
Drip Irrigation at a Soil Water Suction of 30 kPa Helps AMF and GRSP to Enhance Greenhouse Macro-Aggregates
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenhouse Description and Experimental Design
2.2. Crop Management
2.3. Soil Sampling and Aggregate Fractionation
2.4. Analysis of SOC, TN and GRSP
2.5. High-Throughput Sequencing
2.6. Data Analysis
2.6.1. Bioinformatic Analysis
2.6.2. Statistical Analysis
3. Results
3.1. Aggregate Size Class Distribution and Mean Weight Diameter
3.2. Distribution of SOC, TN and C/N Ratio in the Soil Aggregates
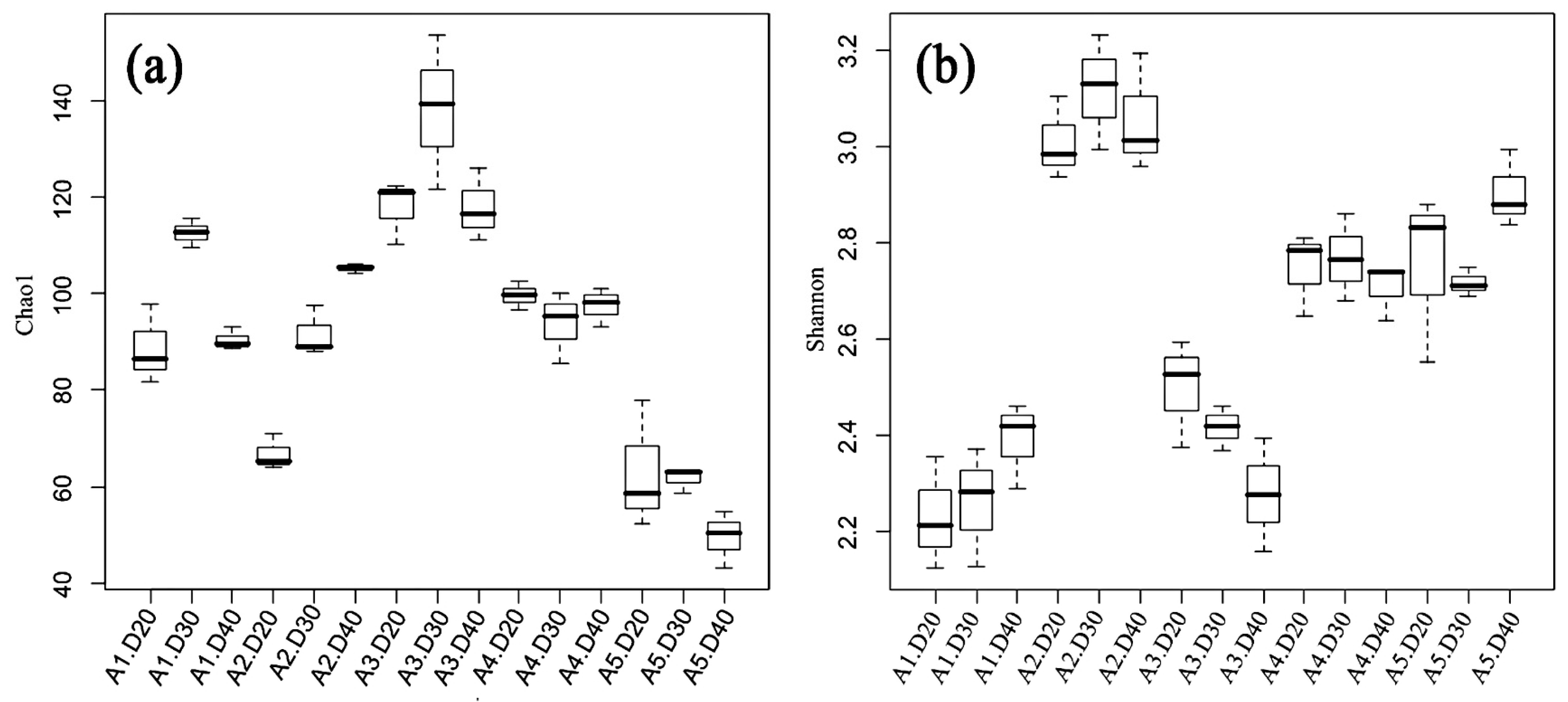
3.3. Diversity and Richness Indices of AMF
3.4. AMF Community Structure
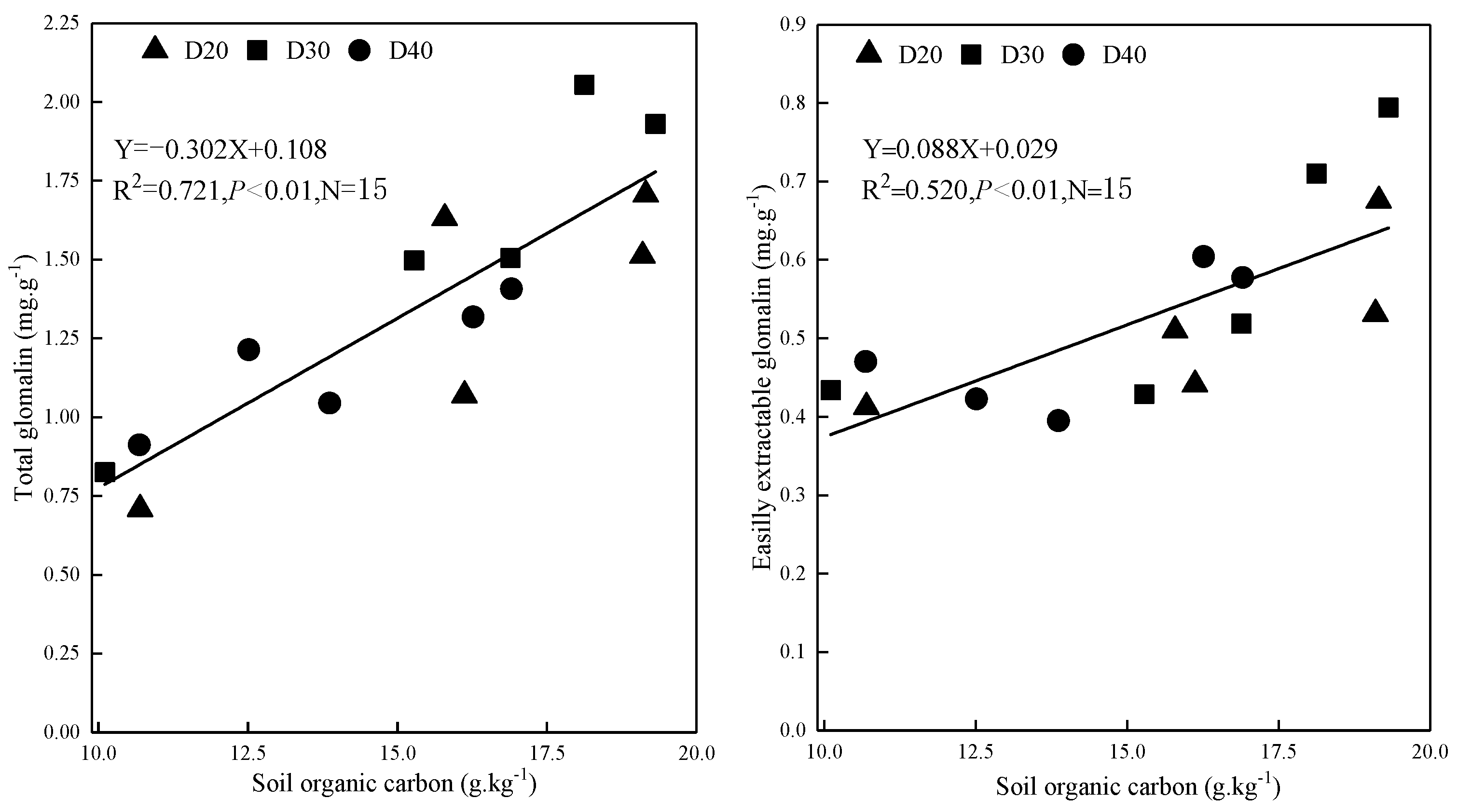
3.5. Variation of Total Glomalin and Easily Extractable Glomalin Levels
3.6. Relationships between GRSP Concentration, AMF OTU Relative Abundance and Soil Properties
4. Discussion
4.1. Distribution of Aggregate Size Classes
4.2. SOC and TN in Different Aggregate Size Classes
4.3. AMF community Diversity and Composition
4.4. Concentration of GRSP
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Camp, C.R. Subsurface drip irrigation: A review. Trans. Asae 1998, 41, 1353–1367. [Google Scholar] [CrossRef]
- Musa, M.; Iqbal, M.; Tariq, M.; Sahi, F.H.; Cheema, N.M.; Jahan, F.N. Comparative water use efficiency of drip and furrow irrigation systems for off-season vegetables under plastic tunnel. SAARC J. Agric. 2014, 12, 62–71. [Google Scholar] [CrossRef]
- Ślusarski, C.; Spotti, C.A. Efficacy of chloropicrin application by drip irrigation in controlling the soil-borne diseases of greenhouse pepper on commercial farms in Poland. Crop Prot. 2016, 89, 216–222. [Google Scholar] [CrossRef]
- Hu, H.; Tian, F.; Hu, H. Soil particle size distribution and its relationship with soil water and salt under mulched drip irrigation in Xinjiang of China. Sci. China Technol. Sci. 2011, 54, 1568. [Google Scholar] [CrossRef]
- Liu, X.B.; Zhang, X.Y.; Wang, Y.X.; Sui, Y.Y.; Zhang, S.L.; Herbert, S.J.; Ding, G. Soil degradation: A problem threatening the sustainable development of agriculture in Northeast China. Plant Soil Environ. 2010, 56, 87–97. [Google Scholar] [CrossRef]
- Tisdall, J.M.; Oades, J.M. Organic matter and water-stable aggregates in soils. Eur. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Bronick, C.J.; Lal, R. Soil structure and management: A review. Geoderma 2005, 124, 3–22. [Google Scholar] [CrossRef]
- Matthiasc, R.; Noorf, M.; Evaf, L.; Pedrom, A. Mycelium of arbuscular mycorrhizal fungi increases soil water repellency and is sufficient to maintain water-stable soil aggregates. Soil Biol. Biochem. 2010, 42, 1189–1191. [Google Scholar]
- Gehring, C.A. Chapter 13—Introduction: Mycorrhizas and soil structure, moisture, and salinity. Fertility, Structure, and Carbon Storage. 2017, 235–240, 235–240. [Google Scholar]
- Wright, S.F.; Upadhyaya, A. A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi. Plant Soil 1998, 198, 97–107. [Google Scholar] [CrossRef]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular Mycorrhizal Fungi as Natural Biofertilizers: Let’s Benefit from Past Successes. Front Microbiol. 2016, 6, 1559. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.; White, P.J.; Li, C. Colonization and community structure of arbuscular mycorrhizal fungi in maize roots at different depths in the soil profile respond differently to phosphorus inputs on a long-term experimental site. Mycorrhiza 2017, 27, 369. [Google Scholar]
- Zhu, Y.G.; Miller, R.M. Carbon cycling by arbuscular mycorrhizal fungi in soil-plant systems. Trends Plant Sci. 2003, 8, 407. [Google Scholar] [CrossRef] [PubMed]
- Kohler, J.; Roldán, A.; Campoy, M.; Caravaca, F. Unraveling the role of hyphal networks from arbuscular mycorrhizal fungi in aggregate stabilization of semiarid soils with different textures and carbonate contents. Plant Soil 2017, 410, 273–281. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Bedini, S.; Pellegrino, E.; Avio, L.; Pellegrini, S.; Bazzoffi, P.; Argese, E.; Giovannetti, M. Changes in soil aggregation and glomalin-related soil protein content as affected by the arbuscular mycorrhizal fungal species Glomus mosseae and Glomus intraradices. Soil Biol. Biochem. 2009, 41, 1491–1496. [Google Scholar] [CrossRef]
- Olsson, P.A.; Johnson, N.C. Tracking carbon from the atmosphere to the rhizosphere. Ecol. Lett. 2005, 8, 1264–1270. [Google Scholar] [CrossRef]
- Fokom, R.; Adamou, S.; Teugwa, M.C.; Boyogueno, A.D.B.; Nana, W.L.; Ngonkeu, M.E.L.; Tchameni, N.S.; Nwaga, D.; Ndzomo, G.T.; Zollo, P.H.A. Glomalin related soil protein, carbon, nitrogen and soil aggregate stability as affected by land use variation in the humid forest zone of south Cameroon. Soil Tillage Res. 2012, 120, 69–75. [Google Scholar] [CrossRef]
- Hodge, A.; Fitter, A.H. Substantial nitrogen acquisition by arbuscular mycorrhizal fungi from organic material has implications for N cycling. Proc. Natl. Acad. Sci. USA 2010, 107, 13754. [Google Scholar] [CrossRef]
- Rillig, M.C. Arbuscular mycorrhizae and terrestrial ecosystem processes. Ecol. Lett. 2004, 7, 740–754. [Google Scholar] [CrossRef]
- Borie, F.; Rubio, R.; Morales, A. Arbuscular mycorrhizal fungi and soil aggregation hongos micorrícicos arbusculares y agregación de suelo. J. Soil Sci. Plant Nutr. 2008, 8, 9–18. [Google Scholar]
- Varelacervero, S.; Vasar, M.; Davison, J.; Barea, J.M. The composition of arbuscular mycorrhizal fungal communities differs among the roots, spores and extraradical mycelia associated with five Mediterranean plant species. Environ. Microbiol. 2015, 17, 2882–2895. [Google Scholar] [CrossRef]
- Hontoria, C.; Velásquez, R.; Benito, M.; Almorox, J.; Moliner, A. Bradford-reactive soil proteins and aggregate stability under abandoned versus tilled olive groves in a semi-arid calcisol. Soil Biol. Biochem. 2009, 41, 1583–1585. [Google Scholar] [CrossRef]
- Gordon, H.; Haygarth, P.M.; Bardgett, R.D. Drying and rewetting effects on soil microbial community composition and nutrient leaching. Soil Biol. Biochem. 2008, 40, 302–311. [Google Scholar] [CrossRef]
- Ye, X.H.; Han, B.; Li, W.; Zhang, X.C.; Zhang, Y.L.; Lin, X.G.; Zou, H.T. Effects of different irrigation methods on nitrous oxide emissions and ammonia oxidizers microorganisms in greenhouse tomato fields. Agric. Water Manag. 2018, 203, 115–123. [Google Scholar] [CrossRef]
- Allison, S.D.; Jastrow, J.D. Activities of extracellular enzymes in physically isolated fractions of restored grassland soils. Soil Biol. Biochem. 2006, 38, 3245–3256. [Google Scholar] [CrossRef]
- Six, J.; Paustian, K.; Elliott, E.T.; Combrink, C. Soil structure and organic matter: I. Distribution of aggregate-size classes and aggregate-associated carbon. Soil Sci. Soc. Am. J. 2000, 64, 681–689. [Google Scholar] [CrossRef]
- Kemper, W.D.; Rosenau, R.C.; Klute, A. Aggregate stability and size distribution. Methods Soil Anal. Part 1 Phys. Mineral. Methods 1986, 5, 425–442. [Google Scholar]
- He, F. Bradford Protein Assay. Bio-Protocol 2011, 1, e45. [Google Scholar] [CrossRef]
- Sato, K.; Suyama, Y.; Saito, M.; Sugawara, K. A new primer for discrimination of arbuscular mycorrhizal fungi with polymerase chain reaction-denature gradient gel electrophoresis. Grassl. Sci. 2005, 51, 179–181. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric Estimation of the Number of Classes in a Population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Ma, R.; Cai, C.; Li, Z.; Wang, J.; Xiao, T.; Peng, G.; Yang, W. Evaluation of soil aggregate microstructure and stability under wetting and drying cycles in two Ultisols using synchrotron-based X-ray micro-computed tomography. Soil Tillage Res. 2015, 149, 1–11. [Google Scholar] [CrossRef]
- Truman, C.C.; Bradford, J.M.; Ferris, J.E. Antecedent Water Content and Rainfall Energy Influence on Soil Aggregate Breakdown. Soil Sci. Soc. Am. J. 1990, 54, 1385–1392. [Google Scholar] [CrossRef]
- Denef, K.; Six, J.; Paustian, K.; Merckx, R. Importance of macroaggregate dynamics in controlling soil carbon stabilization: Short-term effects of physical disturbance induced by dry–wet cycles. Soil Biol. Biochem. 2001, 33, 2145–2153. [Google Scholar] [CrossRef]
- Lado, M.; Ben-Hur, M.; Shainberg, I. Soil Wetting and Texture Effects on Aggregate Stability, Seal Formation, and Erosion. Soil Sci. Soc. Am. J. 2004, 68, 1992–1999. [Google Scholar] [CrossRef]
- Wang, E.; Cruse, R.M.; Chen, X.; Daigh, A. Effects of moisture condition and freeze/thaw cycles on surface soil aggregate size distribution and stability. Can. J. Soil Sci. 2012, 92, 529–536. [Google Scholar] [CrossRef]
- Chen, X.; Li, Z.; Liu, M.; Jiang, C.; Che, Y. Microbial community and functional diversity associated with different aggregate fractions of a paddy soil fertilized with organic manure and/or NPK fertilizer for 20 years. J. Soils Sediments 2015, 15, 292–301. [Google Scholar] [CrossRef]
- Qiu, L.; Wei, X.; Gao, J.; Zhang, X. Dynamics of soil aggregate-associated organic carbon along an afforestation chronosequence. Plant Soil 2015, 391, 237–251. [Google Scholar] [CrossRef]
- Sainju, U.M.; Caesar-TonThat, T.; Jabro, J.D. Carbon and nitrogen fractions in dryland soil aggregates affected by long-term tillage and cropping sequence. Soil Sci. Soc. Am. J. 2009, 73, 1488–1495. [Google Scholar] [CrossRef]
- McCarthy, J.F.; Ilavsky, J.; Jastrow, J.D.; Mayer, L.M.; Perfect, E.; Zhuang, J. Protection of organic carbon in soil microaggregates via restructuring of aggregate porosity and filling of pores with accumulating organic matter. Geochim. Et Cosmochim. Acta 2008, 72, 4725–4744. [Google Scholar] [CrossRef]
- Zinn, Y.L.; Lal, R.; Resck, D.V.S. Texture and organic carbon relations described by a profile pedotransfer function for Brazilian Cerrado soils. Geoderma 2005, 127, 168–173. [Google Scholar] [CrossRef]
- Li, D.; Shao, M.A. Soil organic carbon and influencing factors in different landscapes in an arid region of northwestern China. Catena 2014, 116, 95–104. [Google Scholar] [CrossRef]
- Turner, B.L.; Driessen, J.P.; Haygarth, P.M.; Mckelvie, I.D. Potential contribution of lysed bacterial cells to phosphorus solubilisation in two rewetted Australian pasture soils. Soil Biol. Biochem. 2003, 35, 187–189. [Google Scholar] [CrossRef]
- Mikha, M.M.; Rice, C.W.; Milliken, G.A. Carbon and nitrogen mineralization as affected by drying and wetting cycles. Soil Biol. Biochem. 2005, 37, 339–347. [Google Scholar] [CrossRef]
- Mcinerney, M.; Bolger, T. Temperature, wetting cycles and soil texture effects on carbon and nitrogen dynamics in stabilized earthworm casts. Soil Biol. Biochem. 2000, 32, 335–349. [Google Scholar] [CrossRef]
- Yang, H.; Koide, R.T.; Zhang, Q. Short-term waterlogging increases arbuscular mycorrhizal fungal species richness and shifts community composition. Plant Soil 2016, 404, 373–384. [Google Scholar] [CrossRef]
- Stevens, K.J.; Spender, S.W.; Peterson, R.L. Phosphorus, arbuscular mycorrhizal fungi and performance of the wetland plant Lythrum salicaria L. under inundated conditions. Mycorrhiza 2002, 12, 277. [Google Scholar] [PubMed]
- Yıldırım, O.; Halloran, N.; Çavuşoğlu, S.; ŞEngül, N. Effects of different irrigation programs on the growth, yield, and fruit quality of drip-irrigated melon. Turk. J. Agric. For. 2014, 33, 243–255. [Google Scholar]
- Miller, S.P.; Sharitz, R.R. Manipulation of Flooding and Arbuscular Mycorrhiza Formation Influences Growth and Nutrition of Two Semiaquatic Grass Species. Funct. Ecol. 2010, 14, 738–748. [Google Scholar] [CrossRef]
- Mendoza, R.; Escudero, V.; García, I. Plant growth, nutrient acquisition and mycorrhizal symbioses of a waterlogging tolerant legume (Lotus glaber Mill.) in a saline-sodic soil. Plant Soil 2005, 275, 305–315. [Google Scholar] [CrossRef]
- Braunberger, P.G.; Abbott, L.K.; Robson, A.D. Infectivity of arbuscular mycorrhizal fungi after wetting and drying. New Phytol. 1996, 134, 673–684. [Google Scholar] [CrossRef]
- Dai, J.; Hu, J.; Zhu, A.; Bai, J.; Wang, J.; Lin, X. No tillage enhances arbuscular mycorrhizal fungal population, glomalin-related soil protein content, and organic carbon accumulation in soil macroaggregates. J. Soils Sediments 2015, 15, 1055–1062. [Google Scholar] [CrossRef]
- Daniell, T.J.; Husband, R.; Fitter, A.H.; Young, J.P.W. Molecular diversity of arbuscular mycorrhizal fungi colonising arable crops. Fems Microbiol. Ecol. 2001, 36, 203. [Google Scholar] [CrossRef]
- Ladd, J.N.; Gestel, M.V.; Monrozier, L.J.; Amato, M. Distribution of organic 14 C and 15 N in particle-size fractions of soils incubated with 14 C, 15 N-labelled glucose/NH 4, and legume and wheat straw residues. Soil Biol. Biochem. 1996, 28, 893–905. [Google Scholar] [CrossRef]
- Hammer, E.C.; Rillig, M.C. The Influence of Different Stresses on Glomalin Levels in an Arbuscular Mycorrhizal Fungus—Salinity Increases Glomalin Content. PLoS ONE 2011, 6, e28426. [Google Scholar] [CrossRef] [PubMed]
- Josef, K.; Fuensanta, C.; Antonio, R. Effect of drought on the stability of rhizosphere soil aggregates of Lactuca sativa grown in a degraded soil inoculated with PGPR and AM fungi. Appl. Soil Ecol. 2009, 42, 160–165. [Google Scholar]
- Lovelock, C.E.; Wright, S.F.; Nichols, K.A. Using glomalin as an indicator for arbuscular mycorrhizal hyphal growth: An example from a tropical rain forest soil. Soil Biol. Biochem. 2004, 36, 1009–1012. [Google Scholar] [CrossRef]
- Driver, J.D.; Holben, W.E.; Rillig, M.C. Characterization of glomalin as a hyphal wall component of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2005, 37, 101–106. [Google Scholar] [CrossRef]
- Saidi, A.; Husin, E.F.; Rasyidin, A.; Eddiwal, E.; Ismon, L. Selection of Aruscular Mycorrhizal Fungi (AMF) indigeneus in ultisol for promoting the production of glmalin and aggregate formation processes. Int. J. Adv. Sci. Eng. Inf. Technol. 2015, 4, 430. [Google Scholar] [CrossRef]
- Wu, Q.S.; Cao, M.Q.; Zou, Y.N.; He, X. Direct and indirect effects of glomalin, mycorrhizal hyphae, and roots on aggregate stability in rhizosphere of trifoliate orange. Sci. Rep. 2014, 4, 5823. [Google Scholar] [CrossRef]
- Rillig, M.C.; Maestre, F.T.; Lamit, L.J. Microsite differences in fungal hyphal length, glomalin, and soil aggregate stability in semiarid Mediterranean steppes. Soil Biol. Biochem. 2003, 35, 1257–1260. [Google Scholar] [CrossRef]
- Yang, Y.; He, C.; Li, H.; Ban, Y.; Ming, T. The effects of arbuscular mycorrhizal fungi on glomalin-related soil protein distribution, aggregate stability and their relationships with soil properties at different soil depths in lead-zinc contaminated area. PLoS ONE 2017, 12, e0182264. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.S.; Huang, Y.M.; Yan, L.; Nasrullah; He, X.H. Contribution of arbuscular mycorrhizas to glomalin-related soil protein, soil organic carbon and aggregate stability in citrus rhizosphere. Int. J. Agric. Biol. 2014, 16, 207–212. [Google Scholar]
- Rillig, M.C.; Ramsey, P.W.; Morris, S.; Paul, E.A. Glomalin, an arbuscular-mycorrhizal fungal soil protein, responds to land-use change. Plant Soil 2003, 253, 293–299. [Google Scholar] [CrossRef]
- Peng, S.; Guo, T.; Liu, G. The effects of arbuscular mycorrhizal hyphal networks on soil aggregations of purple soil in southwest China. Soil Biol. Biochem. 2013, 57, 411–417. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aggregate Size Class (cm) | Treatment | SOC (g kg−1) | TN (g kg−1) | C/N Ratio |
|---|---|---|---|---|
| >2 | D20 | 15.79 ± 0.21 a | 1.48 ± 0.02 a | 10.73 ± 0.06 b |
| D30 | 16.89 ± 0.56 a | 1.33 ± 0.05 a | 12.92 ± 0.07 a | |
| D40 | 12.51 ± 1.26 b | 1.24 ± 0.09 b | 10.07 ± 0.38 b | |
| 1–2 | D20 | 19.15 ± 1.42 a | 1.87 ± 0.06 a | 11.02 ± 0.53 a |
| D30 | 19.31 ± 1.92 a | 1.93 ± 0.10 a | 10.08 ± 0.47 a | |
| D40 | 16.91 ± 0.80 b | 1.62 ± 0.03 b | 10.44 ± 1.06 a | |
| 0.25–1 | D20 | 19.10 ± 0.39 a | 1.76 ± 0.01 a | 11.06 ± 0.28 a |
| D30 | 18.13 ± 0.46 b | 1.73 ± 0.01 a | 10.45 ± 0.20 a | |
| D40 | 16.26 ± 0.35 c | 1.68 ± 0.03 b | 10.03 ± 0.16 a | |
| 0.053–0.25 | D20 | 16.12 ± 0.43 a | 1.47 ± 0.11 a | 10.32 ± 0.17 a |
| D30 | 15.28 ± 0.40 a | 1.48 ± 0.14 a | 10.74 ± 0.72 a | |
| D40 | 13.87 ± 0.84 b | 1.31 ± 0. 06 b | 10.16 ± 0.41 a | |
| <0.053 | D20 | 10.70 ± 0.37 a | 1.12 ± 0.07 a | 9.87 ± 0.08 a |
| D30 | 10.11 ± 0.50 a | 1.08 ± 0.04 a | 9.29 ± 0.20 a | |
| D40 | 10.69 ± 0.34 a | 1.14 ± 0.02 a | 9.34 ± 0.16 a | |
| A | *** | *** | * | |
| D | *** | ** | NS | |
| A × D | ** | NS | NS | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, X.; Li, J.; Ma, J.; Fan, Q.; Yu, N.; Zhang, Y.; Zou, H.; Zhang, Y. Drip Irrigation at a Soil Water Suction of 30 kPa Helps AMF and GRSP to Enhance Greenhouse Macro-Aggregates. Agronomy 2023, 13, 522. https://doi.org/10.3390/agronomy13020522
Ye X, Li J, Ma J, Fan Q, Yu N, Zhang Y, Zou H, Zhang Y. Drip Irrigation at a Soil Water Suction of 30 kPa Helps AMF and GRSP to Enhance Greenhouse Macro-Aggregates. Agronomy. 2023; 13(2):522. https://doi.org/10.3390/agronomy13020522
Chicago/Turabian StyleYe, Xuhong, Jiaqi Li, Jianhui Ma, Qingfeng Fan, Na Yu, Yuling Zhang, Hongtao Zou, and Yulong Zhang. 2023. "Drip Irrigation at a Soil Water Suction of 30 kPa Helps AMF and GRSP to Enhance Greenhouse Macro-Aggregates" Agronomy 13, no. 2: 522. https://doi.org/10.3390/agronomy13020522