
Impact of Futuristic Climate Variables on Weed Biology and Herbicidal Efficacy: A Review

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Impact of Climate Change on Weed Biology
2.1. Photosynthetic Cycles
2.1.1. C3 Plants
2.1.2. C4 Plants
2.2. Basic Principles of Weeds’ Response to an Increased CO2 and Temperature Climate
2.3. Weed Response to Elevated Atmospheric CO2 Concentration
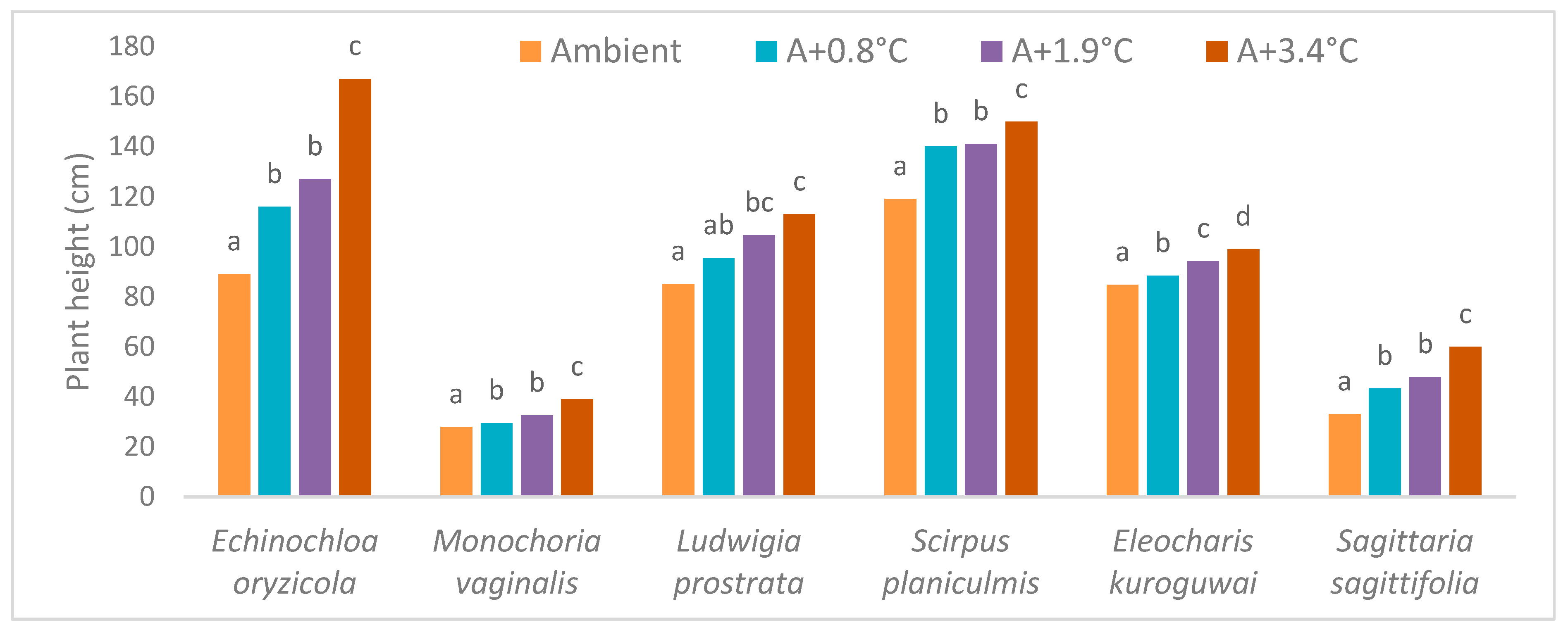
2.4. Weed Response to Increased Temperature
2.5. Weed Response to Changed Rainfall Pattern and Drought Spells
2.6. Weed Response to the Interaction of Climatic Variables
3. Changing Climate and Efficacy of Herbicides
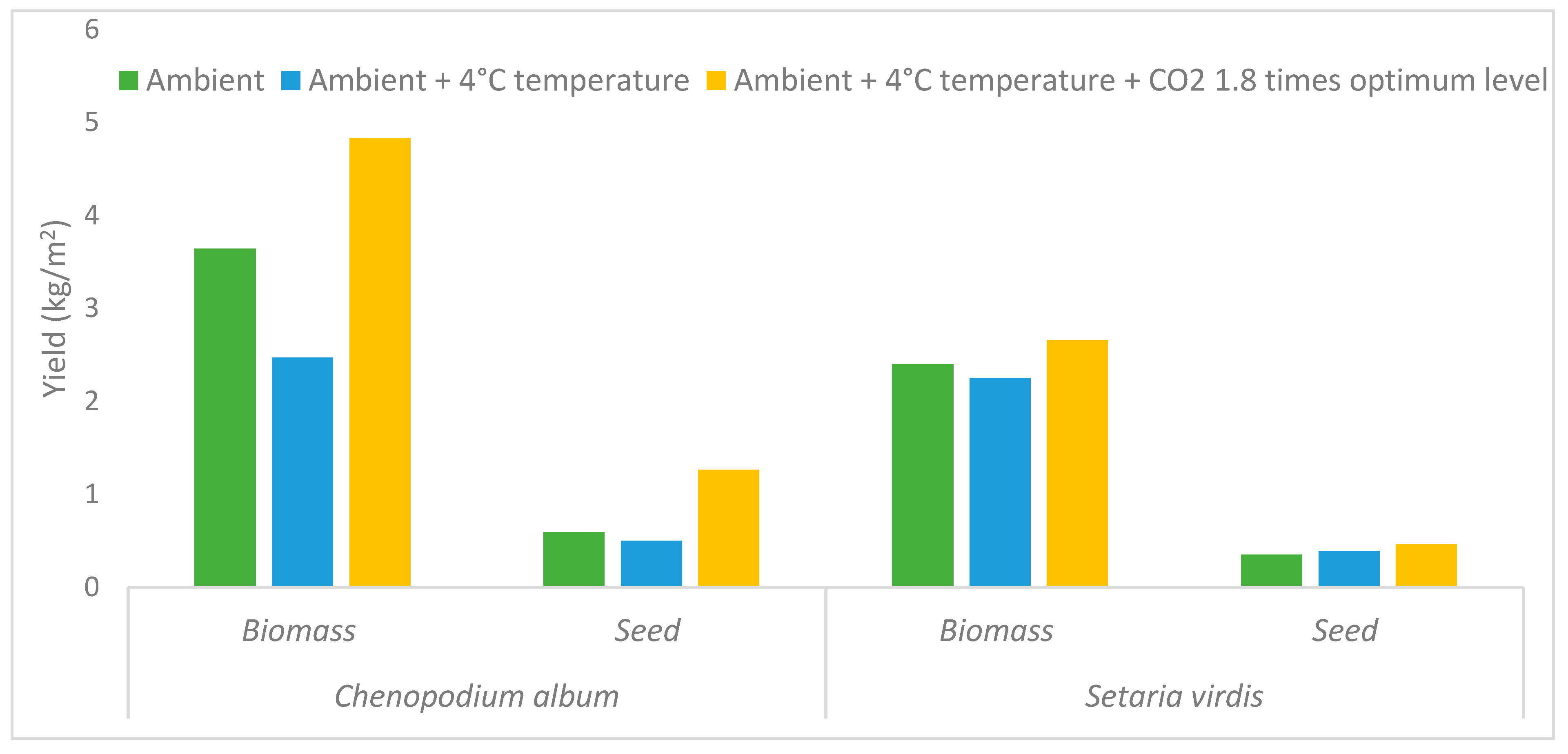
3.1. Increased CO2 Levels and Herbicide Efficacy
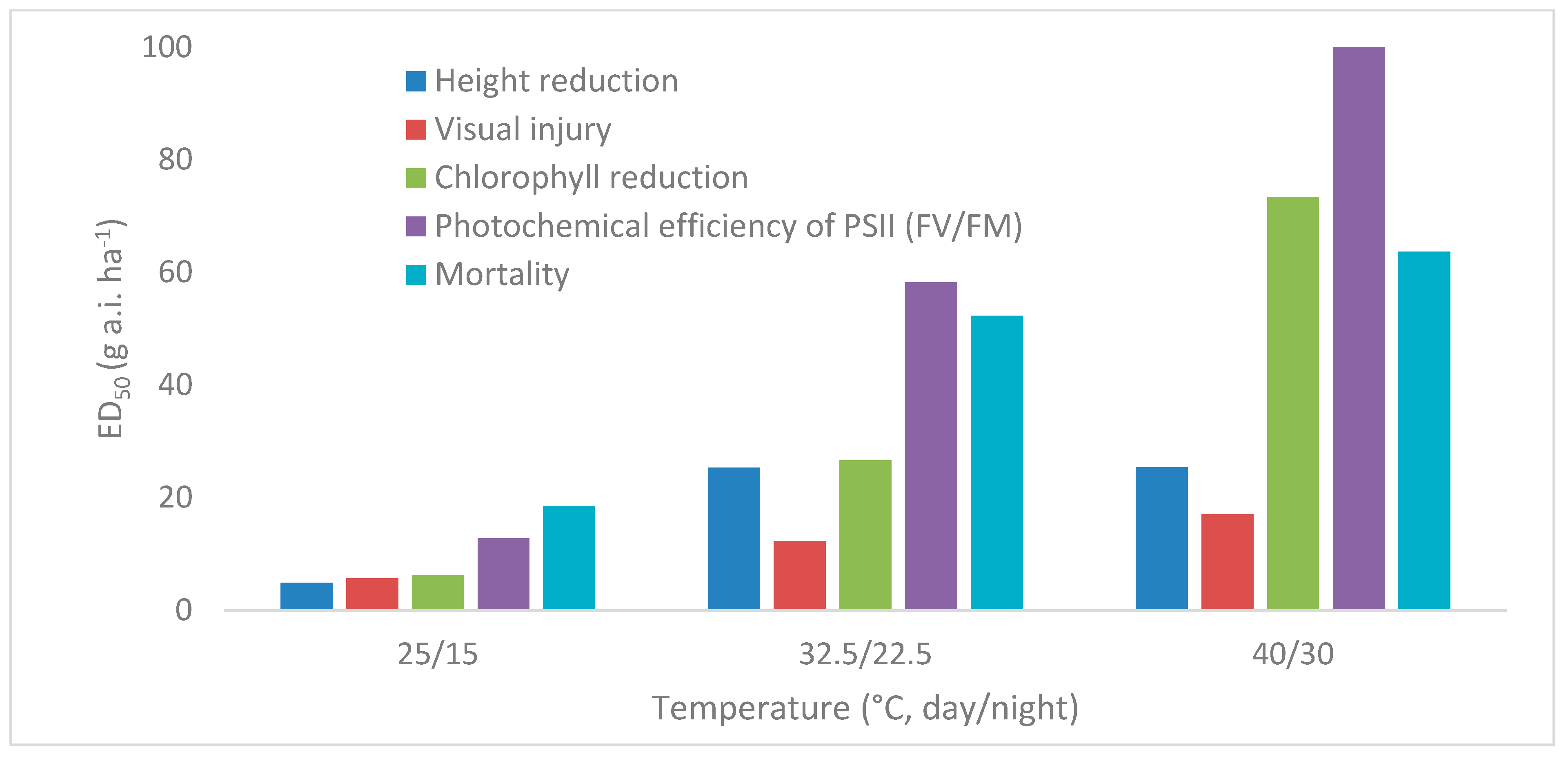
3.2. Temperature and Herbicide Efficacy
3.3. Precipitation, Soil Moisture, and Herbicide Efficacy
3.4. Interaction of Climatic Variables and Herbicide Efficacy
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- IPCC. Working Group I Contribution to the Intergovernmental Panel on Climate Change (IPCC Fifth) Assessment Report Climate Change. The Physical Science Basis, Summary for Policymakers. 2013. Available online: www.climatechange2013.org/images/uploads/WGIAR5SPM_Approved27Sep2013.pdf (accessed on 20 October 2021).
- Coumou, D.; Rahmstorf, S. A decade of weather extremes. Nat. Clim. Chang. 2012, 2, 491–496. [Google Scholar] [CrossRef]
- Gillett, N.P.; Arora, V.K.; Zickfeld, K. Ongoing climate change following a complete cessation of carbon dioxide emissions. Nat. Geosci. 2011, 4, 83–87. [Google Scholar] [CrossRef]
- Robinson, T.M.; Gross, K.L. The impact of altered precipitation variability on annual weed species. Am. J. Bot. 2010, 97, 1625–1629. [Google Scholar] [CrossRef] [PubMed]
- Tubiello, F.N.; Soussana, J.F.; Howden, S.M. Crop and pasture response to climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 19686–19690. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, B.S.; Prabhjyot-Kaur; Mahajan, G.; Randhawa, R.K.; Singh, H.; Kang, M.S. Global Warming and Its Possible Impact on Agriculture in India. Adv. Agron. 2014, 123, 65–121. [Google Scholar] [CrossRef]
- Ziska, L.H.; Blumenthal, D.M.; Franks, S.J. Understanding the nexus of rising CO2, climate change, and evolution in weed biology. Invasive Plant Sci. Manag. 2019, 12, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Das, T.K. Weed Science: Basics and Applications; Jain Brothers: New Delhi, India, 2008. [Google Scholar]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Rana, K.S.; Choudhary, A.K.; Sepat, S. Advances in Field Crop Production; Post Graduate School, IARI: New Delhi, India, 2014; p. 475. [Google Scholar]
- Ziska, L.H. Evaluation of yield loss in field sorghum from a C3 and C4 weed with increasing CO2. Weed Sci. 2003, 51, 14–18. [Google Scholar] [CrossRef]
- Singh, R.P.; Singh, R.K.; Singh, M.K. Impact of climate and carbon dioxide change on weeds and their management—A Review. Indian J. Weed Sci. 2011, 43, 1–11. [Google Scholar]
- Gianessi, L.P. The increasing importance of herbicides in worldwide crop production. Pest Manag. Sci. 2013, 69, 1099–1105. [Google Scholar] [CrossRef]
- Varanasi, A.; Prasad, P.V.V.; Jugulam, M. Impact of climate change factors on weeds and herbicide efficacy. Adv. Agron. 2016, 135, 107–146. [Google Scholar]
- Jordan, D.B.; Ogren, W.L. The CO2/O2 specificity of ribulose 1,5-bisphosphate carboxylase/oxygenase. Planta 1984, 161, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Leegood, R.C. C4 photosynthesis: Principles of CO2 concentration and prospects for its introduction into C3 plants. J. Exp. Bot. 2002, 53, 581–590. [Google Scholar] [CrossRef]
- Hatch, M.D. C4 photosynthesis: A unique elend of modified biochemistry, anatomy and ultrastructure. Biochim. Biophys. Acta (BBA) Rev. Bioenerg. 1987, 895, 81–106. [Google Scholar] [CrossRef]
- Fernando, N.; Manalil, S.; Florentine, S.K.; Chauhan, B.S.; Seneweera, S. Glyphosate Resistance of C3 and C4 Weeds under Rising Atmospheric CO2. Front. Plant Sci. 2016, 7, 910. [Google Scholar] [CrossRef] [Green Version]
- Woodward, F.I.; Cramer, W. Plant functional types and climatic change: Introduction. J. Veg. Sci. 1996, 7, 306–308. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Pautasso, M.; Dehnen-Schmutz, K.; Holdenrieder, O. Plant health and global change—Some implications for landscape management. Biol. Rev. 2010, 85, 729–755. [Google Scholar] [CrossRef] [PubMed]
- Kubisch, A.; Degen, T.; Hovestadt, T.; Poethke, H.J. Predicting range shifts under global change: The balance between local adaptation and dispersal. Ecography 2013, 36, 873–882. [Google Scholar] [CrossRef]
- Kumar, V.; Bana, R.S.; Singh, T.; Kumar, H.; Louhar, G. Ecological weed management approaches for wheat under rice–wheat cropping system. Environ. Sustain. 2021, 4, 51–61. [Google Scholar] [CrossRef]
- Pearman, P.B.; Guisan, A.; Broennimann, O.; Randin, C.F. Niche dynamics in space and time. Trends Ecol. Evol. 2008, 23, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P.; Hodgson, J.G. Botanical contributions to contemporary ecological theory. New Phytol. 1987, 106, 283–295. [Google Scholar] [CrossRef]
- Carroll, S.P.; Hendry, A.P.; Reznick, D.N.; Fox, C.W. Evolution on ecological time-scales. Funct. Ecol. 2007, 21, 387–393. [Google Scholar] [CrossRef]
- Harlan, J.R.; de Wet, J.M.J. Some thoughts about weeds. Econ. Bot. 1965, 19, 16–24. [Google Scholar] [CrossRef]
- Tungate, K.D.; Israel, D.W.; Watson, D.M.; Rufty, T.W. Potential changes in weed competitiveness in an agroecological system with elevated temperatures. Environ. Exp. Bot. 2007, 60, 42–49. [Google Scholar] [CrossRef]
- Patterson, D.T. Weeds in a Changing Climate. Weed Sci. 1995, 43, 685–700. [Google Scholar] [CrossRef]
- Navie, S.C.; Panetta, F.D.; McFadyen, R.E.; Adkins, S.W. The effect of CO2 enrichment on the growth of a C3 weed (Parthenium hysterophorus L.) and its competitive interaction with a C4 grass (Cenchrus ciliaris L.). Plant Protect. Q. 2005, 20, 61–66. [Google Scholar]
- Madhu, M.; Hatfield, J.L. Dynamics of Plant Root Growth under Increased Atmospheric Carbon Dioxide; USDA-ARS/UNL Faculty: Lincoln, NE, USA, 2013; p. 1352. [Google Scholar]
- Ziska, L.H.; Faulkner, S.; Lydon, J. Changes in biomass and root: Shoot ratio of field grown Canada thistle (Cirsium arvense), a noxious, invasive weed, with elevated CO2: Implications for control with glyphosate. Weed Sci. 2004, 52, 584–588. [Google Scholar] [CrossRef]
- Ziska, L.H. Could recent increases in atmospheric CO2 have acted as a selection factor in Avena fatua populations? A case study of cultivated and wild oat competition. Weed Res. 2017, 57, 399–405. [Google Scholar] [CrossRef]
- Awasthi, J.P.; Paraste, K.S.; Rathore, M.; Varun, M.; Jaggi, D.; Kumar, B. Effect of elevated CO2 on Vigna radiata and two weed species: Yield, physiology and crop–weed interaction. Crop. Pasture Sci. 2018, 69, 617–631. [Google Scholar] [CrossRef]
- Ziska, L.H. Observed changes in soyabean growth and seed yield from Abutilon theophrasti competition as a function of carbon dioxide concentration. Weed Res. 2013, 53, 140–145. [Google Scholar] [CrossRef]
- Manea, A.; Leishman, M.R.; Downey, P.O. Exotic C4 Grasses Have Increased Tolerance to Glyphosate under Elevated Carbon Dioxide. Weed Sci. 2011, 59, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Pilipavicius, V. Influence of Climate Change on Weed Vegetation. In Global Warming-Causes, Impacts and Remedies; IntechOpen: London, UK, 2015. [Google Scholar]
- Valerio, M.; Tomecek, M.; Lovelli, S.; Ziska, L.H. Quantifying the effect of drought on carbon dioxide-induced changes in competition between a C3 crop (tomato) and a C4 weed (Amaranthus retroflexus). Weed Res. 2011, 51, 591–600. [Google Scholar] [CrossRef]
- Patterson, D.T.; Westbrook, J.K.; Joyce, R.J.C.; Lingren, P.D.; Rogasik, J. Weeds, insects and diseases. Clim. Change 1999, 43, 711–727. [Google Scholar] [CrossRef]
- Morgan, J.A.; LeCain, D.R.; Mosier, A.R.; Milchunas, D.G. Elevated CO2 enhances water relations and productivity and affects gas exchange in C3 and C4 grasses of the Colorado shortgrass steppe. Glob. Chang. Biol. 2001, 7, 451–466. [Google Scholar] [CrossRef] [Green Version]
- Clements, D.R.; Ditommaso, A. Climate change and weed adaptation: Can evolution of invasive plants lead to greater range expansion than forecasted? Weed Res. 2011, 51, 227–240. [Google Scholar] [CrossRef]
- Weber, E.; Gut, D. A survey of weeds that are increasingly spreading in Europe. Agron. Sustain. Dev. 2005, 25, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Hanzlik, K.; Gerowitt, B. Occurrence and distribution of important weed species in German winter oilseed rape fields. J. Plant Dis. Protect. 2012, 119, 107–120. [Google Scholar] [CrossRef]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Cavero, J.; Zaragoza, C.; Suso, M.L.; Pardo, A. Competition between maize and Datura stramonium in an irrigated field under semi-arid conditions. Weed Res. 1999, 39, 225–240. [Google Scholar] [CrossRef]
- Chadha, A.; Florentine, S.; Javaid, M.; Welgama, A.; Turville, C. Influence of elements of climate change on the growth and fecundity of Datura stramonium. Environ. Sci. Pollut. Res. 2020, 27, 35859–35869. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S. Combined effect of elevated CO2 and temperature on the growth and phenology of two annual C3 and C4 weedy species. Agric. Ecosyst. Environ. 2011, 140, 484–491. [Google Scholar] [CrossRef]
- Bir, M.S.H.; Hien, L.T.; Won Ok, J.; Aung, B.B.; Ruziev, F.; Umurzokov, M.; Jia, W.; Khaitov, B.; Park, K.W. Growth response of weed species in a paddy field under elevated temperatures. Weed Turf Sci. 2018, 7, 321–329. [Google Scholar]
- Rodenburg, J.; Riches, C.R.; Kayeke, J.M. Addressing current and future problems of parasitic weeds in rice. Crop. Protect. 2010, 29, 210–221. [Google Scholar] [CrossRef]
- Patterson, D.T.; Highsmith, M.T.; Flint, E.P. Effects of temperature and CO2 concentration on the growth of cotton (Gossypium hirsutum), spurred anoda (Anoda cristata), and velvetleaf (Abutilon theophrasti). Weed Sci. 1988, 36, 751–757. [Google Scholar] [CrossRef]
- Mortensen, D.A.; Coble, H.D. The Influence of Soil Water Content on Common Cocklebur (Xanthium strumarium) Interference in Soybeans (Glycine max). Weed Sci. 1989, 37, 76–83. [Google Scholar] [CrossRef]
- Donald, W.W.; Khan, M. Yield Loss Assessment for Spring Wheat (Triticum aestivum) Infested with Canada Thistle (Cirsium arvense). Weed Sci. 1992, 40, 590–598. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Abugho, S.B. Effect of Water Stress on the Growth and Development of Amaranthus spinosus, Leptochloa chinensis, and Rice. Am. J. Plant Sci. 2013, 4, 989–998. [Google Scholar] [CrossRef] [Green Version]
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. (Eds.) Climate Change: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC Summary for Policymakers; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Valerio, M.; Tomecek, M.; Lovelli, S.; Ziska, L. Assessing the impact of increasing carbon dioxide and temperature on crop-weed interactions for tomato and a C3 and C4 weed species. Eur. J. Agron. 2013, 50, 60–65. [Google Scholar] [CrossRef]
- Alberto, A.M.; Ziska, L.H.; Cervancia, C.R.; Manalo, P.A. The influence of increasing carbon dioxide and temperature on competitive interactions between a C4 crop, rice (Oryza sativa), and a C3 weed (Echinochloa glabrescens). Aust. J. Plant Physiol. 1996, 23, 793–802. [Google Scholar] [CrossRef]
- Carlson, R.W.; Bazzaz, F.A. The effects of elevated CO2 concentrations on growth, photosynthesis, transpiration and water use efficiency of plants. In Environmental and Climatic Impact of Coal Utilization; Singh, J.J., Deepak, A., Eds.; Academic Press: New York, NY, USA, 1980; pp. 610–622. [Google Scholar]
- Patterson, D.T. Responses of soybean (Glycine max) and three C4 grass weeds to CO2 enrichment during drought. Weed Sci. 1986, 34, 203–210. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Summary for Policymakers; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Travlos, I.; Gazoulis, I.; Kanatas, P.; Tsekoura, A.; Zannopoulos, S.; Papastylianou, P. Key Factors Affecting Weed Seeds’ Germination, Weed Emergence, and Their Possible Role for the Efficacy of False Seedbed Technique as Weed Management Practice. Front. Agron. 2020, 2, 1. [Google Scholar] [CrossRef]
- Jaganathan, G.K.; Liu, B. Role of seed sowing time and microclimate on germination and seedling establishment of Dodonaea viscosa (Sapindaceae) in a seasonal dry tropical environment—An insight into restoration efforts. Botany 2015, 93, 23–29. [Google Scholar] [CrossRef]
- Ooi, M.K.J.; Denham, A.J.; Santana, V.M.; Auld, T.D. Temperature thresholds of physically dormant seeds and plant functional response to fire: Variation among species and relative impact of climate change. Ecol. Evol. 2014, 4, 656–671. [Google Scholar] [CrossRef]
- Zhu, C.; Zeng, Q.; Ziska, L.H.; Zhu, J.; Xie, Z.; Liu, G. Effect of nitrogen supply on carbon dioxide induced changes in competition between rice and barnyard grass (Echinochloa crusgalli). Weed Sci. 2008, 56, 66–71. [Google Scholar] [CrossRef]
- Heap, I.M. The International Survey of Herbicide Resistant Weeds. 2021. Available online: http://www.weedscience.org/ (accessed on 15 August 2021).
- Bunce, J.A. Growth, survival, competition, and canopy carbon dioxide and water vapor exchange of first year alfalfa at an elevated CO2 concentration. Photosynthetica 1993, 29, 557–565. [Google Scholar]
- Alizade, S.; Keshtkar, E.; Mokhtassi-Bidgoli, A.; Sasanfar, H.; Streibig, J.C. Effect of drought stress on herbicide perfor-mance and photosynthetic activity of Avena sterilis subsp. ludoviciana (winter wild oat) and Hordeum spontaneum (wild barley). Weed Res. 2021, 61, 288–297. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Nowak, R.S.; Ellsworth, D.S.; Smith, S.D. Functional responses of plants to elevated atmospheric CO2: Do photosynthetic and productivity data from FACE experiments support early predictions? New Phytol. 2004, 162, 253–280. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H.; Teasdale, J.R. Sustained growth and increased tolerance to glyphosate observed in a C3 perennial weed, quackgrass (Elytrigia repens), grown at elevated carbon dioxide. Funct. Plant Biol. 2000, 27, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Loladze, I. Hidden shift of the ionome of plants exposed to elevated CO2 depletes minerals at the base of human nutrition. eLife 2014, 3, e02245. [Google Scholar] [CrossRef]
- Taub, D.R.; Miller, B.; Allen, H. Effects of elevated CO2 on the protein concentration of food crops: A meta-analysis. Glob. Chang. Biol. 2008, 14, 565–575. [Google Scholar] [CrossRef]
- Ziska, L.H.; Teasdale, J.R.; Bunce, J.A. Future atmospheric carbon dioxide may increase tolerance to glyphosate. Weed Sci. 1999, 47, 608–615. [Google Scholar] [CrossRef]
- Archambault, D.J.; Li, X.; Robinson, D.; Donovan, J.T.O.; Klein, K.K. The Effects of Elevated CO2 and Temperature on Herbicide Efficacy and Weed/Crop Competition. Available online: https://www.parc.ca/project/the-effects-of-elevated-co2-and-temperature-on-herbicide-efficacy-and-weed-crop-competition/ (accessed on 16 December 2022).
- Refatti, J.P.; de Avila, L.A.; Camargo, E.R.; Ziska, L.H.; Oliveira, C.; Salas-Perez, R.; Rouse, C.E.; Roma-Burgos, N. High CO2 and Temperature Increase Resistance to Cyhalofop-Butyl in Multiple-Resistant Echinochloa colona. Front. Plant Sci. 2019, 10, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, C.E. The effect of environment on foliage uptake and translocation of herbicides. In Aspects of Applied Biology 4: Influence of Environmental Factors on Herbicide Performance and Crop and Weed Biology; Biologists, A.O.A., Ed.; The Association of Applied Biologists: Warwick, UK, 1983; pp. 157–169. [Google Scholar]
- Johnson, B.C.; Young, B.G. Influence of temperature and relative humidity on the foliar activity of mesotrione. Weed Sci. 2002, 50, 157–161. [Google Scholar] [CrossRef]
- Pline, W.A.; Wu, J.; Hatzios, K.K. Effects of Temperature and Chemical Additives on the Response of Transgenic Herbicide-Resistant Soybeans to Glufosinate and Glyphosate Applications. Pestic. Biochem. Physiol. 1999, 65, 119–131. [Google Scholar] [CrossRef]
- Godar, A.S.; Varanasi, V.K.; Nakka, S.; Vara Prasad, P.V.; Thompson, C.R.; Mithila, J. Physiological and Molecular Mechanisms of Differential Sensitivity of Palmer Amaranth (Amaranthus palmeri) to Mesotrione at Varying Growth Temperatures. PLoS ONE 2015, 10, e0126731. [Google Scholar] [CrossRef] [Green Version]
- Matzrafi, M.; Seiwert, B.; Reemtsma, T.; Rubin, B.; Peleg, Z. Climate change increases the risk of herbicide-resistant weeds due to enhanced detoxification. Planta 2016, 244, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Burgos, N.R.; Rouse, C.E.; Saski, C.A. Concerted enrichment of gene networks in multiple resistant Echinochloa colona. In Proceedings of the Annual Meeting; Weed Science Society of America: Arlington, VA, USA, 2018; p. 387. [Google Scholar]
- Rouse, C.E.; Burgos, N.R.; Saski, C.A. Co-evolution of abiotic stress adaptation and quinclorac resistance in Echinochloa colona. In Proceedings of the Annual Meeting; Weed Science Society of America: Arlington, VA, USA, 2018; p. 386. [Google Scholar]
- Ganie, Z.A.; Jugulam, M.; Jhala, A.J. Temperature influences efficacy, absorption, and translocation of 2,4-D or glyphosate in glyphosate-resistant and glyphosate-susceptible common ragweed (Ambrosia artemisiifolia) and giant ragweed (Ambrosia trifida). Weed Sci. 2017, 65, 588–602. [Google Scholar] [CrossRef]
- Devine, M.D.; Duke, S.O.; Fedtke, C. Foliar absorption of herbicides. In Physiology of Herbicide Action; Prentice-Hall: Englewood Cliffs, NJ, USA, 1993; pp. 29–52. [Google Scholar]
- Strachan, S.D.; Casini, M.S.; Heldreth, K.M.; Scocas, J.A.; Nissen, S.J.; Bukun, B.; Lindenmayer, R.B.; Shaner, D.L.; Westra, P.; Brunk, G. Vapor Movement of Synthetic Auxin Herbicides: Aminocyclopyrachlor, Aminocyclopyrachlor-Methyl Ester, Dicamba, and Aminopyralid. Weed Sci. 2010, 58, 103–108. [Google Scholar] [CrossRef]
- Behrens, R.; Lueschen, W.E. Dicamba volatility. Weed Sci. 1979, 27, 486–493. [Google Scholar] [CrossRef]
- Dong, X.; Sun, H. Effect of temperature and moisture on degradation of herbicide atrazine in agricultural soil. Int. J. Environ. Agric. Res. 2016, 2, 150–157. [Google Scholar]
- Long, Y.H.; Li, R.T.; Wu, X.M. Degradation of S-metolachlor in soil as affected by environmental factors. J. Soil Sci. Plant Nutr. 2014, 14, 189–198. [Google Scholar] [CrossRef]
- Taylor-Lovell, S.; Sims, G.K.; Wax, L.M. Effects of Moisture, Temperature, and Biological Activity on the Degradation of Isoxaflutole in Soil. J. Agric. Food Chem. 2002, 50, 5626–5633. [Google Scholar] [CrossRef] [PubMed]
- Atienza, J.; Tabernero, M.T.; Álvarez-Benedí, J.; Sanz, M. Volatilisation of triallate as affected by soil texture and air velocity. Chemosphere 2001, 42, 257–261. [Google Scholar] [CrossRef]
- Reinhardt, C.F.; Nel, P.C. The influence of soil type, soil water content and temperature on atrazine persistence. S. Afr. J. Plant Soil 1993, 10, 45–49. [Google Scholar] [CrossRef]
- Jugulam, M.; Varanasi, A.K.; Varanasi, V.K.; Vara Prasad, P.V. Climate change influence on herbicide efficacy and weed management. In Food Security and Climate Change; Yadav, S.S., Redden, R.J., Hatfield, J.L., Ebert, A.W., Hunter, D., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2019; Chapter 18; pp. 433–448. [Google Scholar]
- Blackshaw, R.E.; Moyer, J.R.; Kozub, G.C. Efficacy of downy brome herbicides as influenced by soil properties. Can. J. Plant Sci. 1994, 74, 177–183. [Google Scholar] [CrossRef]
- Zaller, J.G.; Weber, M.; Maderthaner, M.; Gruber, E.; Takács, E.; Mörtl, M.; Klátyik, S.; Győri, J.; Römbke, J.; Leisch, F.; et al. Effects of glyphosate-based herbicides and their active ingredients on earthworms, water infiltration and glyphosate leaching are influenced by soil properties. Environ. Sci. Eur. 2021, 33, 51. [Google Scholar] [CrossRef]
- Keikotlhaile, B.M. Influence of the Processing Factors on Pesticide Residues in Fruits and Vegetables and Its Application in Consumer Risk Assessment. Ph.D. Thesis, Ghent University, Ghent, Belgium, 2011. [Google Scholar]
- Zanatta, J.F.; Procópio, S.O.; Manica, R.; Pauletto, E.A.; Cargnelutti Filho, A.; Vargas, L.; Sganzerla, D.C.; Rosenthal, M.D.A.; Pinto, J.J.O. Teores de água no solo e eficácia do herbicida fomesafen no controle de Amaranthus hybridus. Planta Daninha 2008, 26, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Tao, B.; Messersmith, C.G.; Nalewaja, J.D. Glyphosate Efficacy on Velvetleaf (Abutilon theophrasti) is Affected by Stress. Weed Sci. 2007, 55, 240–244. [Google Scholar] [CrossRef]
- Pereira, M.R.R. Efeito de herbicidas sobre plantas de Brachiaria plantaginea submetidas a estresse hídrico. Planta Daninha 2010, 28, 1047–1058. [Google Scholar] [CrossRef] [Green Version]
- Jursík, M.; Kočárek, M.; Hamouzová, K.; Soukup, J.; Venclova, V. Effect of precipitation on the dissipation, efficacy and selectivity of three chloroacetamide herbicides in sunflower. Plant Soil Environ. 2013, 59, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Dao, T.H.; Lavy, T.L. Extraction of Soil Solution Using a Simple Centrifugation Method for Pesticide Adsorption-Desorption Studies. Soil Sci. Soc. Am. J. 1978, 42, 375–377. [Google Scholar] [CrossRef]
- Moyer, J.R. Effects of soil moisture on the efficacy and selectivity of soil-applied herbicides. Weed Sci. 1987, 3, 19–34. [Google Scholar]
- Amare, T. Review on Impact of Climate Change on Weed and Their Management. Am. J. Biol. Environ. Stat. 2016, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H.; Bunce, J.A. Plant responses to rising carbon dioxide. In Plant Growth and Climate Change; Morison, J.I.L., Morecroft, M.D., Eds.; Blackwell Publishing: Oxford, UK, 2006; pp. 17–47. [Google Scholar]
- Matzrafi, M.; Brunharo, C.; Tehranchian, P.; Hanson, B.D.; Jasieniuk, M. Increased temperatures and elevated CO2 levels reduce the sensitivity of Conyza canadensis and Chenopodium album to glyphosate. Sci. Rep. 2019, 9, 2228. [Google Scholar] [CrossRef] [Green Version]
- Stopps, G.J.; Nurse, R.E.; Sikkema, P.H. The effect of time of day on the activity of post-emergence soybean herbicides. Weed Technol. 2013, 27, 690–695. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | C3 Plants | C4 Plants |
|---|---|---|
| CO2 acceptor | Ribulose-1,5-bisphosaphate | Phosphoenolpyruvate |
| Enzyme | Rubisco | PEP carboxylase |
| Extent of photorespiration | High | Negligible |
| Saturation light intensity | 1000–4000 foot candle | Hardly reaches saturation even in full sunlight |
| CO2 compensation point | 50–150 ppm | 0–10 ppm |
| Operating CO2 fixation pathway | Only C3 pathway | Both C3 and C4 pathway |
| Optimum temperature for growth | 10–25 °C | 30–45 °C |
| Preferred climatic condition | Dominant in temperate regions | Dominant in tropical and sub-tropical areas |
| Abundance | Found in both angiosperms and gymnosperms | Found only in angiosperms |
| C3 Weed Species | C4 Weed Species |
|---|---|
| Avena fatua | Amaranthus viridis |
| Abuliton theophrasti | Boerhavia diffusa |
| Ageratum conyzoides | Cynodon dactylon |
| Bidens pilosa | Cyperus rotundus |
| Chenopodium album | Dactyloctenium aegypticum |
| Cirsium arvense | Digitaria sanguinalis |
| Commelina benghalensis | Echinochloa crus-galli |
| Convolvulus arvensis | Elusine indica |
| Eichhornia crassipes | Fimbristylis miliacea |
| Phalaris minor | Ischaemun rugosum |
| Striga asiatica | Portulaca oleracea |
| Rumex dentatus | Sorghum halpense |
| Xanthium strumarium | Trianthema portulacastrum |
| C3 Species | Biomass (% of Ambient) | C4 Species | Biomass (% of Ambient) |
|---|---|---|---|
| Ambrosia artemisiifolia | 110–133 | Amaranthus retroflexus | 96–141 |
| Datura stramonium | 174–272 | Cyperus rotundus | 102 |
| Elymus repens | 164 | Digitaria ciliaris | 106–121 |
| Rumex acetosella | 131 | Echinochloa crusgalli | 95–159 |
| Cirsium arvense | 121 | Elusine indica | 102–121 |
| Chenopodium album | 100–155 | Sorghum halepense | 56–110 |
| Crotolaria spectabilis | 167 | Paspalum pilcatulum | 108 |
| Parthenium hysterophorous | 953 | Cenchrus ciliaris | 104 |
| Sesbania cannabina | 100 | Setaria faberi | 9 |
| Setaria virdis | 19 | ||
| Sorghum halapense | 8 | ||
| Panicum dichotomiflorum | 24 |
| Weed Species | Herbicide | Low Temperature (20/11 °C Day/Night) | High Temperature (29/17 °C Day/Night) | |||
|---|---|---|---|---|---|---|
| ED50 | GR50 | ED50 | GR50 | |||
| Ambrosia artemisiifolia | 2,4-D | 187 | 20 | 61 | 17 | |
| Glyphosate | Susceptible biotype | 437 | 45 | 130 | 39 | |
| Resistant biotype | 2821 | 323 | 1307 | 306 | ||
| A. trifida | 2,4-D | 71 | 15 | 13 | 25 | |
| Glyphosate | Susceptible biotype | 119 | 59 | 62 | 49 | |
| Resistant biotype | 1429 | 349 | 1164 | 218 | ||
| Low Temperature (17.5/7.5 °C Day/Night) | High Temperature (32.5/22.5 °C Day/Night) | |||||
| ED50 | GR50 | ED50 | GR50 | |||
| Kochia scorpia | Glyphosate | Pratt county population | 39 | 34 | 173 | 171 |
| Riley county population | 36 | 46 | 186 | 187 | ||
| Dicamba | Pratt county population | 52 | 21 | 106 | 73 | |
| Riley county population | 105 | 46 | 410 | 225 | ||
| Low Temperature (18 °C) | High Temperature (32 °C) | |||||
| GR50 | GR50 | |||||
| Amaranthus tuberculatus | Mesotrione | 3.6 | 22.6 | |||
| Digitaria sanguinalis | Mesotrione | 11.1 | 82.1 | |||
| Adjuvant | Source of Stress | ||
|---|---|---|---|
| None | Drought | Flooding | |
| None | 84 | 46 | 50 |
| Ammonium sulphate | 90 | 58 | 60 |
| Ammonium nitrate | 85 | 50 | 48 |
| Non-ionic surfactant | 89 | 60 | 62 |
| Methylated seed oil | 84 | 45 | 50 |
| Petroleum oil concentrate | 83 | 50 | 53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, V.; Kumari, A.; Price, A.J.; Bana, R.S.; Singh, V.; Bamboriya, S.D. Impact of Futuristic Climate Variables on Weed Biology and Herbicidal Efficacy: A Review. Agronomy 2023, 13, 559. https://doi.org/10.3390/agronomy13020559
Kumar V, Kumari A, Price AJ, Bana RS, Singh V, Bamboriya SD. Impact of Futuristic Climate Variables on Weed Biology and Herbicidal Efficacy: A Review. Agronomy. 2023; 13(2):559. https://doi.org/10.3390/agronomy13020559
Chicago/Turabian StyleKumar, Vipin, Annu Kumari, Andrew J. Price, Ram Swaroop Bana, Vijay Singh, and Shanti Devi Bamboriya. 2023. "Impact of Futuristic Climate Variables on Weed Biology and Herbicidal Efficacy: A Review" Agronomy 13, no. 2: 559. https://doi.org/10.3390/agronomy13020559