
The Current Distribution of Carex alatauensis in the Qinghai–Tibet Plateau Estimated by MaxEnt
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
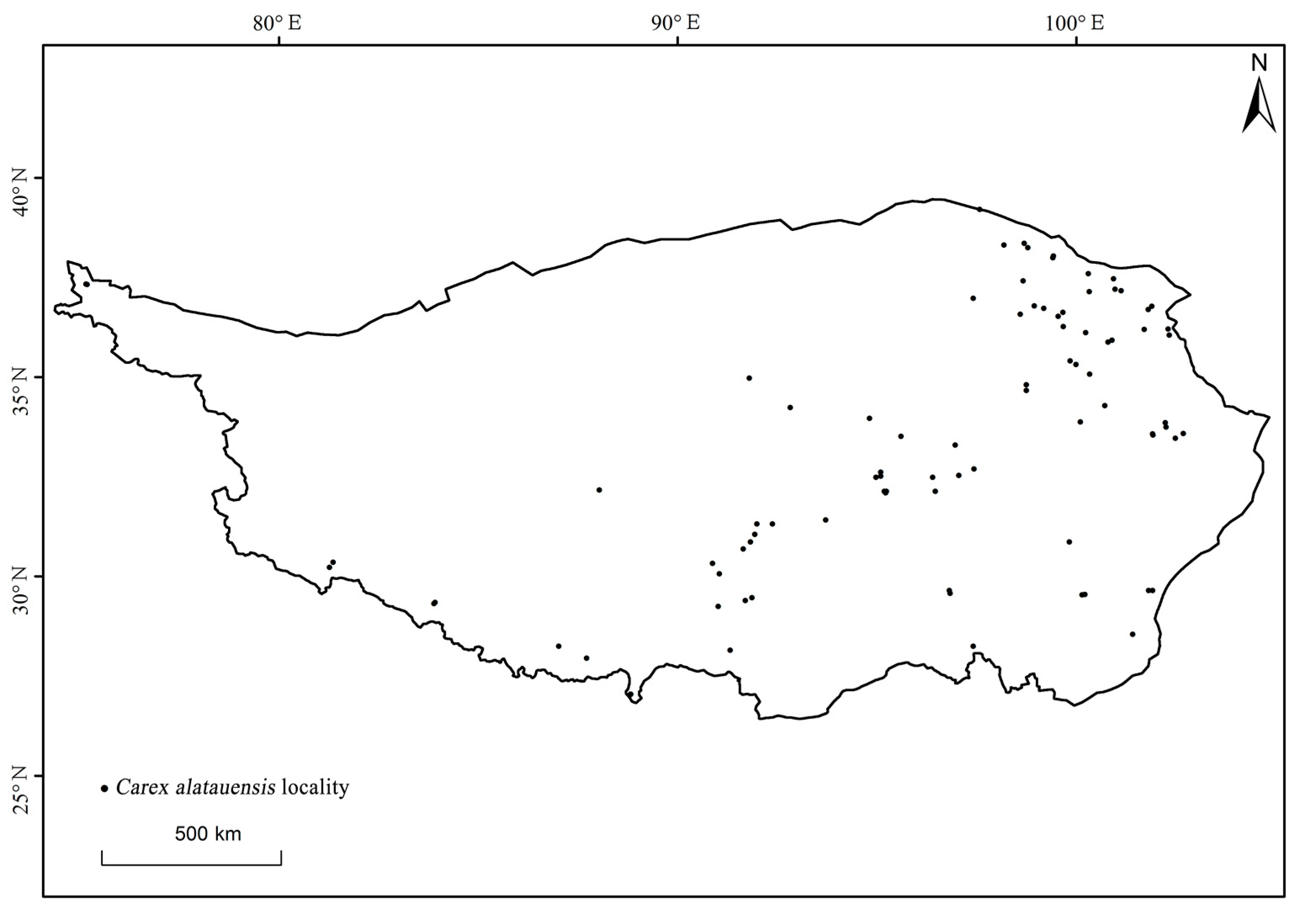
2.2. Carex alatauensis Distribution Data
2.3. Climatic Data
2.4. Maximum Entropy Niche-Based Model
3. Results
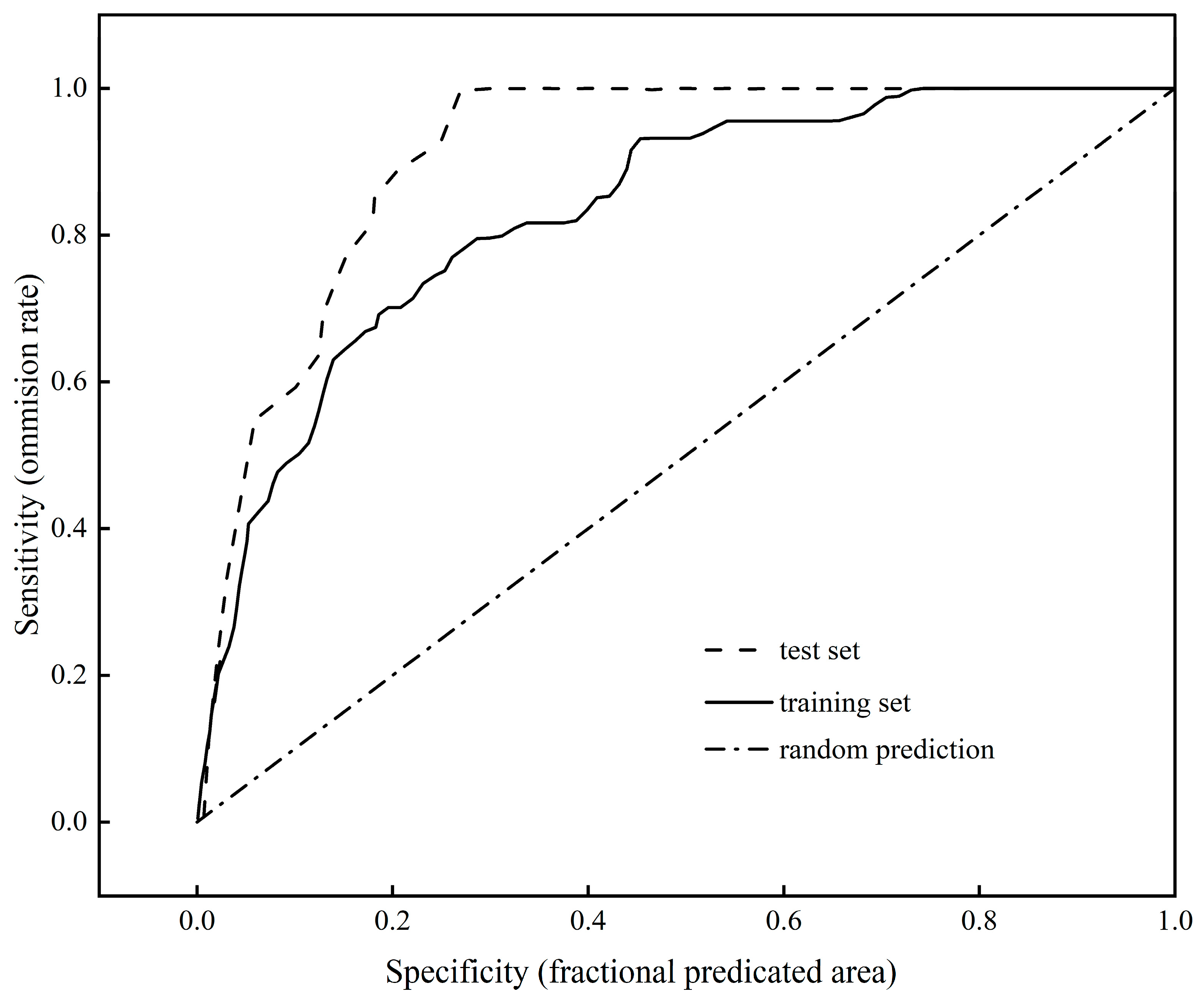
3.1. Accuracy of the Maxent Model
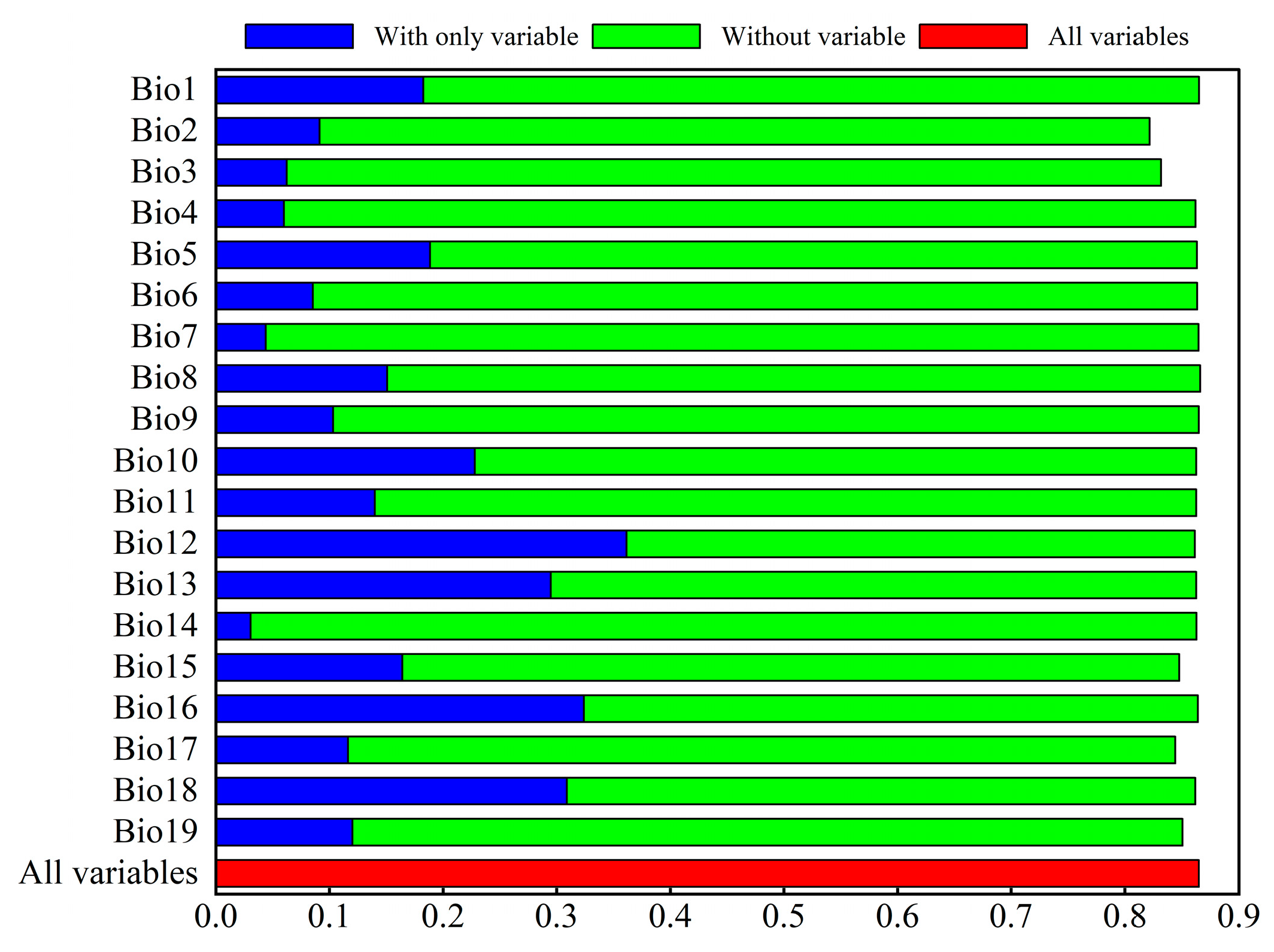
3.2. The Variables’ Contribution to Suitability
3.3. The Variables’ Response to Suitability
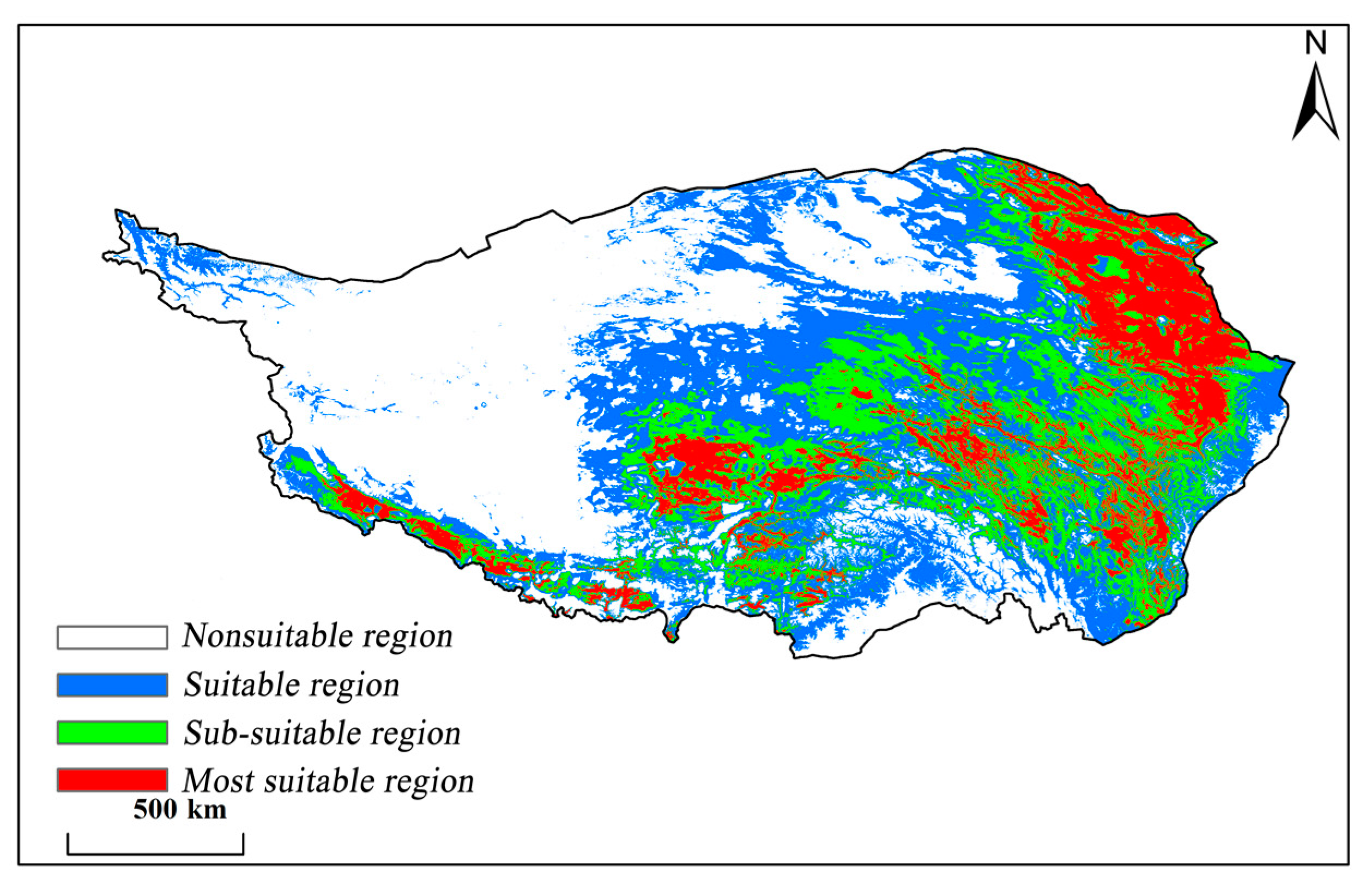
3.4. Potential Distribution of Carex alatauensis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, G.; Wu, T. Responses of permafrost to climate change and their environmental significance, Qinghai-Tibet Plateau. J. Geophys. Res. Atmos. 2007, 112, F02S03. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.B. Rangeland degradation on the Qinghai-Tibetan plateau: A review of the evidence of its magnitude and causes. J. Arid. Environ. 2010, 74, 1–12. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, B.; Zhang, D. A discussion on the boundary and area of the Tibetan Plateau in China. Geogr. Res. 2002, 21, 1–8. [Google Scholar] [CrossRef]
- Lu, Q.; Wu, S.; Zhao, D. Variations in Alpine Grassland Cover and Its Correlation with Climate Variables on the Qinghai-Tibet Plateau in 1982-2013. Sci. Geogr. Sin. 2017, 37, 292–300. [Google Scholar] [CrossRef]
- Liao, G.; Jia, Y. Rangeland Resources of China; China Science and Technology Press: Beijing, China, 1996; pp. 489–492. [Google Scholar]
- Raiesi, F.; Riahi, M. The influence of grazing exclosure on soil C stocks and dynamics, and ecological indicators in upland arid and semi-arid rangelands. Ecol. Indic. 2014, 41, 145–154. [Google Scholar] [CrossRef]
- Luan, J.; Cui, L.; Xiang, C.; Wu, J.; Song, H.; Ma, Q.; Hu, Z. Different grazing removal exclosures effects on soil C stocks among alpine ecosystems in east Qinghai–Tibet Plateau. Ecol. Eng. 2014, 64, 262–268. [Google Scholar] [CrossRef]
- Borer, E.T.; Seabloom, E.W.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Lind, E.M.; Adler, P.B.; Alberti, J.; Anderson, T.M.; Bakker, J.D.; et al. Herbivores and nutrients control grassland plant diversity via light limitation. Nature 2014, 508, 517–520. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Wu, H.; Qin, J.; Lin, C.; Tang, W.; Chen, Y. Recent climate changes over the Tibetan Plateau and their impacts on energy and water cycle: A review. Glob. Planet. Chang. 2014, 112, 79–91. [Google Scholar] [CrossRef]
- Wen, L.; Dong, S.; Li, Y.; Wang, X.; Li, X.; Shi, J.; Dong, Q. The impact of land degradation on the C pools in alpine grasslands of the Qinghai-Tibet Plateau. Plant and Soil 2013, 368, 329–340. [Google Scholar] [CrossRef]
- Cui, X.; Graf, H.-F. Recent land cover changes on the Tibetan Plateau: A review. Clim. Chang. 2009, 94, 47–61. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Dai, L.; Tang, Y.; Li, P. Reipublicae Popularis Sinicae, Tomus 12; Science Press: Beijing, China, 2000; pp. 35–41. [Google Scholar]
- Liu, W.; Wang, F.; Yang, X.; Liu, Y.; Feng, B.; Yu, Y.; Zhang, C.; Cao, Q.; Dong, Q. Effects of the Traits of Reproductive and Vegetative Branches of Kobresia humilis under Different Herbivore Assemblage Grazing in Alpine Grassland. Acta Agrestia Sin. 2022, 30, 2231–2238. [Google Scholar] [CrossRef]
- Wang, S. Vegetation degradation and protection strategy in the “Three rivers fountainhead”area in the Qinghai province. Acta Prataculturae Sin. 2003, 19, 1–9. [Google Scholar]
- Schleuss, P.-M.; Heitkamp, F.; Sun, Y.; Miehe, G.; Xu, X.; Kuzyakov, Y. Nitrogen Uptake in an Alpine Kobresia Pasture on the Tibetan Plateau: Localization by 15N Labeling and Implications for a Vulnerable Ecosystem. Ecosystems 2015, 18, 946–957. [Google Scholar] [CrossRef] [Green Version]
- Ingrisch, J.; Biermann, T.; Seeber, E.; Leipold, T.; Li, M.; Ma, Y.; Xu, X.; Miehe, G.; Guggenberger, G.; Foken, T.; et al. Carbon pools and fluxes in a Tibetan alpine Kobresia pygmaea pasture partitioned by coupled eddy-covariance measurements and 13CO2 pulse labeling. Sci. Total. Environ. 2015, 505, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Miehe, G.; Schleuss, P.-M.; Seeber, E.; Babel, W.; Biermann, T.; Braendle, M.; Chen, F.; Coners, H.; Foken, T.; Gerken, T.; et al. The Kobresia pygmaea ecosystem of the Tibetan highlands—Origin, functioning and degradation of the world’s largest pastoral alpine ecosystem: Kobresia pastures of Tibet. Sci. Total. Environ. 2019, 648, 754–771. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Lei, X.; Gao, W.; Guo, H.; Xie, Y.; Fu, L.; Lei, Y.; Li, Y.; Zhang, Z.; Tang, S. Mapping the potential distribution suitability of 16 tree species under climate change in northeastern China using Maxent modelling. J. For. Res. 2022, 33, 1739–1750. [Google Scholar] [CrossRef]
- Kaky, E.; Nolan, V.; Alatawi, A.; Gilbert, F. A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: A case study with Egyptian medicinal plants. Ecol. Inform. 2020, 60, 101150. [Google Scholar] [CrossRef]
- Yan, X.; Wang, S.; Duan, Y.; Han, J.; Huang, D.; Zhou, J. Current and future distribution of the deciduous shrub Hydrangea macrophylla in China estimated by MaxEnt. Ecol. Evol. 2021, 11, 16099–16112. [Google Scholar] [CrossRef]
- Hu, Z.; Guo, K.; Jin, S.; Pan, H. The influence of climatic changes on distribution pattern of six typical Kobresia species in Tibetan Plateau based on MaxEnt model and geographic information system. Theor. Appl. Clim. 2019, 135, 375–390. [Google Scholar] [CrossRef]
- Jian, N. A Simulation of Biomes on the Tibetan Plateau and Their Responses to Global Climate Change. Mt. Res. Dev. 2000, 20, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Yue, P.P.; Lu, X.F.; Ye, R.R.; Zhang, C.X.; Yang, S.B.; Zhou, Y.B.; Peng, M. Distribution of Stipa purpurea steppe in the Northeastern Qinghai-Xizang Plateau (China). Russ. J. Ecol. 2011, 42, 50–56. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Elith, J.H.; Graham, C.P.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.J.; Hijmans, R.; Huettmann, F.; R. Leathwick, J.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Rae, D.A.; Armbruster, W.S.; Edwards, M.E.; Svengård-Barre, M. Influence of microclimate and species interactions on the composition of plant and invertebrate communities in alpine northern Norway. Acta Oecologica 2006, 29, 266–282. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, G.; Yi, S.; Chen, S.; Wu, Z.; Guan, J.; Zhao, C.; Zhao, Q.; Ye, B. Effects of environmental factors on the distribution of plant communities in a semi-arid region of the Qinghai-Tibet Plateau. Ecol. Res. 2012, 27, 667–675. [Google Scholar] [CrossRef]
- Li, J.; Zhang, F.; Lin, L.; Li, H.; Du, Y.; Li, Y.; Cao, G. Response of the plant community and soil water status to alpine Kobresia meadow degradation gradients on the Qinghai–Tibetan Plateau, China. Ecol. Res. 2015, 30, 589–596. [Google Scholar] [CrossRef]
- Li, L. Study on the Suitable Distribution Mechanism and Response of the Kobresia Meadow in the Three-Rivers Source Region. Master’s Thesis, Lanzhou University, Lanzhou, China, 2017. [Google Scholar]
- Kennedy, A.D. Water as a Limiting Factor in the Antarctic Terrestrial Environment: A Biogeographical Synthesis. Arct. Alp. Res. 1993, 25, 308–315. [Google Scholar] [CrossRef]
- Wu, L.; Wang, M.; Ouyang, H.; Cheng, S.; Song, M. Spatial Distribution Modelling of Kobresia pygmaea (Cyperaceae) on the Qinghai-Tibetan Plateau. J. Resour. Ecol. 2017, 8, 20–29. [Google Scholar] [CrossRef]
- Boyce, R.L.; Clark, R.; Dawson, C. Factors Determining Alpine Species Distribution on Goliath Peak, Front Range, Colorado, U.S.A. Arctic, Antarct. Alp. Res. 2005, 37, 88–96. [Google Scholar] [CrossRef]
- Zhou, Y.; Fan, J.; Zhang, W.; Harris, W.; Zhong, H.; Hu, Z.; Song, L. Factors influencing altitudinal patterns of C3 plant foliar carbon isotope composition of grasslands on the Qinghai-Tibet Plateau, China. Alp. Bot. 2011, 121, 79. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Quiroz, C.L.; Molina-Montenegro, M.A. Facilitation of the non-native Taraxacum officinale by native nurse cushion species in the high Andes of central Chile: Are there differences between nurses? Funct. Ecol. 2008, 22, 148–156. [Google Scholar] [CrossRef]
- Liu, J.; Duan, Y.; Hao, G.; Ge, X.; Sun, H. Evolutionary history and underlying adaptation of alpine plants on the Qinghai–Tibet Plateau. J. Syst. Evol. 2014, 52, 241–249. [Google Scholar] [CrossRef]
- Song, M.; Duan, D.; Chen, H.; Hu, Q.; Zhang, F.; Xu, X.; Tian, Y.; Ouyang, H.; Peng, C. Leaf δ13C reflects ecosystem patterns and responses of alpine plants to the environments on the Tibetan Plateau. Ecography 2008, 31, 499–508. [Google Scholar] [CrossRef]
- Liang, E.; Lu, X.; Ren, P.; Li, X.; Zhu, L.; Eckstein, D. Annual increments of juniper dwarf shrubs above the tree line on the central Tibetan Plateau: A useful climatic proxy. Ann. Bot. 2012, 109, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Liang, E.; Dawadi, B.; Pederson, N.; Eckstein, D. Is the growth of birch at the upper timberline in the Himalayas limited by moisture or by temperature? Ecology 2014, 95, 2453–2465. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, H.; Levy, J.K. Spatiotemporal vegetation cover variations in the Qinghai-Tibet Plateau under global climate change. Sci. Bull. 2008, 53, 915–922. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Li, X.; Kong, X.; Ma, L.; Hu, X.; Yang, Y. Transcriptome analysis reveals diversified adaptation of Stipa purpurea along a drought gradient on the Tibetan Plateau. Funct. Integr. Genom. 2015, 15, 295–307. [Google Scholar] [CrossRef]
- Genxu, W.; Ju, Q.; Guodong, C.; Yuanmin, L. Soil organic carbon pool of grassland soils on the Qinghai-Tibetan Plateau and its global implication. Sci. Total. Environ. 2002, 291, 207–217. [Google Scholar] [CrossRef]
- Cao, X.; Wang, J.; Lu, S.; Zhang, X. Simulation of the potential distribution patterns of Picea crassifolia in climate change scenarios based on the maximum entropy (Maxent) model. Acta Ecol. Sin. 2019, 39, 5232–5240. [Google Scholar] [CrossRef]
- Zhang, X.; Jiang, Y.; Bi, Y.; Liu, X.; Li, X.; Sun, T.; Chen, H.; Li, J. Identification of potential distribution area for Hippophae rhamnoides subsp. sinensis by the MaxEnt mode. Acta Ecol. Sin. 2022, 42, 1420–1428. [Google Scholar] [CrossRef]
- Ma, Y.; Lang, B.; Li, Q.; Shi, J.; Dong, Q. Study on rehabilitating and rebuilding technologies for degenerated alpine meadow in the Changjiang and Yellow river source region. Acta Prataculturae Sin. 2002, 19, 1–5. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description |
|---|---|
| Bio1 | Mean annual temperature |
| Bio2 | Mean diurnal range |
| Bio3 | Isothermality |
| Bio4 | Temperature seasonality |
| Bio5 | Max temperature of the warmest month |
| Bio6 | Minimum temperature of the coldest month |
| Bio7 | Temperature annual range |
| Bio8 | Mean temperature of the wettest quarter |
| Bio9 | Mean temperature of the driest quarter |
| Bio10 | Mean temperature of the warmest quarter |
| Bio11 | Mean temperature of the coldest quarter |
| Bio12 | Annual precipitation |
| Bio13 | Precipitation of the wettest month |
| Bio14 | Precipitation of the driest month |
| Bio15 | Precipitation seasonality |
| Bio16 | Precipitation of the wettest quarter |
| Bio17 | Precipitation of the driest quarter |
| Bio18 | Precipitation of the warmest quarter |
| Bio19 | Precipitation of the coldest quarter |
| Variables | Percent Contribution/% |
|---|---|
| Bio12 | 42.31 ± 3.41 |
| Bio10 | 14.63 ± 5.49 |
| Bio2 | 12.62 ± 1.23 |
| Bio14 | 7.38 ± 0.87 |
| Bio5 | 5.17 ± 4.45 |
| Bio3 | 5.07 ± 1.61 |
| Bio15 | 3.57 ± 1.08 |
| Bio18 | 3.52 ± 2.28 |
| Bio17 | 2.10 ± 0.95 |
| Bio7 | 1.48 ± 1.05 |
| Bio4 | 0.82 ± 0.49 |
| Bio19 | 0.62 ± 0.16 |
| Bio16 | 0.29 ± 0.18 |
| Bio1 | 0.17 ± 0.21 |
| Bio6 | 0.11 ± 0.14 |
| Bio11 | 0.06 ± 0.15 |
| Bio9 | 0.05 ± 0.05 |
| Bio8 | 0 |
| Bio13 | 0 |
| Classification | Simulation Area (km2) |
|---|---|
| Most suitable region | 37.41 × 105 |
| Sub-suitable region | 62.84 × 105 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, Y.; Shi, J.; De, K.; Wang, X.; Wang, W.; Ma, Y.; Zhang, H.; He, M.; Liu, Q. The Current Distribution of Carex alatauensis in the Qinghai–Tibet Plateau Estimated by MaxEnt. Agronomy 2023, 13, 564. https://doi.org/10.3390/agronomy13020564
Xing Y, Shi J, De K, Wang X, Wang W, Ma Y, Zhang H, He M, Liu Q. The Current Distribution of Carex alatauensis in the Qinghai–Tibet Plateau Estimated by MaxEnt. Agronomy. 2023; 13(2):564. https://doi.org/10.3390/agronomy13020564
Chicago/Turabian StyleXing, Yunfei, Jianjun Shi, Kejia De, Xiaoli Wang, Wei Wang, Yuan Ma, Hairong Zhang, Miaohua He, and Qingqing Liu. 2023. "The Current Distribution of Carex alatauensis in the Qinghai–Tibet Plateau Estimated by MaxEnt" Agronomy 13, no. 2: 564. https://doi.org/10.3390/agronomy13020564