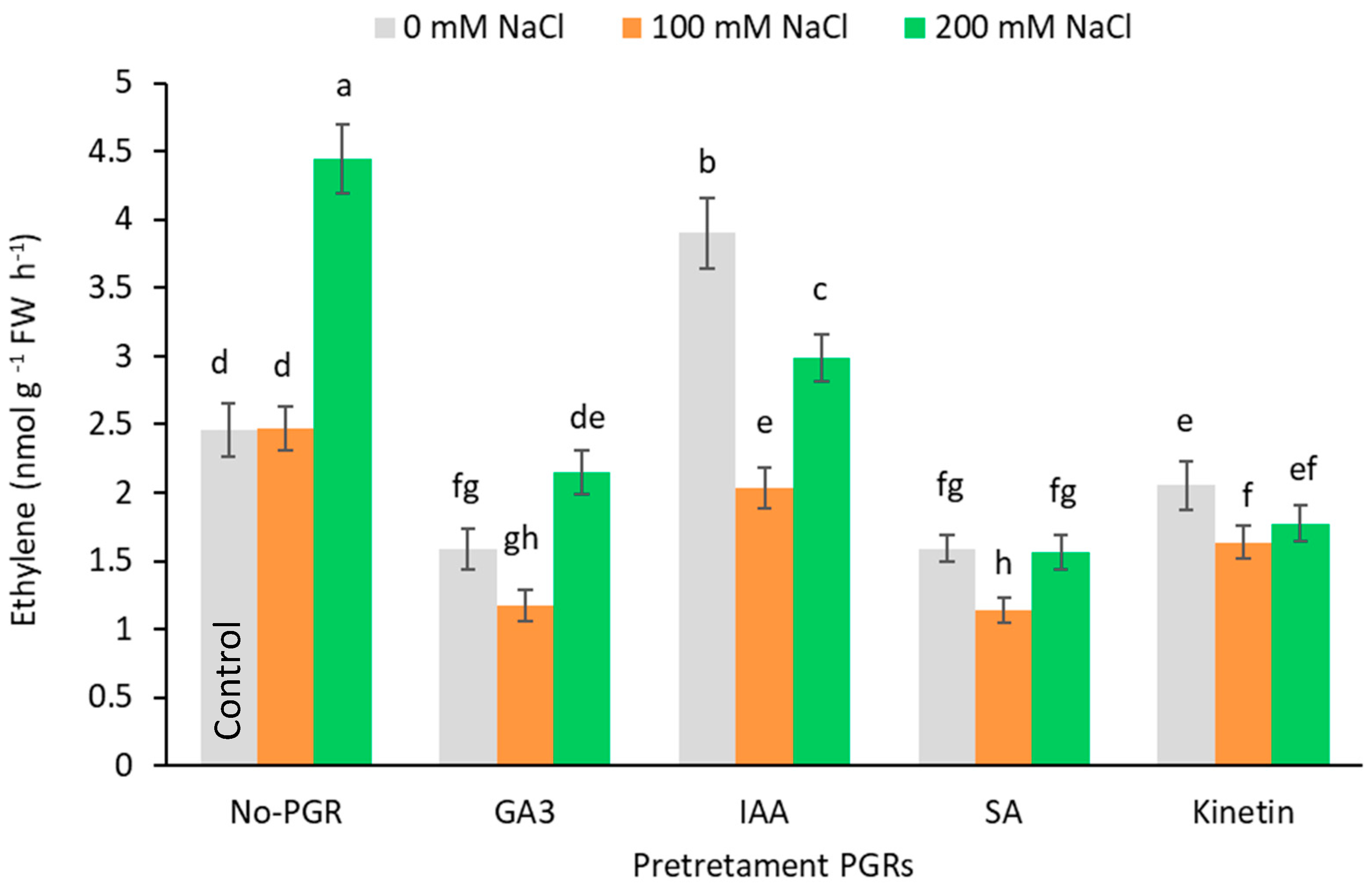
3.5. Effects of the Application of Plant Growth Regulators (PGRs) on Free Polyamine Content and Ethylene production
3.6. Multivariable Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haj-Amor, Z.; Araya, T.; Kim, D.G.; Bouri, S.; Lee, J.; Ghiloufi, W.; Yang, Y.; Kang, H.; Jhariya, M.K.; Banerjee, A.; et al. Soil salinity and its associated effects on soil microorganism, greenhouse gas emissions, crop yield, biodiversity and desertification: A review. Sci. Total Environ. 2022, 843, 156946. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Díaz-Vivancos, P.; Sánchez-Blanco, M.J.; Hernández, J.A. Plant responses to salt stress: Adaptative mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassani, A.; Azapagic, A.; Shokri, N. Global predictions of primary soil salinization under changing climate in the 21st century. Nat. Commun. 2021, 12, 6663. [Google Scholar] [CrossRef] [PubMed]
- Leisner, C.P. Review: Climate change impacts on food security-focus on perennial cropping systems and nutritional value. Plant Sci. 2020, 293, 110412. [Google Scholar] [CrossRef]
- Cuevas, J.; Daliakopoulos, I.N.; Moral, F.; Hueso, J.J.; Tsanis, I.K. A Review of Soil-Improving Cropping Systems for Soil Salinization. Agronomy 2019, 9, 295. [Google Scholar] [CrossRef] [Green Version]
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The threat of soil salinity: A European scale review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef]
- Feeding the World, Eradicating Hunger. In Proceedings of the World Summit on Food Security, Rome, Italy, 16–18 November 2019. (WSFS 2009/INF/2).
- Khan, M.Z.; Zaidi, S.S.A.; Amin, I.; Mansoor, S. A CRISPR way for fast-forward crop domestication. Trends Plant Sci. 2019, 24, 293–296. [Google Scholar] [CrossRef]
- Huang, Y.; Cao, H.; Yang, L.; Chen, C.; Shabala, L.; Xiong, M.; Niu, M.; Liu, J.; Zheng, Z.; Zhou, L.; et al. Tissue-specific respiratory burst oxidase homolog-dependent H2O2 signaling to the plasma membrane H+-ATPase confers potassium uptake and salinity tolerance in Cucurbitaceae. J. Exp. Bot. 2019, 70, 5879–5893. [Google Scholar] [CrossRef]
- Shailani, A.; Joshi, R.; Singla-Pareek, S.L.; Pareek, A. Stacking for future: Pyramiding genes to improve drought and salinity tolerance in rice. Physiol. Plant. 2021, 172, 1352–1362. [Google Scholar] [CrossRef]
- Chen, G.; Zheng, D.; Feng, N.; Zhou, H.; Mu, D.; Liu, L.; Zhao, L.; Shen, X.; Rao, G.; Li, T. Effects of exogenous salicylic acid and abscisic acid on growth, photosynthesis and antioxidant system of rice. Chil. J. Agric. Res. 2022, 82, 21–32. [Google Scholar] [CrossRef]
- Johnson, R.; Puthur, J.T. Seed priming as a cost effective technique for developing plants with cross tolerance to salinity stress. Plant Physiol. Biochem. 2021, 162, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Pacholczak, A.; Zajączkowska, M.; Nowakowska, K. The effect of brassinosteroids on rootting of stem cuttings in two barberry (Berberis thunbergii L.) cultivars. Agronomy 2021, 11, 699. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Imran, S.; Rauf, F.; Khatun, M.; Baskin, C.C.; Murata, Y.; Hasanuzzaman, M. Seed priming with phytohormones: An effective approach for the mitigation of abiotic stress. Plants 2021, 10, 37. [Google Scholar] [CrossRef]
- Tahir, M.T.; Anjum, M.A.; Saqib, M.; Khalid, M.F.; Hussain, S. Seed priming and foliar application of plant growth regulators affect the growth and yield of okra under calcareous soils. Acta Sci. Pol. Hortorum Cultus 2019, 18, 25–33. [Google Scholar] [CrossRef]
- Ashfaq, M.; Khan, S. Role of phytohormones in improving the yield of oilseed crops. In Oils Seed Crops: Yield and Adaptation Under Environmental Stress, 1st ed.; Ahmad, P., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; Volume 9, pp. 165–183. [Google Scholar] [CrossRef]
- Zheng, L.; Ma, H.; Jiao, Q.; Ma, C.; Wang, P. Phytohormones: Important participators in plant salt tolerance. Int. J. Agric. Biol. 2020, 24, 319–332. [Google Scholar] [CrossRef]
- Jayakannan, M.; Bose, J.; Babourina, O.; Rengel, Z.; Shabala, S. Salicylic acid in plant salinity stress signalling and tolerance. Plant Growth Regul. 2015, 76, 25–40. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Alaee, T.; Tavasolee, A. Salicylic acid but not jasmonic acid improved canola root response to salinity stress. Rhizosphere 2019, 9, 69–71. [Google Scholar] [CrossRef]
- Khoshbakht, D.; Asgharei, M.R. Influence of foliar-applied salicylic acid on growth, gas-exchange characteristics, and chlorophyll fluorescence in citrus under saline conditions. Photosynthetica 2015, 53, 410–418. [Google Scholar] [CrossRef]
- Khan, M.S.; Akther, T.; Ali, D.M.; Hemalatha, S. An investigation of the role of salicylic acid alleviates the saline stress in rice crop (Oryza sativa (L.)). Biocatal. Agric. Biotechnol. 2019, 18, 101027. [Google Scholar] [CrossRef]
- Bukhat, S.; Manzoor, H.; Athar, H.R.; Zafar, Z.U.; Azeem, F.; Rasul, S. Salicylic acid induced photosynthetic adaptability of Raphanus sativus to salt stress is associated with antioxidant capacity. J. Plant Growth Regul. 2020, 39, 809–822. [Google Scholar] [CrossRef]
- Dheeba, B.; Selvakumar, S.; Kannan, M.; Kannan, K. Effect of gibberellic acid on black gram (Vigna mungo) irrigated with different levels of saline water. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 709–720, WOS:000413622200121. [Google Scholar]
- Miceli, A.; Vetrano, F.; Moncada, A. Effects of foliar application of gibberellic acid on the salt tolerance of tomato and sweet pepper transplants. Horticulturae 2020, 6, 93. [Google Scholar] [CrossRef]
- Shahzad, K.; Hussain, S.; Arfan, M.; Hussain, S.; Waraich, E.A.; Zamir, S.; Saddique, M.; Rauf, A.; Kamal, K.Y.; Hano, C.; et al. Exogenously applied gibberellic acid enhances growth and salinity stress tolerance of maize through modulating the morpho-physiological, biochemical and molecular attributes. Biomolecules 2021, 11, 1005. [Google Scholar] [CrossRef] [PubMed]
- Moula, I.; Boussadia, O.; Koubouris, G.; Ben Hassine, M.; Boussetta, W.; Van Labeke, M.C.; Braham, M. Ecophysiological and biochemical aspects of olive tree (Olea europaea L.) in response to salt stress and gibberellic acid-induced alleviation. S. Afr. J. Bot. 2020, 132, 38–44. [Google Scholar] [CrossRef]
- Szepesi, A. Halotropism: Phytohormonal aspects and potential applications. Front. Plant Sci. 2020, 11, 571025. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latef, A.A.H.; Tahjib-Ul-Arif, M.; Rhaman, M.S. Exogenous auxin-mediated salt stress alleviation in faba bean (Vicia faba L.). Agronomy 2021, 11, 547. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, A.L.; Okant, A.M. Effect of foliar applied kinetin and indole acetic acid on maize plants grown under saline conditions. Turk. J. Agric. Forests. 2010, 34, 529–538. [Google Scholar] [CrossRef]
- Shah, S.H. Kinetin improves photosynthetic and antioxidant responses of Nigella sativa to counteract salt stress. Russ. J. Plant Physiol. 2011, 58, 454–459. [Google Scholar] [CrossRef]
- Nimir, N.E.A.; Zhou, G.; Guo, W.; Ma, B.; Lu, S.; Wang, Y. Effect of foliar application of GA3, kinetin, and salicylic acid on ions content, membrane permeability, and photosynthesis under salt stress of sweet sorghum [Sorghum bicolor (L.) Moench.]. Can. J. Plant Sci. 2017, 97, 525–535. [Google Scholar] [CrossRef]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small amines with large effects on plant abiotic stress tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef] [PubMed]
- Zapata, P.J.; Serrano, M.; García-Legaz, M.F.; Pretel, M.T.; Botella, M.A. Short term effect of salt shock on ethylene and polyamines depends on plant salt sensitivity. Front. Plant Sci. 2017, 8, 855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Therios, I. Olives: Crop Production Science in Horticulture 18; CABI Publishing: Wallingford, UK, 2009. [Google Scholar] [CrossRef]
- Aguilera, F.; Valenzuela, L.R. Time trends in the viability of pollen grains in the “Picual” olive (Olea europaea L.) cultivar. Palynology 2013, 37, 28–34. [Google Scholar] [CrossRef]
- Coppa, C.; Goncalves, B.; Lee, S.; Nunes, V.; Goncalves, C.; Rodrigues, C.; Oliveira, C. Extraction of oleuropein from olive leaves and applicability in foods. Qual. Assur. Saf. Crop. Foods 2020, 12, 50–62. [Google Scholar] [CrossRef]
- IOC. International Olive Council. Available online: http://www.internationaloliveoil.org (accessed on 28 December 2022).
- Rufino-Palomares, E.E.; Pérez-Jimenez, A.; García-Salguero, L.; Mokhtari, K.; Reyes-Zurita, F.J.; Peragón-Sanchez, J.; Lupiáñez, J.A. Nutraceutical role of polyphenols and triterpenes present in the extracts of fruits and leaves of Olea europaea as antioxidants, anti-infectives and anticancer agents on healthy growth. Molecules 2022, 27, 2341. [Google Scholar] [CrossRef] [PubMed]
- Brito, C.; Dinis, L.T.; Moutinho-Pereira, J.; Correia, C.M. Drought stress effects and olive tree acclimation under a changing climate. Plants 2019, 8, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trabelsi, L.; Gargouri, K.; Ayadi, M.; Mbadra, C.; Ben Nasr, M.; Ben Mbarek, H.; Ghrab, M.; Ben Ahemd, G.; Kammoun, Y.; Loukil, E.; et al. Impact of drought and salinity on olive potential yield, oil and fruit qualities (cv. Chemlali) in an arid climate. Agric. Water Manag. 2022, 269, 107726. [Google Scholar] [CrossRef]
- Methenni, K.; Ben Abdallah, M.; Nouairi, I.; Smaoui, A.; Ben Ammar, W.; Zarrouk, M.; Ben Youssef, N. Salicylic acid and calcium pretreatments alleviate the toxic effect of salinity in the Oueslati olive variety. Sci. Hort. 2018, 233, 349–358. [Google Scholar] [CrossRef]
- Perica, S.; Goreta, S.; Selak, G.V. Growth, biomass allocation and leaf ion concentration of seven olive (Olea europaea L.) cultivars under increased salinity. Sci. Hortic. 2008, 117, 123–129. [Google Scholar] [CrossRef]
- Soda, N.; Ephrath, J.E.; Dag, A.; Beiersdorf, I.; Presnov, E.; Yermiyahu, U.; Ben-Gal, A. Root growth dynamics of olive (Olea europaea L.) affected by irrigation induced salinity. Plant Soil 2017, 411, 305–318. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.L. The water culture methods for growing plants without soil. Calif. Agric. Exp. Stn. Bull. 1950, 374, 1–39. Available online: hdl.handle.net/2027/uc2.ark:/13960/t51g1sb8j (accessed on 20 February 2023).
- Aliniaeifard, S.; Hajilou, J.; Tabatabaei, S.J. Photosynthetic and growth responses of olive to proline and salicylic acid under salinity condition. Not. Bot. Horti. Agrobo. 2016, 44, 579–585. [Google Scholar] [CrossRef] [Green Version]
- Bueno, M.; Cordovilla, M.P. Spermidine pretreatments mitigate the effects of saline stress by improving growth and saline excretion in Frankenia pulverulenta. Agronomy 2021, 11, 1515. [Google Scholar] [CrossRef]
- Bueno, M.; Cordovilla, M.P. Plant growth regulators application enhance tolerance to salinity and benefit the halophyte Plantago coronopus in saline agriculture. Plants 2021, 10, 1872. [Google Scholar] [CrossRef]
- Tounekti, T.; Hernández, I.; Müller, M.; Khemira, H.; Munné-Bosch, S. Kinetin applications alleviate salt stress and improve the antioxidant composition of leaf extracts in Salvia officinalis. Plant Physiol. Biochem. 2011, 49, 1165–1176. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Santos, D.L.; Moutinho-Pereira, J.M.; GonÇalves, B.C.; Ferreira, H.F.; Correia, C.M. Inmediate responses and adaptative strategies of three olive cultivars under contrasting water availability regimes: Changes on structure and chemical composition of foliage and oxidative damage. Plant Sci. 2006, 170, 596–605. [Google Scholar] [CrossRef]
- Tattini, M.; Traversi, M.L. On the mechanism of salt tolerance in olive (Olea europea L.) under low- or high-Ca2+ supply. Environ. Exp. Bot. 2009, 65, 72–81. [Google Scholar] [CrossRef]
- Arnon, D.T. Copper enzyme in isolated chloroplasts polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Bates, L.S. Rapid determination of free proline water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bartolozzi, F.; Bertazza, G.; Bassi, D.; Cristoferi, G. Simultaneous determination of soluble sugars and organic acids as their trimethylsilyl derivatives in apricot fruits by gas-liquid chromatography. J. Chromatogr. 1997, 758, 99–107. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Emerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Chow, P.S.; Landhäusser, S.M. A method for routine measurements of total sugar and starch content in woody plant tissues. Tree Physiol. 2004, 24, 1129–1136. [Google Scholar] [CrossRef]
- Bueno, M.; Lendínez, M.L.; Calero, J.; Cordovilla, M.P. Salinity responses of three halophytes from inland saltmarshes of Jaén (southern Spain). Flora 2020, 266, 151589. [Google Scholar] [CrossRef]
- Bueno, M.; Lendínez, M.L.; Aparicio, C.; Cordovilla, M.P. Effect of salinity on polyamines and ethylene in Atriplex prostrata and Plantago coronopus. Biol. Plant. 2015, 59, 596–600. [Google Scholar] [CrossRef]
- Smith, M.A.; Davies, P.J. Separation and quantitation of polyamines in plant tissue by high performance liquid chromatography of their dansyl derivates. Plant Physiol. 1985, 78, 89–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bader, B.; Aissaoui, F.; Kmicha, I.; Ben Salem, A.; Chehab, H.; Gargouri, K.; Boujnah, D.; Chaieb, M. Effects of salinity stress water desalination, olive tree (Olea europaea L. cvs ‘Picholine’, ‘Meski’ and ‘Ascolana’) growth and ion accumulation. Desalination 2015, 364, 46–52. [Google Scholar] [CrossRef]
- Zörb, L.C.; Geilfus, C.M.; Dietz, K.J. Salinity and crop yield. Plant Biol. 2019, 21, 31–38. [Google Scholar] [CrossRef]
- Jie, S.; Baoshan, W. Using euhalophytes to understand salt tolerance and to develop saline agricultura: Suaeda salsa as a promising model. Rev. Ann. Bot. 2015, 115, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Amir, R.; Munir, F.; Khan, M.; Iqbal, T. Use of plant hormones for the improvement of plant growth and production under salt stress. In Sal Stress, Microbes, and Plant Interactions: Causes and Solution; Akhtar, M.S., Ed.; Springer Nature: Singapore, 2019; pp. 59–90. ISBN 978-981-13-8800-2. [Google Scholar] [CrossRef]
- Chunxin, Y.; Fan, Z.; Ruonan, W.; Zhaojin, R.; Weiming, T.; Linjiang, J.; Shunyan, C.; Zouli, X.; Yitao, X.; Yuyi, Z.; et al. B2, an abscisic acid mimic, improves salinity tolerance in winter wheat seedlings via improving activity of antioxidant enzymes. Front. Plant Sci. 2022, 13, 916287. [Google Scholar] [CrossRef]
- Islam, M.S.; Hasan, M.K.; Islam, B.; Renu, N.A.; Hakim, M.A.; Islam, M.R.; Chowdhyry, M.K.; Ueda, A.; Saneoka, M.A.; Raza, M.A.; et al. Responses of wáter and pigments status, dry matter partitioning, seed production, and traits of yield and quality to foliar application of GA3 in mugnbean (Vigna radiata L.). Front. Agron. 2021, 2, 596850. [Google Scholar] [CrossRef]
- Quamruzzaman, M.; Manik, S.M.N.; Shabala, S.; Zhou, M. Improving performance of salt-grown crops by exogenous application of plant growth regulators. Biomolecules 2021, 11, 788. [Google Scholar] [CrossRef]
- Zahid, G.; Iftikhar, S.; Shimira, F.; Ahmad, H.M.; Kaçar, Y.A. An overview and recent progress of plant growth regulators (PGRs) in the mitigation of abiotic stresses in frutis: A review. Sci. Hortic. 2023, 309, 111621. [Google Scholar] [CrossRef]
- Aparicio, C.; Urrestarazu, M.; Cordovilla, M.P. Comparative physiological analysis of salinity effects in six olive genotypes. HortScience 2014, 49, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, M.; Varol, N.; Yolcu, S.; Pelvan, A.; Kaya, Ü.; Aydoğdu, E.; Bor, M.; Özdemir, F.; Türkan, I. Three (Turkish) olive cultivars display contrasting salt stress-coping mechanisms under high salinity. Trees 2021, 35, 1283–1298. [Google Scholar] [CrossRef]
- Regni, L.; Del Pino, A.M.; Mousavi, S.; Palmerini, C.A.; Baldoni, L.; Mariotti, R.; Mairech, H.; Gardi, T.; D’Amato, R.; Proietti, P. Behavior of four olive cultivars during salt stress. Front. Plant. Sci. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Hoveizeh, N.F.; Gholami, R.; Abdelrahman, M.; Phan Tran, L.S. Exogenous melatonin mitigates salinity-induced damage in olive seedling by modulating ion homeostasis, antioxidant defense, and phytohormone balance. Physiol. Plant. 2021, 173, 1682–1694. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 25, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Kchaou, H.; Larbi, A.; Chaieb, M.; Sagardoy, R.; Msallem, M.; Morales, F. Genotypic differentiation in the stomatal response to salinity and contrasting photosynthetic and photoprotection responses in five olive (Olea europaea L.) cultivars. Sci. Hort. 2013, 160, 129–138. [Google Scholar] [CrossRef]
- Awais, G.W.; Amina, M.; Khurram, Z.; Basharat, A.; Muzammil, J.M.; Waseem, K.R.; Imran, K.; Muhammad, A.; Anam, N. Exogenously applied GA3 promotes plant growth in onion by reducing oxidative stress under saline conditions. J. Agric. Sci. 2021, 27, 122–128. [Google Scholar]
- Liu, Y.G.; Ye, N.H.; Liu, R.; Chen, M.; Zhang, J. H2O2 mediates the regulation of ABA catabolism and GA biosynthesis in Arabidopsis seed dormancy and germination. J. Exp. Bot. 2010, 61, 2979–2990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Yanli, L.; Zhenwei, Y.; Xiangbo, D. The role of cytokinins in plant under salt stress. J. Plant Growth Regul. 2022, 41, 2279–2291. [Google Scholar] [CrossRef]
- Husen, A.; Iqbal, M.; Aref, I.M. IAA-induced alteration in growth and photosynthesis of pea (Pisum sativum L.) plants grown under salt stress. J. Environ. Biol. 2016, 37, 421–429, WOS:000377747700014. [Google Scholar]
- Ribba, T.; Garrido-Vargas, F.; O’Brien, J.A. Auxin-mediated responses under salt stress: From development regulation to biotecnological applications. J. Exp. Bot. 2020, 13, 3843–3853. [Google Scholar] [CrossRef]
- Skodra, C.; Michailidis, M.; Dasenaki, M.; Ganopoulos, I.; Thomaidis, N.S.; Tanou, G.; Molassiotis, A. Uraveling salt-responsive tissue-specific metabolic pathways in olive tree. Physiol. Plant. 2021, 173, 1643–1656. [Google Scholar] [CrossRef]
- Demiral, M.A.; Uygun, D.A.; Uygun, M.; Kasirğa, E.; Karagözler, A.A. Biochemical response of Olea europea cv. Gemlik to short-term salt stress. Turk. J. Biol. 2011, 35, 433–442. [Google Scholar] [CrossRef]
- Vita, F.; Sabbatini, L.; Sillo, F.; Ghignone, S.; Vergine, M.; Nissim, W.G.; Fortunato, S.; Salzano, A.M.; Scaloni, A.; Luvisi, A.; et al. Salt stress in olive tree shapes resident endophytic microbiota. Front. Plant. Sci. 2022, 13, 992395. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Chartzoulakis, K. Salinity and olive: Growth, salt tolerance, photosynthesis and yield. Agric. Water Manag. 2005, 78, 108–121. [Google Scholar] [CrossRef]
- Ashraf, M.; Aasiya, K. Relationship between ion accumulation and growth in two spring wheat lines differing in salt-tolerance at different growth statges. J. Agron. Crop Sci. 1997, 178, 39–51. [Google Scholar] [CrossRef]
- Dehnavi, A.R.; Zahedi, M.; Ludwiczak, A.; Piernik, A. Foliar application of salicylic acid improves salt tolerance of sorghum (Sorghum bicolor (L.) Moench). Plants 2022, 11, 368. [Google Scholar] [CrossRef]
- Yang, F.; Xiao, X.; Zhang, S.; Korpelainen, H.; Li, C. Salt stress responses in Populus cathayana Rehder. Plant Sci. 2009, 176, 669–677. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to highsalinity. Annu. Rev. Plant. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjum, N.A.; Aref, I.M.; Duarte, A.C.; Pereira, E.; Ahmad, I.; Iqbal, M. Glutathione and proline cam coordinately make plants withstand the joint attack of metal (loid) and salinity stresses. Front. Plant Sci. 2014, 5, 662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Li, Z.G.; Duan, X.Q.; Min, X.; Zhou, Z.H. Methylglyoxal as a novel signal molecule, induces the salt tolerance of wheat by regulating the glyoxalase system, the antioxidant system, and osmolytes. Protoplasma 2017, 254, 1995–2006. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.; Abdelbaset, A.K.; Abd-Elkader, D. Salicylic acid effects on growth, yield, and fruit quality of strawberry cultivars. J. Med. Act. Plants 2018, 6, 1–11. Available online: https://scholarworks.umass.edu/jmap/vol6/iss2/2 (accessed on 20 February 2023).
- Roshdy, A.E.-D.; Alebidi, A.; Almutairi, K.; Al-Obeed, R.; Elsabagh, A. The effect of saliylic acid on the performances of salt stressed strawberry plants, enzymes activity, and salt tolerance index. Agronomy 2021, 11, 775. [Google Scholar] [CrossRef]
- Jabeen, N.; Ahmad, R. Foliar spray with potassium nitrate and salicylic acid for improving growth, yield and nutrients uptake by olive trees under salinity stress conditions. Not. Bot. Horti. Agrobo. 2011, 39, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Bashir, C.; Silvestri, C.; Coppa, E.; Brunori, E.; Cristofori, V.; Rugini, E.; Ahmad, T.; Hafiz, I.A.; Abbasi, N.A.; Nawaz Shah, M.K.; et al. Response of olive shoots to salinity stress suggests the involvement of sulfur metabolism. Plants 2021, 10, 350. [Google Scholar] [CrossRef]
- Khalid, A.; Aftab, F. Effect of exogenous application of IAA and GA3 on growth, protein content, and antioxidant enzymes of Solanum tubersoum L. grown in vitro under salt stress. Vitr. Cell. Dev. Biol. Plant 2020, 56, 377–389. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savoure, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sing, P.; Choudhary, K.K.; Chaudhary, N.; Gupta, S.; Sahu, M.; Tejaswini, B.; Sarkar, S. Salt stress resilience in plants mediated through osmolyte accumulation and its crosstalk mechanism with phytohormones. Front. Plant Sci. 2022, 13, 1006617. [Google Scholar] [CrossRef]
- Shomer, I.; Frenkel, H.; Polinger, C. The existence of a diffuse electric doublé layer at cellulose fbrill surfaces and its role in the swelling mechanism of parenchyma plant cell walls. Carbohydr. Polym. 1991, 16, 199–210. [Google Scholar] [CrossRef]
- Rady, M.M.; Taha, R.S.; Semida, W.M.; Alharby, H.F. Modulation of salt stress effects on Vicia faba L. plantas grown on reclaimed-saline soil by salicylic acid application. Rom. Agric. Res. 2017, 34, 175–185, DII 2067-5720 RAR 2017-3. [Google Scholar]
- Ashraf, U.; Hussain, S.; Anjum, S.A.; Abbas, F.; Tanveer, M.; Noor, M.A.; Tang, X. Alterations in growth, oxidative damage, and metal uptake of five aromatic rice cultivars under lead toxicity. Plant Physiol. Biochem. 2017, 115, 461–471. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Ali, E.F. Evaluation of proline functions in saline conditions. Phytochemistry 2017, 140, 52–68. [Google Scholar] [CrossRef]
- Per, T.S.; Fatma, M.; Asgher, M.; Asgher, M.; Javied, S.; Khan, N.A. Salicylic acid and nutrients interplay in abiotic stress tolerance. In Salicylic Acid: A Multifaceted Hormone; Springer: Singapore, 2017; pp. 221–237. [Google Scholar] [CrossRef]
- Ben Ahmed, C.; Magdich, S.; Ben Rouina, B.; Sensoy, S.; Boukhris, M.; Ben Abdullah, F. Exogenous proline effects on water relations and asyaskions contents in leaves and roots of young olive. Amino Acids 2011, 40, 565–573. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Das Bhowmik, S.; Long, H.; Cheng, Y.; Mundree, S.; Hoang, L.T.M. Rapid accumulation of proline enhances salinity tolerance in australian wild rice Oryza australiensis Domin. Plants 2021, 10, 2044. [Google Scholar] [CrossRef]
- Poury, n.; Seifi, E.; Alizadeh, M. Effects of salinity and proline on growth and physiological characteristics of tree olive cultivars. Gesunde Pflanz. 2022, 74, 1–12. [Google Scholar] [CrossRef]
- Lin, C.C.; Hsu, Y.T.; Kao, C.H. The effect of NaCl on proline accumulation in rice leaves. Plant Growth Regul. 2002, 36, 275–285, WOS:000177049600013. [Google Scholar] [CrossRef]
- Grzesiak, M.; Fileck, M.; Barbasz, A.; Kreczmer, B.; Hartikainen, H. Relationships between polyamines, ethylene, osmoprotectans and antioxidant enzymes activities in wheat seedlings after short-term PEG- and NaCl-induced stresses. Plant Growth Regul. 2013, 69, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Conde, C.; Delrot, S.; Gero’s, H. Physiological, biochemical and molecular changes occurring during olive development and ripening. J. Plant Physiol. 2008, 165, 1545–1562. [Google Scholar] [CrossRef]
- Kumari, A.; Parida, A.K. Metabolomics and network analysis reveal the potential metabolites and biological pathways involved in salinity tolerance of hte halophyte Salvadora persica. Environ. Exp. Bot. 2018, 148, 85–99. [Google Scholar] [CrossRef]
- Bano, C.; Amis, N.; Singh, N.B. Role of polyamines in plants abiotic stress tolerance: Advances and future prospects. In Plant Life Under Changing Environment; Tripathi, D.K., Sing, V.P., Chauhan, D.K., Prasad, S.M., Dubey, N.K., Ramawat, N., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 481–496. [Google Scholar]
- Bueno, M.; Cordovilla, M.P. Polyamines in halophytes. Front. Plant Sci. 2019, 10, 439. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Smith, J.A.C.; Harberd, N.P.; Jiang, C. The regulatory roles of ethylene and reactive oxygen species (ROS) in plant salt stress responses. Plant Mol. Biol. 2016, 91, 651–659. [Google Scholar] [CrossRef]
- Liu, J.H.; Wang, W.; Wu, H.; Gong, X.; Moriguchi, T. Polyamines function in stress tolerance: From synthesis to regulation. Front. Plant Sci. 2015, 6, 827. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, R.; Barchi, B.; Turlapati, S.A.; Gagne, M.; Minocha, R.; Long, S.; Minocha, S.C. Glutamate, ornithine, arginine, proline, and polyamine metabolic interactions: The pathway is regulated at the post-transcriptional level. Front. Plant Sci. 2016, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; He, J. Advance in metabolism and response to stress of polyamines in plant. Acta Agric. Boreali Sinica 2012, 27, 240–245. [Google Scholar] [CrossRef]
- Parvin, S.; Lee, O.R.; Sathiyaraj, G.; Khorolragchaa, A.; Kim, Y.J.; Yang, D.C. Spermidine alleviates the growth of saline-stressed ginseng seedlings through antioxidative defense system. Gene 2014, 537, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Jain, V.; Vart, S.; Verma, E.; Malhotra, S.P. Spermine reduces salinity-induced oxidative damage by enhancing antioxidative system and decreasing lipid peroxidation in rice seedlings. J. Plant Biochem. Bioecnol. 2015, 24, 316–323. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGRs without NaCl | Root Dry Weight (g plant−1) | Stem Dry Weight (g plant−1) | Leaf Dry Weight (g plant−1) | LRWC (%) |
|---|---|---|---|---|
| Control: 0 mM NaCl (no-PGR) | 0.56 ± 0.023 a | 0.78 ± 0.026 a | 1.16 ± 0.030 ab | 93.37 ± 0.351 a |
| GA3 + 0 mM | 0.43 ± 0.013 cd | 0.66 ± 0.025 b | 1.08 ± 0.048 bc | 83.21 ± 0.525 e |
| IAA + 0 mM | 0.36 ± 0.012 e | 0.55 ± 0.019 cd | 1.05 ± 0.042 c | 93.26 ± 0.350 a |
| SA + 0 mM | 0.57 ± 0.021 a | 0.68 ± 0.031 b | 1.17 ± 0.033 a | 85.46 ± 0.453 cd |
| Kinetin + 0 mM | 0.50 ± 0.014 b | 0.51 ± 0.008 c | 1.00 ± 0.024 cd | 88.76 ± 0.701 b |
| PGRs with 100 mM NaCl | Root dry weight (g plant−1) | Stem dry weight (g plant−1) | Leaf dry weight (g plant−1) | LRWC (%) |
| 100 mM (no-PGR) | 0.32 ± 0.007 ef | 0.43 ± 0.015 d | 0.75 ± 0.030 f | 82.30 ± 0.276 ef |
| GA3 + 100 mM | 0.40 ± 0.014 d | 0.52 ± 0.022 c | 1.07 ± 0.041 c | 82.13 ± 0.471 ef |
| IAA + 100 mM | 0.34 ± 0.013 ef | 0.67 ± 0.030 b | 0.93 ± 0.033 de | 86.30 ± 0.653 c |
| SA + 100 mM | 0.43 ± 0.015 cd | 0.43 ± 0.015 d | 0.96 ± 0.037 de | 82.49 ± 0.611 ef |
| Kinetin + 100 mM | 0.46 ± 0.011 c | 0.52 ± 0.022 c | 0.95 ± 0.029 de | 84.60 ± 0.456 d |
| PGRs with 200 mM NaCl | Root dry weight (g plant−1) | Stem dry weight (g plant−1) | Leaf dry weight (g plant−1) | LRWC (%) |
| 200 mM (no-PGR) | 0.26 ± 0.008 g | 0.37 ± 0.015 e | 0.66 ± 0.016 g | 77.84 ± 0.471 h |
| GA3 + 200 mM | 0.26 ± 0.010 g | 0.51 ± 0.021 c | 0.89 ± 0.019 e | 78.75 ± 0.565 h |
| IAA + 200 mM | 0.26 ± 0.010 g | 0.65 ± 0.023 b | 0.90 ± 0.036 e | 81.41 ± 0.210 fg |
| SA + 200 mM | 0.36 ± 0.011 e | 0.49 ± 0.021 c | 0.94 ± 0.040 de | 77.80 ± 0.524 h |
| Kinetin + 200 mM | 0.34 ± 0.012 e | 0.52 ± 0.010 c | 0.94 ± 0.020 de | 79.25 ± 0.436 g |
| PGRs without NaCl | Leaf | ||
|---|---|---|---|
| Na+ (mmol kg−1 DW) | K+ (mmol kg−1 DW) | K+/Na+ Ratio | |
| Control: 0 mM NaCl (no-PGR) | 66.96 ± 0.50 gh | 357.38 ± 1.56 a | 4.900 ± 0.023 c |
| GA3 + 0 mM | 53.37 ± 3.86 gh | 299.03 ± 4.31 c | 5.603 ± 0.081 b |
| IAA + 0 mM | 76.31± 2.34 gh | 305.12 ± 1.08 c | 3.999 ± 0.014 d |
| SA + 0 mM | 98.01± 0.17 g | 331.81 ± 0.71 b | 3.386 ± 0.007 e |
| Kinetin + 0 mM | 47.25± 2.84 h | 289.14 ± 2.86 d | 6.120 ± 0.061 a |
| PGRs with 100 mM NaCl | Na+ (mmol kg−1 DW) | K+ (mmol kg−1 DW) | K+/Na+ ratio |
| 100 mM NaCl (no-PGR) | 610.13 ± 28.70 b | 247.15 ± 1.51 fg | 0.405 ± 0.002 ij |
| GA3 + 100 mM | 490.22 ± 1.11 d | 224.25 ± 4.19 h | 0.457 ± 0.009 i |
| IAA + 100 mM | 347.57 ± 17.26 f | 265.86 ± 5.42 e | 0.765 ± 0.015 f |
| SA + 100 mM | 334.68 ± 16.64 f | 264.06 ± 3.04 e | 0.789 ± 0.009 f |
| Kinetin + 100 mM | 457.37 ± 23.00 de | 246.81 ± 2.17 g | 0.540 ± 0.005 h |
| PGRs with 200 mM NaCl | Na+ (mmol kg−1 DW) | K+ (mmol kg−1 DW) | K+/Na+ ratio |
| 200 mM NaCl (no-PGR) | 693.93± 12.25 a | 252.23 ± 3.42 fg | 0.364 ± 0.005 j |
| GA3 + 200 mM | 552.14± 31.25 c | 251.55 ± 4.84 fg | 0.456 ± 0.009 i |
| IAA + 200 mM | 472.06 ± 2.18 d | 302.96 ± 2.83 c | 0.642 ± 0.006 g |
| SA + 200 mM | 414.96 ± 23.37 e | 267.24 ± 2.47 e | 0.644 ± 0.006 g |
| Kinetin + 200 mM | 593.00 ± 32.77 bc | 256.08 ± 2.34 f | 0.432 ± 0.004 ij |
| PGRs without NaCl | Root | ||
|---|---|---|---|
| Na+ (mmol kg−1 DW) | K+ (mmol kg−1 DW) | K+/Na+ Ratio | |
| Control: 0 mM NaCl (no-PGR) | 122.62 ± 0.71 k | 856.44 ± 2.96 a | 6.984 ± 0.024 b |
| GA3 + 0 mM | 159.88 ± 0.69 j | 603.58 ± 1.90 e | 3.775 ± 0.012 e |
| IAA + 0 mM | 120.54 ± 0.91 k | 835.34 ± 0.73 b | 6.930 ± 0.006 c |
| SA + 0 mM | 159.13 ± 0.48 j | 686.18 ± 0.59 d | 4.312 ± 0.004 d |
| Kinetin + 0 mM | 97.28 ± 0.17 l | 750.89 ± 1.15 c | 7.719 ± 0.012 a |
| PGRs with 100 mM NaCl | Na+ (mmol kg−1 DW) | K+ (mmol kg−1 DW) | K+/Na+ ratio |
| 100 mM NaCl (no-PGR) | 1790.29 ± 8.15 b | 367.19 ± 1.21 g | 0.205 ± 0.002 g |
| GA3 + 100 mM | 1187.19 ± 2.55 h | 309.95 ± 1.24 i | 0.261 ± 0.001 f |
| IAA + 100 mM | 1594.39 ± 1.21 e | 397.57 ± 2.90 f | 0.249 ± 0.001 f |
| SA + 100 mM | 1232.79 ± 6.06 g | 210.26 ± 1.63 l | 0.177 ± 0.001 h |
| Kinetin + 100 mM | 1144.83 ± 0.37 i | 170.35 ± 2.27 m | 0.149 ± 0.002 i |
| PGRs with 200 mM NaCl | Na+ (mmol kg−1 DW) | K+ (mmol kg−1 DW) | K+/Na+ ratio |
| 200 mM NaCl (no-PGR) | 1899.94 ± 1.51 a | 267.02 ± 3.64 j | 0.141 ± 0.002 i |
| GA3 + 200 mM | 1392.42 ± 2.43 f | 124.27 ± 1.17 n | 0.089 ± 0.001 j |
| IAA + 200 mM | 1778.69 ± 3.61 c | 323.92 ± 4.27 h | 0.182 ± 0.002 h |
| SA + 200 mM | 1658.42 ± 2.57 d | 243.37 ± 2.99 k | 0.147 ± 0.002 i |
| Kinetin + 200 mM | 1592.74 ± 3.21 e | 162.79 ± 2.37 m | 0.102 ± 0.001 j |
| PGRs without NaCl | Proline (µg g−1 DW) | Sucrose (mg g−1 DW) | Starch (mg g−1 DW) |
|---|---|---|---|
| Control: 0 mM NaCl (no PGR) | 167.60 ± 4.64 c | 13.40 ± 0.09 ab | 16.11 ± 0.63 cd |
| GA3 + 0 mM | 183.63 ± 4.24 ab | 12.41 ± 0.12 d | 16.47 ± 0.14 cd |
| IAA + 0 mM | 137.29 ± 3.05 d | 13.47 ± 0.26 ab | 2.42 ± 0.51 f |
| SA + 0 mM | 159.26 ± 2.57 c | 11.72 ± 0.19 e | 21.68 ± 1.92 b |
| Kinetin + 0 mM | 163.32 ± 1.76 c | 13.77 ± 0.14 a | 15.71 ± 0.59 d |
| PGRs with 100 mM NaCl | Proline (µg g−1 DW) | Sucrose (mg g−1 DW) | Starch (mg g−1 DW) |
| 100 mM NaCl (no PGR) | 188.19 ± 2.51 ab | 13.66 ± 0.24 ab | 16.28 ± 0.78 cd |
| GA3 + 100 mM | 133.90 ± 0.55 d | 13.46 ± 0.07 ab | 22.31 ± 0.78 b |
| IAA + 100 mM | 203.10 ± 3.17 a | 11.17 ± 0.20 e | 2.89 ± 0.35 f |
| SA + 100 mM | 199.94 ± 4.20 a | 11.60 ± 0.25 e | 15.63 ± 0.62 d |
| Kinetin + 100 mM | 199.98 ± 4.11 a | 12.81 ± 0.27 cd | 30.44 ± 0.57 a |
| PGRs with 200 mM NaCl | Proline (µg g−1 DW) | Sucrose (mg g−1 DW) | Starch (mg g−1 DW) |
| 200 mM NaCl (no PGR) | 171.02 ± 1.62 c | 6.90 ± 0.16 h | 5.79 ± 0.11 e |
| GA3 + 200 mM | 158.25 ± 2.84 d | 9.18 ± 0.22 f | 1.05 ± 0.05 f |
| IAA + 200 mM | 197.19 ± 1.89 a | 8.36 ± 0.14 g | 6.31 ± 0.26 e |
| SA + 200 mM | 190.51 ± 3.62 ab | 8.13 ± 0.16 g | 17.97 ± 0.19 c |
| Kinetin + 200 mM | 167.85 ± 3.24 c | 6.79 ± 0.32 h | 2.72 ± 0.63 f |
| PGRs without NaCl | Fructose (mg g−1 DW) | Glucose (mg g−1 DW) | Inositol (mg g−1 DW) | Mannitol (mg g−1 DW) |
|---|---|---|---|---|
| Control: 0 mM NaCl (no PGR) | 14.99 ± 0.19 e | 23.10 ± 0.35 e | 1.59 ± 0.046 a | 22.52 ± 1.41 d |
| GA3 + 0 mM | 21.36 ± 0.12 b | 14.96 ± 0.91 g | 0.86 ± 0.054 d | 17.32 ± 1.23 e |
| IAA + 0 mM | 22.46 ± 0.44 b | 30.77 ± 0.73 d | 1.66 ± 0.052 a | 17.79 ± 0.34 e |
| SA + 0 mM | 19.36 ± 0.37 c | 15.60 ± 0.90 g | 0.76 ± 0.034 e | 14.61 ± 0.87 f |
| Kinetin + 0 mM | 17.49 ± 0.22 d | 37.12 ± 1.21 c | 1.25 ± 0.036 b | 17.54 ± 0.76 e |
| PGRs with 100 mM NaCl | Fructose (mg g−1 DW) | Glucose (mg g−1 DW) | Inositol (mg g−1 DW) | Mannitol (mg g−1 DW) |
| 100 mM NaCl (no PGR) | 14.60 ± 0.10 e | 45.24 ± 2.80 b | 0.90 ± 0.013 d | 29.79 ± 0.57 c |
| GA3 + 100 mM | 14.90 ± 0.13 e | 19.34 ± 0.82 f | 0.87 ± 0.031 d | 29.64 ± 1.62 c |
| IAA + 100 mM | 21.10 ± 0.06 b | 14.99 ± 1.48 g | 1.01 ± 0.060 c | 33.07 ± 1.11 b |
| SA + 100 mM | 17.49 ± 0.38 d | 15.58 ± 0.60 g | 0.77 ± 0.041 e | 30.30 ± 1.21 c |
| Kinetin + 100 mM | 21.49 ± 0.06 b | 17.32 ± 0.87 fg | 0.62 ± 0.025 g | 33.94 ± 0.83 b |
| PGRs with 200 mM NaCl | Fructose (mg g−1 DW) | Glucose (mg g−1 DW) | Inositol (mg g−1 DW) | Mannitol (mg g−1 DW) |
| 200 mM NaCl (no PGR) | 13.95 ± 0.18 ef | 31.85 ± 0.27 d | 0.50 ± 0.020 h | 23.92 ± 0.27 d |
| GA3 + 200 mM | 13.04 ± 0.12 f | 23.92 ± 0.90 e | 0.69 ± 0.015 f | 33.71 ± 1.66 b |
| IAA + 200 mM | 23.97 ± 0.42 a | 50.13 ± 0.77 a | 0.40 ± 0.021 h | 35.74 ± 1.21 a |
| SA + 200 mM | 16.84 ± 0.15 d | 36.57 ± 1.55 c | 0.64 ± 0.040 g | 30.53 ± 0.37 c |
| Kinetin + 200 mM | 16.45 ± 0.35 d | 46.77 ± 2.10 b | 0.48 ± 0.019 h | 37.95 ± 1.12 a |
| PDW | RDW | SDW | LDW | LRWC | Leaf Na+ | Leaf K+ | |
|---|---|---|---|---|---|---|---|
| PDW | 1 | ||||||
| RDW | 0.8092 ** | 1 | |||||
| SDW | 0.7951 ** | 0.4840 | 1 | ||||
| LDW | 0.9264 ** | 0.6428 ** | 0.6175 * | 1 | |||
| LRWC | 0.7559 ** | 0.6077 * | 0.5570 * | 0.7245 ** | 1 | ||
| Leaf Na+ | −0.8690 ** | −0.7438 ** | −0.6246 * | −0.8158 ** | −0.7737 ** | 1 | |
| Leaf K+ | 0.7212 ** | 0.5228 * | 0.7212 ** | 0.6091 * | 0.6070 * | −0.7841 ** | 1 |
| Leaf K+/Na+ | 0.6949 ** | 0.6545 ** | 0.5045 | 0.6365 * | 0.6998 ** | −0.9093 ** | 0.7239 ** |
| Root Na+ | −0.8455 ** | −0.7809 ** | −0.5252 * | −0.8300 ** | −0.7650 ** | 0.9304 ** | −0.6926 ** |
| Root K+ | 0.7182 ** | 0.6002 * | 0.5595 * | 0.6955 ** | 0.8494 ** | −0.8569 ** | 0.7818 ** |
| Root K+/Na+ | 0.7052 ** | 0.6388 * | 0.4574 | 0.8339 ** | 0.8339 ** | −0.9199 ** | 0.7354 ** |
| Js-Na | −0.7999 ** | −0.7435 ** | −0.5577 * | −0.7174 ** | −0.7563 ** | 0.9764 ** | −0.7861 ** |
| Js-K | 0.8350 ** | 0.6437 ** | 0.6306* | 0.8501 ** | 0.7150 ** | −0.7894 ** | 0.7049 ** |
| Chl | 0.5039 | 0.5230 * | 0.2466 | 0.4662 | 0.5401 * | −0.5616 * | 0.4609 |
| Chla/b | 0.5468 * | 0.4873 | 0.3065 | 0.5326 * | 0.4120 | −0.4890 | 0.4066 |
| Car | 0.2412 | 0.2321 | 0.1985 | 0.1669 | 0.2457 | −0.2362 | 0.2293 |
| Proline | −0.2345 | −0.1155 | 0.0231 | −0.4396 | −0.1977 | 0.1183 | 0.0663 |
| Fructose | 0.3904 | 0.0937 | 0.4410 | 0.3928 | 0.3379 | −0.4178 | 0.4368 |
| Glucose | −0.4568 | −0.4795 | −o.2779 | −0.3839 | −0.2298 | 0.3864 | −0.0613 |
| Inositol | 0.6037 * | 0.4641 | 0.3635 | 0.6469 ** | 0.8994 ** | −0.7045 ** | 0.4449 |
| Mannitol | −0.5269 * | −0.5735 * | −0.2495 | −0.5232 * | −0.5349 * | 0.7630 ** | 0.6334 * |
| Sucrose | 0.5544 * | 0.6131 * | 0.2536 | 0.528 * | 0.7128 ** | −0.5960 * | 0.1919 |
| Starch | 0.3059 | 0.6559 ** | 0.0383 | 0.1778 | 0.0760 | −0.2074 | −0.0301 |
| Spd | 0.0170 | 0.1233 | 0.0209 | −0.0655 | 0.2079 | −0.3418 | 0.3508 |
| Spm | 0.2843 | 0.1468 | 0.1413 | 0.3141 | −0.0572 | −0.1882 | 0.2791 |
| Ethylene | −0.3180 | −0.4574 | −0.1582 | −0.2197 | 0.1455 | 0.1422 | 0.1709 |
| Chl | Chla/b | Car | Proline | Fructose | Glucose | Inositol | |
| Chl | 1 | ||||||
| Chla/b | 0.9175 ** | 1 | |||||
| Car | 0.8799 ** | 0.8425 ** | 1 | ||||
| Proline | 0.3009 | 0.2226 | 0.5777 * | 1 | |||
| Fructose | 0.6258 * | 0.5921 * | 0.6137 * | 0.3294 | 1 | ||
| Glucose | 0.0028 | −0.0092 | 0.1048 | 0.0769 | −0.0719 | 1 | |
| Inositol | 0.2856 | 0.1636 | −0.0188 | −0.3429 | 0.0517 | −0.2112 | 1 |
| Mannitol | −0.2284 | −0.0730 | 0.1936 | 0.3290 | −0.1032 | 0.3068 | −0.5718 * |
| Sucrose | 0.2544 | 0.2032 | −0.0606 | −0.2482 | 0.1440 | −0.4065 | 0.7127 ** |
| Starch | 0.1918 | 0.1716 | 0.0461 | 0.0171 | 0.0289 | −0.3616 | −0.0280 |
| Spd | 0.4439 | 0.1871 | 0.3503 | 0.3610 | 0.4445 | 0.27777 | 0.1192 |
| Spm | 0.4941 | 0.6689 ** | 0.5334 * | 0.2186 | 0.4168 | −0.0702 | −0.1733 |
| Ethylene | −0.1539 | −0.1539 | −0.2110 | −0.0708 | 0.0352 | 0.3927 | 0.1839 |
| Leaf K+/Na+ | Root Na+ | Root K+ | Root K+/Na+ | Js-Na | Js-K | ||
| Leaf K+/Na+ | 1 | ||||||
| Root Na+ | −0.9093 ** | 1 | |||||
| Root K+ | 0.8818 ** | −0.8452 ** | 1 | ||||
| Root K+/Na+ | 0.8818 ** | 0.9092 ** | 0.9412 ** | 1 | |||
| Js-Na | −0.8855 ** | −0.8744 ** | −0.8506 ** | −0.8433 ** | 1 | ||
| Js-K | 0.7972 ** | −0.9117 ** | 0.7995 ** | 0.8296 ** | −0.6905 ** | 1 | |
| Chl | 0.4103 | −0.4456 | 0.3837 | 0.4542 | −0.6140 * | 0−3224 | |
| Chla/b | 0.2591 | −0.364 | 0.2492 | 0.2956 | −0.4930 | 0.3098 | |
| Car | 0.0640 | −0.0521 | 0.0107 | 0.0714 | −0.2929 | −0.0220 | |
| Proline | −0.1896 | 0.3479 | −0.2804 | −0.3207 | −0.0250 | −0.4719 | |
| Fructose | 0.2313 | −0.2440 | 0.2476 | 0.1745 | −0.4419 | 0.2214 | |
| Glucose | −0.1943 | 0.4030 | −0.1204 | −0.10006 | 0.3644 | −0.2708 | |
| Inositol | 0.6789 ** | −0.7065 ** | 0.8257 ** | 0.8151 ** | −0.6784 ** | 0.6328 * | |
| Mannitol | −0.8294 ** | 0.8182 ** | −0.832 ** | −0.8107 ** | 0.7788 ** | −0.7032 ** | |
| Sucrose | 0.5347 * | −0.6429 ** | 0.6046 * | 0.5575 * | −0.5875 * | 0.4866 | |
| Starch | 0.1360 | −0.2680 | 0.0656 | 0.0606 | −0.2610 | 0.1052 | |
| Spd | 0.4061 | −0.1614 | 0.3627 | 0.3522 | −0.4692 | −0.0793 | |
| Spm | −0.1414 | 0.0273 | −0.069 | −0.1130 | −0.2154 | −0.0613 | |
| Ethylene | 0.0299 | 0.1015 | 0.2341 | 0.1890 | 0.0896 | 0.0033 | |
| Mannitol | Sucrose | Starch | Spd | Spm | Ethylene | ||
| Mannitol | 1 | ||||||
| Sucrose | −0.4836 | 1 | |||||
| Starch | −0.2035 | 0.5082 | 1 | ||||
| Spd | −0.3385 | 0.0933 | 0.1495 | 1 | |||
| Spm | 0.0688 | −0.2388 | 0.0366 | 0.0932 | 1 | ||
| Ethylene | −0.1032 | −0.2325 | −0.5471 * | 0.2777 | −0.3262 | 1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
del Pilar Cordovilla, M.; Aparicio, C.; Melendo, M.; Bueno, M. Exogenous Application of Indol-3-Acetic Acid and Salicylic Acid Improves Tolerance to Salt Stress in Olive Plantlets (Olea europaea L. Cultivar Picual) in Growth Chamber Environments. Agronomy 2023, 13, 647. https://doi.org/10.3390/agronomy13030647
del Pilar Cordovilla M, Aparicio C, Melendo M, Bueno M. Exogenous Application of Indol-3-Acetic Acid and Salicylic Acid Improves Tolerance to Salt Stress in Olive Plantlets (Olea europaea L. Cultivar Picual) in Growth Chamber Environments. Agronomy. 2023; 13(3):647. https://doi.org/10.3390/agronomy13030647
Chicago/Turabian Styledel Pilar Cordovilla, María, Carolina Aparicio, Manuel Melendo, and Milagros Bueno. 2023. "Exogenous Application of Indol-3-Acetic Acid and Salicylic Acid Improves Tolerance to Salt Stress in Olive Plantlets (Olea europaea L. Cultivar Picual) in Growth Chamber Environments" Agronomy 13, no. 3: 647. https://doi.org/10.3390/agronomy13030647