
The Effect of Water Stress on the Glucosinolate Content and Profile: A Comparative Study on Roots and Leaves of Brassica oleracea L. Crops
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Morphometric Traits
2.3. Glucosinolates Extraction
2.4. High-Performance Liquid Chromatography (HPLC) Analysis
2.5. Statistical Analysis
3. Results
3.1. Agronomic Trait Analysis
3.2. Comparison between the Total Amount of GLS between Roots and Leaves
3.3. Variation in Individual Glucosinolate in Roots
3.3.1. Aliphatic Glucosinolates
3.3.2. Indolic Glucosinolates
3.3.3. Aromatic Glucosinolates
3.4. Variation of Individual Glucosinolate in Leaves
3.4.1. Aliphatic Glucosinolate
3.4.2. Indolic Glucosinolate
3.4.3. Aromatic Glucosinolates
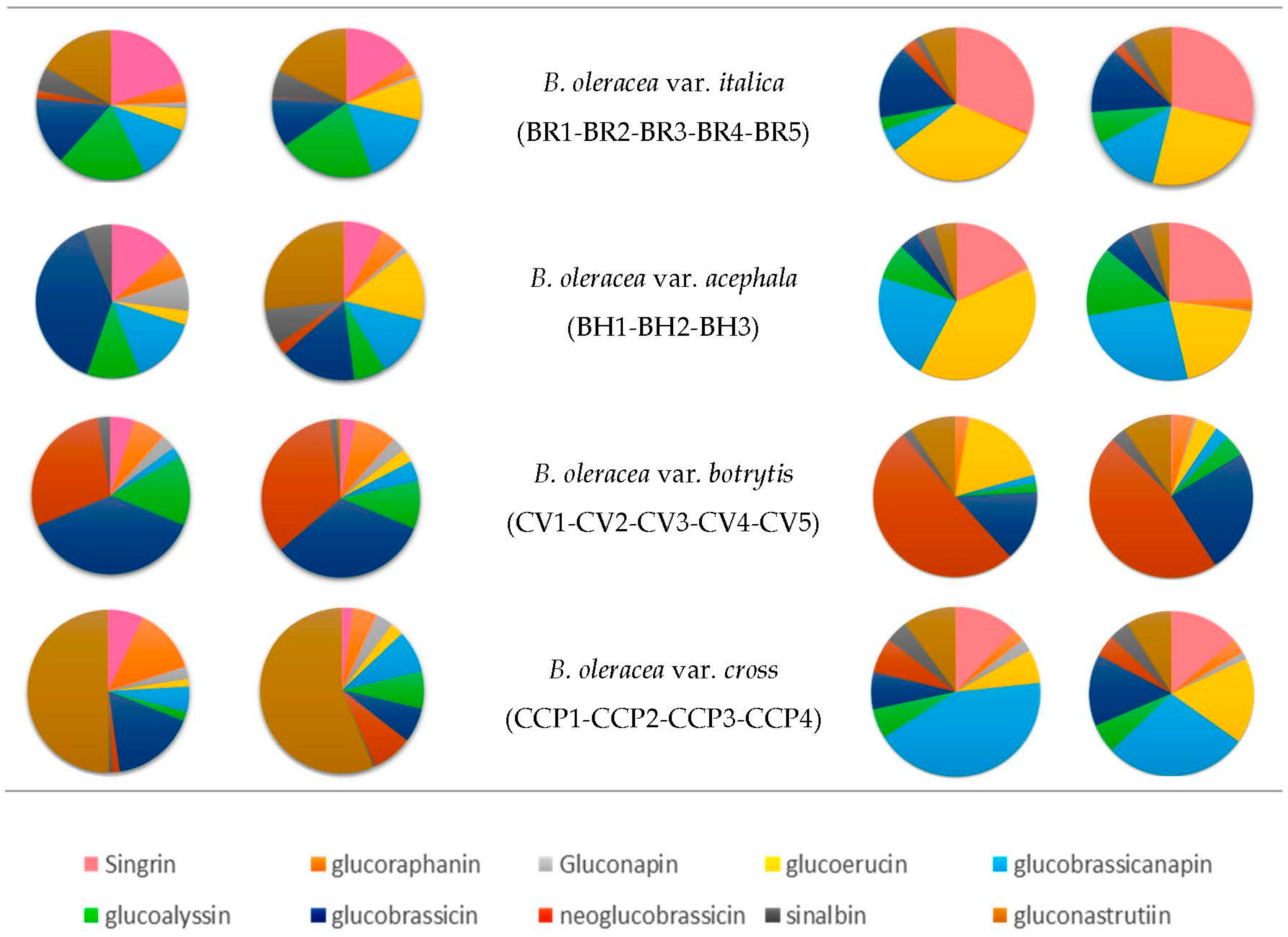
3.5. Chemotaxonomy of the Different Accessions
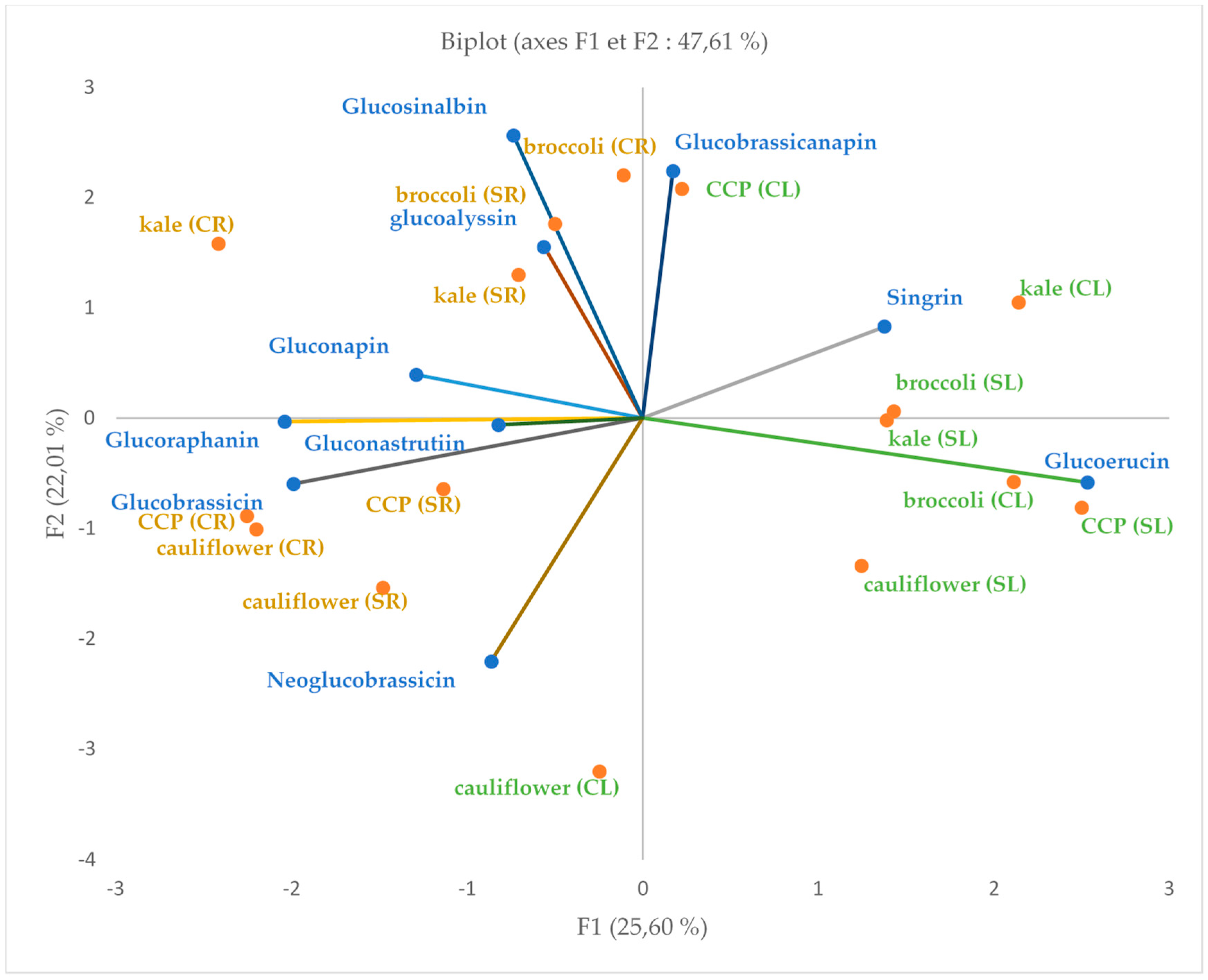
3.6. Principal Component Analysis of Glucosinolate Profile in Leaves and Roots
3.7. Variation of the Glucosinolates Compounds and Profile of the Leaves
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Parkash, V.; Singh, S. A review on potential plant-based water stress indicators for vegetable crops. Sustainability 2020, 12, 3945. [Google Scholar] [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecology 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zittis, G.; Almazroui, M.; Alpert, P.; Ciais, P.; Cramer, W.; Dahdal, Y.; Fnais, M.; Francis, D.; Hadjinicolaou, P.; Howari, F.; et al. Climate change and weather extremes in the Eastern Mediterranean and Middle East. Rev. Geophys. 2022, 60, e2021RG000762. [Google Scholar] [CrossRef]
- Ferreira, C.S.S.; Seifollahi-Aghmiuni, S.; Destouni, G.; Ghajarnia, N.; Kalantari, Z. Soil degradation in the European Mediterranean region: Processes, status and consequences. Sci. Total Environ. 2022, 805, 150106. [Google Scholar] [CrossRef]
- Branca, F.; Chiarenza, G.L.; Cavallaro, C.; Gu, H.; Zhao, Z.; Tribulato, A. Diversity of Sicilian broccoli (Brassica oleracea var. italica) and cauliflower (Brassica oleracea var. botrytis) landraces and their distinctive bio-morphological, antioxidant, and genetic traits. Genet. Resour. Crop. Evol. 2018, 65, 485–502. [Google Scholar] [CrossRef]
- Ben Ammar, H.; Picchi, V.; Arena, D.; Treccarichi, S.; Bianchi, G.; Lo Scalzo, R.; Marghali, S.; Branca, F. Variation of Bio-Morphometric Traits and Antioxidant Compounds of Brassica oleracea L. Accessions in Relation to Drought Stress. Agronomy 2022, 12, 2016. [Google Scholar] [CrossRef]
- Ullah, F.; Bano, A.; Nosheen, A. Effects of plant growth regulators on growth and oil quality of canola (Brassica napus L.) under drought stress. Pak. J. Bot. 2012, 44, 1873–1880. [Google Scholar]
- HongBo, S.; ZongSuo, L.; MingAn, S. Changes of anti-oxidative enzymes and MDA content under soil water deficits among 10 wheat (Triticum aestivum L.) genotypes at maturation stage. Colloids Surf. B Biointerfaces 2005, 45, 7–13. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Khaishany, M.Y.; Al-Qutami, M.A.; Al-Whaibi, M.H.; Grover, A.; Ali, H.M.; Al-Wahibi, M.S.; Bukhari, N.A. Response of different genotypes of faba bean plant to drought stress. Int. J. Mol. Sci. 2015, 16, 10214–10227. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, E.; Ekinci, M.; Turan, M. Impact of biochar in mitigating the negative effect of drought stress on cabbage seedlings. J. Soil Sci. Plant Nutr. 2021, 21, 2297–2309. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Helyes, L. Physiological responses of selected vegetable crop species to water stress. Agronomy 2019, 9, 447. [Google Scholar] [CrossRef] [Green Version]
- Latif, M.; Akram, N.A.; Ashraf, M. Regulation of some biochemical attributes in drought-stressed cauliflower (Brassica oleracea L.) by seed pre-treatment with ascorbic acid. J. Hortic. Sci. Biotechnol. 2016, 91, 129–137. [Google Scholar] [CrossRef]
- La, V.H.; Lee, B.R.; Zhang, Q.; Park, S.H.; Islam, M.; Kim, T.H. Salicylic acid improves drought-stress tolerance by regulating the redox status and proline metabolism in Brassica rapa. Hortic. Environ. Biotechnol. 2019, 60, 31–40. [Google Scholar] [CrossRef]
- Miękus, N.; Marszałek, K.; Podlacha, M.; Iqbal, A.; Puchalski, C.; Świergiel, A.H. Health Benefits of Plant-Derived Sulfur Compounds, Glucosinolates, and Organosulfur Compounds. Molecules 2020, 25, 3804. [Google Scholar] [CrossRef]
- Vik, D.; Mitarai, N.; Wulff, N.; Halkier, B.A.; Burow, M. Dynamic modeling of indole glucosinolate hydrolysis and its impact on auxin signaling. Front. Plant Sci. 2018, 9, 550. [Google Scholar] [CrossRef] [Green Version]
- Jensen, L.M.; Jepsen, H.K.S.; Halkier, B.A.; Kliebenstein, D.J.; Burow, M. Natural variation in cross-talk between glucosinolates and onset of flowering in Arabidopsis. Front. Plant Sci. 2015, 6, 697. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.S.; Ye, W.; Hossain, M.A.; Okuma, E.; Uraji, M.; Nakamura, Y.; Mori, I.; Murata, Y. Glucosinolate degradation products, isothiocyanates, nitriles, and thiocyanates, induce stomatal closure accompanied by peroxidase-mediated reactive oxygen species production in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2013, 77, 120928–120983. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ballesta, M.; Moreno-Fernández, D.A.; Castejón, D.; Ochando, C.; Morandini, P.A.; Carvajal, M. The impact of the absence of aliphatic glucosinolates on water transport under salt stress in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 524. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ballesta, M.d.C.; Muries, B.; Moreno, D.Á.; Dominguez-Perles, R.; García-Viguera, C.; Carvajal, M. Involvement of a glucosinolate (sinigrin) in the regulation of water transport in Brassica oleracea grown under salt stress. Physiol. Plantarum. 2014, 150, 145–160. [Google Scholar] [CrossRef]
- Variyar, P.S.; Banerjee, A.; Akkarakaran, J.J.; Suprasanna, P. Role of Glucosinolates in Plant Stress Tolerance. Emerging Technologies and Management of Crop Stress Tolerance. Acad. Press 2014, 1, 271–291. [Google Scholar] [CrossRef]
- Augustine, R.; Bisht, N. Biofortification of oilseed Brassica juncea with the anti-cancer compound glucoraphanin by suppressing GSL-ALK gene family. Sci. Rep. 2016, 5, 18005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.; Liu, C.; Zheng, H.; Zheng, L. Melatonin treatment affects the glucoraphanin-sulforaphane system in postharvest fresh-cut broccoli (Brassica oleracea L.). Food Chem. 2020, 307, 125562. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.X.; Kim, H.U.; Kim, J.A.; Lim, M.H.; Jin, M.; Lee, S.C.; Hong, J.K.; Park, B.S. Genome-wide identification of glucosinolate synthesis genes in Brassica rapa. FEBS J. 2009, 276, 3559–3574. [Google Scholar] [CrossRef]
- Franzke, A.; Lysak, M.A.; Al-Shehbaz, I.A.; Koch, M.A.; Mummenhoff, K. Cabbage family affairs: The evolutionary history of Brassicaceae. Trends Plant Sci. 2011, 16, 108–116. [Google Scholar] [CrossRef]
- Francisco, M.; Joseph, B.; Caligagan, H.; Li, B.; Corwin, J.A.; Lin, C.; Kerwin, R.; Burow, M.; Kliebenstein, D.J. The defense metabolite, allyl glucosinolate, modulates Arabidopsis thaliana biomass dependent upon the endogenous glucosinolate pathway. Front. Plant Sci. 2016, 7, 774. [Google Scholar] [CrossRef] [Green Version]
- Chhajed, S.; Mostafa, I.; He, Y.; Abou-Hashem, M.; El-Domiaty, M.; Chen, S. Glucosinolate Biosynthesis and the Glucosinolate–MyrosiGSTe System in Plant Defense. Agronomy 2020, 10, 1786. [Google Scholar] [CrossRef]
- Ishida, M.; Hara, M.; Fukino, N.; Kakizaki, T.; Morimitsu, Y. Glucosinolate metabolism, functionality and breeding for the improvement of Brassicaceae vegetables. Breed. Sci. 2014, 64, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Guo, Q.; Gu, Y. Factors Influencing Glucoraphanin and Sulforaphane Formation in Brassica Plants: A Review. J. Integr. Agric. 2012, 11, 1804–1816. [Google Scholar] [CrossRef]
- Van Dam, N.M.; Tytgat, T.O.G.; Kirkegaard, J.A. Root and shoot glucosinolates: A comparison of their diversity, function and interactions in natural and managed ecosystems. Phytochem. Rev. 2009, 8, 171–186. [Google Scholar] [CrossRef] [Green Version]
- Rios, J.J.; Agudelo, A.; Moreno, D.A.; Carvajal, M. Growing broccoli under salinity: The influence of cultivar and season on glucosinolates content. Scientia Agricola. 2020, 77. [Google Scholar] [CrossRef] [Green Version]
- Chorol, S.; Angchok, D.; Stobdan, T. Irrigation timing as a glucosinolate alteration factor in radish (Raphanus sativus L.) (Gya Labuk and Tsentay Labuk) in the Indian Trans-Himalayan region of Ladakh. J. Food Compos. Anal. 2021, 100, 103904. [Google Scholar] [CrossRef]
- Di Gioia, F.; Rosskopf, E.N.; Leonardi, C.; Giuffrida, F. Effects of application timing of saline irrigation water on broccoli production and quality. Agric. Water Manag. 2018, 203, 97–104. [Google Scholar] [CrossRef]
- Velasco, P.; Cartea, M.E.; González, C.; Vilar, M.; Ordás, A. Factors Affecting the Glucosinolate Content of Kale (Brassica oleracea acephala Group). J. Agric. Food Chem. 2007, 55, 955–962. [Google Scholar] [CrossRef]
- Ciska, E.; Martyniak-Przybyszewska, B.; Kozlowska, H. Content of glucosinolates in cruciferous vegetables grown at the same site for two years under different climatic conditions. J. Agric. Food Chem. 2000, 48, 2862–2867. [Google Scholar] [CrossRef] [PubMed]
- Engelen-Eigles, G.; Holden, G.; Cohen, J.D.; Gardner, G. The Effect of Temperature, Photoperiod, and Light Quality on Gluconasturtiin Concentration in Watercress (gluconasturtium officinale R. Br.). J. Agric. Food Chem. 2006, 54, 328–334. [Google Scholar] [CrossRef] [PubMed]
- ISO 9167-1:1992; Rapeseed-Determination of Glucosinolates Content-Part 1: Method Using High-Performance Liquid Chromatography. International Standard Organization: Geneva, Switzerland, 1992; Volume 9167, pp. 1–9.
- Aires, A.; Carvalho, R. Rapid separation of indole glucosinolates in roots of Chinese cabbage (Brassica rapa Subsp. Pekinensis) by high-performance liquid chromatography with diode array detection. Int. J. Anal. Chem. 2017, 2017, 5125329. [Google Scholar] [CrossRef] [Green Version]
- Oguz, M.C.; Aycan, M.; Oguz, E.; Poyraz, I.; Yildiz, M. Drought Stress Tolerance in Plants: Interplay of Molecular, Biochemical and Physiological Responses in Important Development Stages. Physiologia 2022, 2, 180–197. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Issarakraisila, M.; Ma, Q.; Turner, D.W. Photosynthetic and growth responses of juvenile Chinese kale (Brassica oleracea var. alboglabra) and Caisin (Brassica rapa subsp. parachinensis) to waterlogging and water deficit. Sci. Hortic. 2007, 111, 107–113. [Google Scholar] [CrossRef]
- Souza, A.P.; da Silva, A.C.; Tanaka, A.A.; de Souza, M.E.; Pizzatto, M.; Felipe, R.T.A.; Martim, C.C.; Ferneda, B.G.; da Silva, S.G. Yield and water use efficiency of cauliflower under irrigation different levels in tropical climate. Afr. J. Agric. Res. 2018, 13, 1621–1632. [Google Scholar]
- Rosa, E.A.; Heaney, C.A.M.; Portas, C.A.; Fenwick, G.R. Changes in glucosinolate concentrations in Brassica crops (B. oleracea and B. napus) throughout growing seasons. J. Sci. Food Agr. 1996, 71, 237–244. [Google Scholar] [CrossRef]
- Rosa, E.A.S. Daily variation in glucosinolate concentrations in the leaves and roots of cabbage seedlings in two constant temperature regimes. J. Sci. Food Agr. 1997, 73, 364–368. [Google Scholar] [CrossRef]
- Rosa, E.A.S.; Heaney, R.K.; Fenwick, G.R.; Portas, C. Glucosinolates in crop plants. Hort. Rev. 1997, 19, 99–215. [Google Scholar]
- Schreiner, M.; Beyene, B.; Krumbein, A.; Stutzel, H. Ontogenetic changes of 2-propenyl and 3-indolylmethyl glucosinolates in Brassica carinata leaves as affected by water supply. J. Agric. Food Chem. 2009, 57, 7259–7726. [Google Scholar] [CrossRef]
- Khan, M.A.M.; Ulrichs, C.; Mewis, I. Influence of water stress on the glucosinolate profile of Brassica oleracea var. italica and the performance of Brevicoryne brassicae and Myzus persicae. Entomol. Exp. Appl. 2010, 137, 229–236. [Google Scholar] [CrossRef]
- Di Bella, M.C.; Toscano, S.; Arena, D.; Moreno, D.A.; Romano, D.; Branca, F. Effects of growing cycle and genotype on the morphometric properties and glucosinolates amount and profile of sprouts, microgreens, and baby leaves of broccoli (Brassica oleracea L. varr. italica Plenck) and Kale (B. oleracea L. var. acephala DC.). Agronomy 2021, 11, 1685. [Google Scholar] [CrossRef]
- Cartea, M.E.; Di Bella, M.C.; Velasco, P.; Soengas, P.; Toscano, S.; Branca, F. Evaluation of Italian and Spanish Accessions of Brassica rapa L.: Effect of flowering earliness on fresh yield and biological value. Agronomy 2020, 11, 29. [Google Scholar] [CrossRef]
- Wiesner, M.; Hanschen, F.; Schreiner, M.; Glatt, H.; Zrenner, R. Induced Production of 1-Methoxy-indol-3-ylmethyl Glucosinolate by Jasmonic Acid and Methyl Jasmonate in Sprouts and Leaves of Pak Choi (Brassica rapa ssp. chinensis). Int. J. Mol. Sci. 2013, 14, 14996–15016. [Google Scholar] [CrossRef] [Green Version]
- López-Berenguer, C.; Martínez-Ballesta, M.C.; García-Viguera, C.; Carvajal, M. Leaf water balance mediated by aquaporins under salt stress and associated glucosinolate synthesis in broccoli. Plant Sci. 2008, 174, 321–328. [Google Scholar] [CrossRef]
- Pfalz, M.; Mikkelsen, M.D.; Bednarek, P.; Olsen, C.E.; Halkier, B.A.; Kroymann, J. Metabolic engineering in Nicotiana benthamiana reveals key enzyme functions in Arabidopsis indole glucosinolate modification. Plant Cell 2011, 23, 716–729. [Google Scholar] [CrossRef] [Green Version]
- Hornbacher, J.; Horst-Niessen, I.; Herrfurth, C.; Feussner, I.; Papenbrock, J. First experimental evidence suggests use of glucobrassicin as source of auxin in drought-stressed Arabidopsis thaliana. Front. Plant Sci. 2022, 13, 1025969. [Google Scholar] [CrossRef] [PubMed]
- Kushad, M.M.; Brown, A.F.; Kurilich, A.C.; Juvik, J.A.; Klein, B.P.; Wallig, M.A.; Jeffery, E.H. Variation of Glucosinolates in Vegetable Crops of Brassica oleracea. J. Agric. Food Chem. 1999, 47, 1541–1548. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, F.P. Correlation analyses between volatiles and glucosinolates show no evidence for chemical defense signaling in Brassica rapa. Front. Ecol. Evol. 2014, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.S.; Riley, M.B.; Norsworthy, J.K.; Bridges, W., Jr. Glucosinolate profile variation of growth stages of wild radish (Raphanus raphanistrum). J. Agric. Food Chem. 2010, 58, 3309–3315. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zheng, S.; Liu, Y.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Lv, H.; Wang, Y.; Xu, D. Characterization of glucosinolates in 80 broccoli genotypes and different organs using UHPLC-Triple-TOF-MS method. Food Chem. 2021, 334, 127519–127529. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Cheng, B.; Wang, Y.; Liu, G.; Hu, L.; Yu, X.; He, H. Effects of Fresh-cut and Storage on Glucosinolates Profile Using Broccoli as A Case Study. Hortic. Plant. J. 2022, in press. [Google Scholar] [CrossRef]
- Huseby, S.; Koprivova, A.; Lee, B.R.; Saha, S.; Mithen, R.; Wold, A.B.; Bengtsson, G.B.; Kopriva, S. Diurnal and light regulation of sulphur assimilation and glucosinolate biosynthesis in Arabidopsis. J. Exp. Bot. 2013, 64, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef]
- Treccarichi, S.; BenAmmar, H.; Amari, M.; Calì, R.; Tribulato, A.; Branca, F. Molecular Markers for Detecting Inflorescence Size of Brassica oleracea L. Crops and B. oleracea Complex Species (n = 9) Useful for Breeding of Broccoli (B. oleracea var. italica) and Cauliflower (B. oleracea var. botrytis). Plants 2023, 12, 407. [Google Scholar] [CrossRef]
- Treccarichi, S.; Di Gaetano, C.; Di Stefano, F.; Gasparini, M.; Branca, F. Using Simple Sequence Repeats in 9 Brassica Complex Species to Assess Hypertrophic Curd Induction. Agriculture 2021, 11, 622. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Code | Descriptors |
|---|---|
| PW | Plant weight (g) |
| PH | Plant height (cm) |
| SD | Stem diameter (mm) |
| NL | Number of leaves (n) |
| SPAD | SPAD (0–99.9) |
| RW | Root weight (g) |
| MRL | Main root length (cm) |
| SIN | Singrin (µmol. g−1 d.w.) |
| GRA | Glucoraphanin (µmol. g−1 d.w.) |
| GNA | Gluconapin (µmol. g−1 d.w.) |
| GER | Glucoerucin (µmol. g−1 d.w.) |
| GBN | Glucobrassicanapin (µmol. g−1 d.w.) |
| GAL | Glucoalyssin (µmol. g−1 d.w.) |
| GBS | Glucobrassicin (µmol. g−1 d.w.) |
| NGBS | Neoglucobrassicin (µmol. g−1 d.w.) |
| SIB | Sinalbin (µmol. g−1 d.w.) |
| GST | Gluconastrutiin (µmol. g−1 d.w.) |
| GLST | Total GLSs amount (µmol. g−1 d.w.) |
| Genotype | PW (g) | PH (cm) | SD (mm) | NL (n) | SPAD (0 to 99.9) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IRR | NIR | VI | Mean | IRR | NIR | VI | Mean | IRR | NIR | VI | Mean | IRR | NIR | VI | Mean | IRR | NIR | VI | Mean | |
| BH 1 | 300.0 ± 2.0 | 211.0 ± 1.0 | −29.7 | 255.5 ± 1.5 | 69.0 ± 1.0 | 26.8 ± 0.6 | −61.2 | 47.9 ± 0.8 | 2.9 ± 0.0 | 2.3 ± 0.1 | −20.7 | 2.6 ± 0.1 | 8.0 ± 1.0 | 7.0 ± 1.0 | −12.5 | 7.5 ± 1.0 | 59.9 ± 0.3 | 48.4 ± 3.9 | −19.2 | 54.2 ± 2.1 |
| BH 2 | 165.0 ± 13.0 | 145.5 ± 3.5 | −11.8 | 155.3 ± 8.3 | 69.8 ± 1.8 | 30.9 ± 0.7 | −55.7 | 50.4 ± 1.3 | 3.8 ± 0.1 | 2.5 ± 0.2 | −34.2 | 3.2 ± 0.2 | 9.0 ± 1.0 | 6.0 ± 1.0 | −33.3 | 7.5 ± 1.0 | 49.1 ± 3.8 | 35.1 ± 3.4 | −28.5 | 42.1 ± 3.6 |
| BH 3 | 304.5 ± 55.5 | 279.0 ± 31.0 | −8.4 | 291.8 ± 43.3 | 64.0 ± 1.0 | 35.6 ± 0.8 | −44.4 | 49.8 ± 0.9 | 4.1 ± 0.0 | 2.4 ± 0.3 | −41.5 | 3.3 ± 0.2 | 8.0 ± 1.0 | 8.0 ± 1.0 | 0.0 | 8.0 ± 1.0 | 60.1 ± 0.7 | 57.4 ± 2 | −4.5 | 58.8 ± 1.4 |
| BR 1 | 472.5 ± 38.5 | 192.0 ± 50.0 | −59.4 | 332.3 ± 44.3 | 45.5 ± 6.5 | 31.1 ± 8.1 | −31.6 | 38.3 ± 7.3 | 5.1 ± 0.3 | 1.3 ± 0.4 | −74.5 | 3.2 ± 0.4 | 7.0 ± 1.0 | 5.0 ± 1.0 | −28.6 | 6.0 ± 1.0 | 55.1 ± 0.1 | 45.7 ± 6.1 | −17.1 | 50.4 ± 3.1 |
| BR 2 | 369.0 ± 19.0 | 199.0 ± 23.0 | −46.1 | 284.0 ± 21.0 | 42.5 ± 3.5 | 39.9 ± 1.7 | −6.1 | 41.2 ± 2.6 | 4.2 ± 0.6 | 2.1 ± 1.1 | −50.0 | 3.2 ± 0.9 | 10.0 ± 2.0 | 4.0 ± 1.0 | −60.0 | 7.0 ± 1.5 | 57.6 ± 2 | 54.9 ± 2.3 | −4.7 | 56.3 ± 2.2 |
| BR 3 | 535.0 ± 15.0 | 212.0 ± 46.0 | −60.4 | 373.5 ± 30.5 | 42.5 ± 1.5 | 33.6 ± 6.2 | −20.9 | 38.1 ± 3.9 | 3.3 ± 0.2 | 2.4 ± 0.1 | −27.3 | 2.9 ± 0.2 | 12.0 ± 0.0 | 8.0 ± 2.0 | −33.3 | 10.0 ± 1.0 | 61.4 ± 0.4 | 46.4 ± 5 | −24.4 | 53.9 ± 2.7 |
| BR 4 | 508.0 ± 15.0 | 296.0 ± 16.0 | −41.7 | 402.0 ± 15.5 | 49.1 ± 2.1 | 35.8 ± 4.4 | −27.1 | 42.5 ± 3.3 | 4.7 ± 0.0 | 1.7 ± 0.3 | −63.8 | 3.2 ± 0.2 | 13.0 ± 2.0 | 5.0 ± 1.0 | −61.5 | 9.0 ± 1.5 | 56.4 ± 0.5 | 39.5 ± 0.6 | −30.0 | 48.0 ± 0.6 |
| BR 5 | 382.0 ± 42.0 | 288.0 ± 76.0 | −24.6 | 335.0 ± 59.0 | 58.3 ± 3.3 | 45.8 ± 2.8 | −21.4 | 52.1 ± 3.1 | 3.4 ± 0.0 | 2.4 ± 0.1 | −29.4 | 2.9 ± 0.1 | 9.0 ± 1.0 | 7.0 ± 1.0 | −22.2 | 8.0 ± 1.0 | 50.6 ± 9 | 47.4 ± 0.7 | −6.3 | 49.0 ± 4.9 |
| CCP 1 | 426.0 ± 76.0 | 170.0 ± 60.0 | −60.1 | 298.0 ± 68.0 | 50.8 ± 1.8 | 35.5 ± 11.5 | −30.1 | 43.2 ± 6.7 | 4.5 ± 0.6 | 1.9 ± 0.0 | −57.8 | 3.2 ± 0.3 | 8.0 ± 1.0 | 5.0 ± 2.0 | −37.5 | 6.5 ± 1.5 | 55.4 ± 5.5 | 48.2 ± 1.6 | −13.0 | 51.8 ± 3.6 |
| CCP 2 | 452.0 ± 48.0 | 138.0 ± 44.0 | −69.5 | 295.0 ± 46.0 | 48.5 ± 7.5 | 24.2 ± 2.2 | −50.1 | 36.4 ± 4.9 | 4.9 ± 0.1 | 2.8 ± 0.1 | −42.9 | 3.9 ± 0.1 | 11.0 ± 0.0 | 4.0 ± 0.0 | −63.6 | 7.5 ± 0.0 | 54.6 ± 0.9 | 54.6 ± 0.9 | 0.0 | 54.6 ± 0.9 |
| CCP 3 | 300.0 ± 42.0 | 172.0 ± 8.0 | −42.7 | 236.0 ± 25.0 | 36.5 ± 4.0 | 20.9 ± 1.5 | −42.7 | 28.7 ± 2.8 | 4.8 ± 0.8 | 2.7 ± 0.5 | −43.8 | 3.8 ± 0.7 | 10.0 ± 0.0 | 5.0 ± 1.0 | −50.0 | 7.5 ± 0.5 | 62.1 ± 2.3 | 51.1 ± 2.7 | −17.7 | 56.6 ± 2.5 |
| CCP 4 | 192.0 ± 32.0 | 255.5 ± 1.5 | 33.1 | 223.8 ± 16.8 | 37.0 ± 1.0 | 34.6 ± 1.2 | −6.5 | 35.8 ± 1.1 | 4.2 ± 0.5 | 1.3 ± 0.1 | −69.0 | 2.8 ± 0.3 | 7.0 ± 1.0 | 5.0 ± 1.0 | −28.6 | 6.0 ± 1.0 | 47.2 ± 1.6 | 46.8 ± 1.6 | −0.8 | 47 ± 1.6 |
| CV 1 | 413.0 ± 83.0 | 112.0 ± 4.0 | −72.9 | 262.5 ± 43.5 | 47.5 ± 8.5 | 26.2 ± 4.0 | −44.8 | 36.9 ± 6.3 | 3.5 ± 0.4 | 1.7 ± 0.1 | −51.4 | 2.6 ± 0.3 | 10.0 ± 1.0 | 6.0 ± 1.0 | −40.0 | 8.0 ± 1.0 | 56.9 ± 6.0 | 44.0 ± 0.5 | −22.7 | 50.5 ± 3.3 |
| CV 2 | 289.0 ± 48.0 | 177.0 ± 59.0 | −38.8 | 233.0 ± 53.5 | 26.0 ± 3.0 | 13.7 ± 0.7 | −47.3 | 19.9 ± 1.9 | 4.0 ± 0.8 | 2.2 ± 0.1 | −45.0 | 3.1 ± 0.5 | 11.0 ± 3.0 | 5.0 ± 1.0 | −54.5 | 8.0 ± 2.0 | 57.5 ± 1.9 | 44.8 ± 0.2 | −22.1 | 51.2 ± 1.1 |
| CV 3 | 333.0 ± 45.0 | 362.5 ± 17.5 | 8.9 | 347.8 ± 31.3 | 38.0 ± 6.0 | 16.8 ± 5.3 | −55.8 | 27.4 ± 5.7 | 4.4 ± 0.4 | 1.7 ± 0.1 | −61.4 | 3.1 ± 0.3 | 15.0 ± 3.0 | 5.0 ± 2.0 | −66.7 | 10.0 ± 2.5 | 50.9 ± 3.8 | 41.7 ± 2.8 | −18.1 | 46.3 ± 3.3 |
| CV 4 | 401.0 ± 49.0 | 306.5 ± 18.2 | −23.6 | 353.9 ± 33.6 | 44.5 ± 3.5 | 27.6 ± 9.6 | −38.0 | 36.1 ± 6.6 | 4.5 ± 1.2 | 2.4 ± 0.5 | −46.7 | 3.5 ± 0.9 | 6.0 ± 0.0 | 6.0 ± 2.0 | 0.0 | 6.0 ± 1.0 | 60.8 ± 3.9 | 51.7 ± 5.2 | −15.0 | 56.3 ± 4.6 |
| CV 5 | 525.5 ± 99.5 | 231.0 ± 45.0 | −56.0 | 378.3 ± 72.3 | 34.0 ± 2.0 | 16.6 ± 1.4 | −51.2 | 25.3 ± 1.7 | 4.2 ± 0.5 | 2.0 ± 0.4 | −52.4 | 3.1 ± 0.5 | 13.0 ± 0.0 | 6.0 ± 2.0 | −53.8 | 9.5 ± 1.0 | 62.8 ± 5.3 | 43.1 ± 4.6 | −31.4 | 53.0 ± 5.0 |
| Mean | 380.0 ± 102.4 | 214.9 ± 64.5 | 47.3 ± 12.2 | 29.1 ± 8.8 | 4.1 ± 0.6 | 2.1 ± 0.4 | 9.8 ± 2.5 | 5.7 ± 1.2 | 56.4 ± 4.7 | 47.1 ± 5.7 | ||||||||||
| Significancy of the differences by ANOVA Student–Newman–Keuls | ||||||||||||||||||||
| IR | ** | ** | *** | ** | * | |||||||||||||||
| GE | *** | *** | ** | *** | *** | |||||||||||||||
| IR × GE | *** | *** | *** | *** | *** | |||||||||||||||
| Genotype | RW (g) | MRL (cm) | ||||||
|---|---|---|---|---|---|---|---|---|
| IRR | NIR | VI | Mean | IRR | NIR | VI | Mean | |
| BH 1 | 43.6 ± 7.4 | 25.1 ± 3.2 | −42.4 | 34.3 ± 5.3 | 18.6 ± 1.9 | 6.0 ± 0.7 | −67.7 | 12.3 ± 1.3 |
| BH 2 | 14.2 ± 2.5 | 14.1 ± 4.1 | −0.7 | 14.1 ± 3.3 | 21.0 ± 2.2 | 5.4 ± 1.1 | −74.3 | 13.2 ± 1.7 |
| BH 3 | 25.6 ± 7.1 | 15.0 ± 3.0 | −41.4 | 20.3 ± 5.1 | 16.6 ± 2.5 | 2.8 ± 0.5 | −83.1 | 9.7 ± 1.5 |
| BR 1 | 39.0 ± 7.4 | 28.0 ± 2.0 | −28.2 | 33.5 ± 4.7 | 13.6 ± 3.3 | 5.0 ± 0.0 | −63.2 | 9.3 ± 1.7 |
| BR 2 | 30.6 ± 3.5 | 29.0 ± 1.0 | −5.2 | 29.8 ± 2.3 | 11.9 ± 1.2 | 7.6 ± 1.8 | −36.1 | 9.8 ± 1.5 |
| BR 3 | 43.9 ± 5.5 | 15.0 ± 5.0 | −65.8 | 29.5 ± 5.3 | 12.6 ± 1.0 | 5.8 ± 0.5 | −54.0 | 9.2 ± 0.8 |
| BR 4 | 41.7 ± 5.2 | 13.5 ± 3.2 | −67.6 | 27.6 ± 4.2 | 14.7 ± 1.2 | 3.4 ± 0.2 | −76.9 | 9.1 ± 0.7 |
| BR 5 | 31.6 ± 4.1 | 19.0 ± 1.0 | −39.9 | 25.3 ± 2.6 | 17.5 ± 1.6 | 5.6 ± 0.4 | −68.0 | 11.6 ± 1.0 |
| CCP 1 | 35.0 ± 6.0 | 16.0 ± 2.0 | −54.3 | 25.5 ± 4.0 | 15.2 ± 1.3 | 4.5 ± 0.3 | −70.4 | 9.9 ± 0.8 |
| CCP 2 | 37.4 ± 7.9 | 16.0 ± 8.0 | −57.2 | 26.7 ± 8.0 | 14.5 ± 1.9 | 4.3 ± 0.3 | −70.3 | 9.4 ± 1.1 |
| CCP 3 | 24.9 ± 3.2 | 19.0 ± 1.0 | −23.7 | 22.0 ± 2.1 | 10.8 ± 0.7 | 6.0 ± 0.4 | −44.4 | 8.4 ± 0.6 |
| CCP 4 | 16.5 ± 4.2 | 16.0 ± 4.0 | −3.0 | 16.3 ± 4.1 | 10.9 ± 0.9 | 4.1 ± 1.1 | −62.4 | 7.5 ± 1.0 |
| CV 1 | 33.9 ± 6.4 | 28.0 ± 8.0 | −17.4 | 31.0 ± 7.2 | 14.2 ± 4.0 | 5.0 ± 0.3 | −64.8 | 9.6 ± 2.2 |
| CV 2 | 24.5 ± 11.0 | 13.0 ± 3.0 | −46.9 | 18.8 ± 7.0 | 7.6 ± 0.1 | 5.1 ± 1.3 | −32.9 | 6.35 ± 0.7 |
| CV 3 | 27.9 ± 6.5 | 24.0 ± 4.0 | −14.0 | 26.0 ± 5.3 | 11.3 ± 1.2 | 5.5 ± 0.5 | −51.3 | 8.4 ± 0.9 |
| CV 4 | 33.3 ± 7.4 | 23.0 ± 13.0 | −30.9 | 28.2 ± 10.2 | 13.3 ± 1.0 | 5.5 ± 0.3 | −58.6 | 9.4 ± 0.7 |
| CV 5 | 43.5 ± 12.4 | 8.0 ± 0.0 | −81.6 | 25.8 ± 6.2 | 10.1 ± 0.6 | 4.3 ± 0.4 | −57.4 | 7.2 ± 0.5 |
| Mean | 32.2 ± 9.1 | 18.9 ± 6.2 | 13.8 ± 3.4 | 5.0 ± 1.1 | ||||
| Significance of the differences by ANOVA Student–Newman–Keuls | ||||||||
| IR | n.s. | * | ||||||
| GE | *** | *** | ||||||
| IR × GE | *** | *** | ||||||
| Roots | Leaves | |||||
|---|---|---|---|---|---|---|
| Genotypes | IRR | NIR | Mean | IRR | NIR | Mean |
| BH 1 | 5.1 ± 1.0 | 9.7 ± 0.3 | 9.7 ± 3.6 | 122.7 ± 3.0 | 126.0 ± 3.7 | 124.4 ± 2.3 |
| BH 2 | 11.8 ± 0.6 | 38.2 ± 1.6 | 38.2 ± 18.7 | 39.1 ± 2.5 | 82.5 ± 2.4 | 60.8 ± 30.7 |
| BH 3 | 3.5 ± 0.2 | 5.1 ± 0.2 | 5.1 ± 1.1 | 48.4 ± 5.0 | 76.9 ± 1.7 | 62.7 ± 20.2 |
| BR 1 | 4.7 ± 0.6 | 32.2 ± 1.8 | 32.2 ± 19.4 | 39.8 ± 2.8 | 72.7 ± 3.6 | 56.3 ± 23.3 |
| BR 2 | 4.2 ± 0.0 | 6.2 ± 0.0 | 6.2 ± 1.4 | 184.3 ± 14.9 | 264.9 ± 4.2 | 224.6 ± 56.9 |
| BR 3 | 8.8 ± 1.6 | 10.7 ± 0.1 | 10.7 ± 1.3 | 81.6 ± 3.2 | 98.2 ± 0.0 | 89.9 ± 11.7 |
| BR 4 | 28.9 ± 5.4 | 36.1 ± 6.0 | 36.1 ± 1 | 291.4 ± 91 | 578.9 ± 33.5 | 435.2 ± 203.3 |
| BR 5 | 9.2 ± 1.9 | 20.1 ± 1.9 | 20.1 ± 7.7 | 222.2 ± 17.9 | 336.7 ± 51.5 | 279.5 ± 80.9 |
| CCP 1 | 2.5 ± 0.1 | 2.7 ± 0.2 | 2.7 ± 0.1 | 45.5 ± 1.0 | 66.2 ± 2.6 | 55.9 ± 14.7 |
| CCP 2 | 4.3 ± 0.1 | 6.6 ± 0.6 | 6.6 ± 1.6 | 103.9 ± 67.3 | 138.7 ± 16.8 | 121.3 ± 24.6 |
| CCP 3 | 19.6 ± 7.5 | 23.6 ± 5.3 | 23.6 ± 2.8 | 115.3 ± 35.9 | 400.1 ± 10.5 | 257.7 ± 201.4 |
| CCP 4 | 5.2 ± 0.7 | 8.3 ± 2.1 | 8.3 ± 2.2 | 48.9 ± 0.0 | 99.2 ± 4.6 | 74.1 ± 35.6 |
| CV 1 | 29.0 ± 0.1 | 32.1 ± 3.2 | 32.1 ± 2.2 | 111.8 ± 23.7 | 411.6 ± 5.1 | 261.7 ± 211.9 |
| CV 2 | 4.9 ± 1.1 | 18.2 ± 1.2 | 18.2 ± 9.4 | 66.9 ± 7.2 | 78.6 ± 2.9 | 72.8 ± 8.3 |
| CV 3 | 6.1 ± 0.9 | 16.6 ± 2.3 | 16.6 ± 7.4 | 35.8 ± 0.0 | 88.1 ± 0.0 | 62.0 ± 36.9 |
| CV 4 | 10.9 ± 0.4 | 12.8 ± 1.6 | 12.8 ± 1.3 | 39.02 ± 2.4 | 66.9 ± 7.3 | 53.0 ± 19.7 |
| CV 5 | 14.8 ± 1.1 | 15.3 ± 4.4 | 15.3 ± 0.4 | 114.6 ± 0.0 | 245.7 ± 0.0 | 180.2 ± 92.7 |
| Mean | 10.2 ± 8.4 | 8.4 ± 11.4 | 100.6 ± 72.7 | 190.1 ± 156.0 | ||
| Significancy of the differences by ANOVA Student–Newman–Keuls | ||||||
| IR | ** | ** | ||||
| GE | ** | ** | ||||
| IR × GE | ** | *** | ||||
| Correlations | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SIN | GRA | GNA | GER | GBN | GAL | GBS | NGBS | SIB | GST | IW | IH | ISD | IRW | IRL | IL | ISPAD | |
| SIN | 1 | 0.064 | −0.012 | −0.246 | 0.071 | −0.043 | −0.043 | −0.478 ** | 0.342 * | −0.225 | −0.093 | 0.483 ** | 0.208 | −0.151 | −0.271 | 0.150 | 0.064 |
| GRA | 1 | 0.180 | −0.157 | −0.052 | −0.149 | −0.244 | 0.239 | 0.029 | −0.210 | −0.275 | −0.068 | −0.044 | −0.311 | 0.283 | −0.234 | −0.152 | |
| GNA | 1 | −0.223 | −0.151 | −0.112 | −0.125 | −0.036 | −0.089 | −0.199 | −0.197 | −0.066 | −0.307 | −0.037 | −0.062 | −0.014 | −0.051 | ||
| GER | 1 | −0.214 | −0.459 ** | −0.202 | −0.279 | −0.353 * | 0.503 ** | −0.325 | −0.240 | 0.228 | −0.187 | −0.208 | −0.019 | −0.200 | |||
| GBN | 1 | 0.302 | −0.253 | −0.223 | 0.394 * | 0.134 | 0.204 | 0.128 | −0.056 | 0.460 ** | 0.088 | −0.006 | 0.215 | ||||
| GAL | 1 | 0.083 | −0.041 | 0.284 | 0.067 | 0.314 | 0.224 | −0.289 | 0.362 * | −0.101 | 0.132 | 0.338 | |||||
| GBS | 1 | −0.127 | 0.335 | −0.137 | 0.347 * | −0.090 | 0.075 | 0.144 | −0.258 | 0.373 * | 0.230 | ||||||
| NGBS | 1 | −0.165 | −0.293 | 0.089 | −0.361 * | −0.128 | −0.188 | 0.431* | −0.363 * | −0.239 | |||||||
| SIB | 1 | −0.211 | 0.321 | 0.215 | 0.118 | 0.336 | −0.151 | 0.259 | 0.449 ** | ||||||||
| GST | 1 | −0.252 | −0.115 | −0.118 | −0.103 | −0.170 | −0.212 | −0.122 | |||||||||
| IW | 1 | −0.146 | 0.142 | 0.415 * | −0.136 | 0.373 * | 0.116 | ||||||||||
| IH | 1 | −0.297 | 0.146 | 0.162 | 0.016 | 0.410 * | |||||||||||
| ISD | 1 | −0.170 | −0.064 | 0.330 | −0.050 | ||||||||||||
| IRW | 1 | 0.184 | 0.105 | 0.312 | |||||||||||||
| IRL | 1 | −0.434 * | −0.064 | ||||||||||||||
| IL | 1 | 0.204 | |||||||||||||||
| ISPAD | 1 | ||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Ammar, H.; Arena, D.; Treccarichi, S.; Di Bella, M.C.; Marghali, S.; Ficcadenti, N.; Lo Scalzo, R.; Branca, F. The Effect of Water Stress on the Glucosinolate Content and Profile: A Comparative Study on Roots and Leaves of Brassica oleracea L. Crops. Agronomy 2023, 13, 579. https://doi.org/10.3390/agronomy13020579
Ben Ammar H, Arena D, Treccarichi S, Di Bella MC, Marghali S, Ficcadenti N, Lo Scalzo R, Branca F. The Effect of Water Stress on the Glucosinolate Content and Profile: A Comparative Study on Roots and Leaves of Brassica oleracea L. Crops. Agronomy. 2023; 13(2):579. https://doi.org/10.3390/agronomy13020579
Chicago/Turabian StyleBen Ammar, Hajer, Donata Arena, Simone Treccarichi, Maria Concetta Di Bella, Sonia Marghali, Nadia Ficcadenti, Roberto Lo Scalzo, and Ferdinando Branca. 2023. "The Effect of Water Stress on the Glucosinolate Content and Profile: A Comparative Study on Roots and Leaves of Brassica oleracea L. Crops" Agronomy 13, no. 2: 579. https://doi.org/10.3390/agronomy13020579