
Response of Organic Fertilizer Application to Soil Microorganisms and Forage Biomass in Grass–Legume Mixtures
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling
2.2. DNA Extraction, Amplification, Sequencing, and Sequence Analysis
2.3. Sequence Data Preprocessing and Bioinformatics Approaches
2.4. Statistical Analysis
2.5. Source Tracker Analysis
2.6. Stochastic Matrix Theory Based Molecular Ecology Networks and Analysis
3. Results
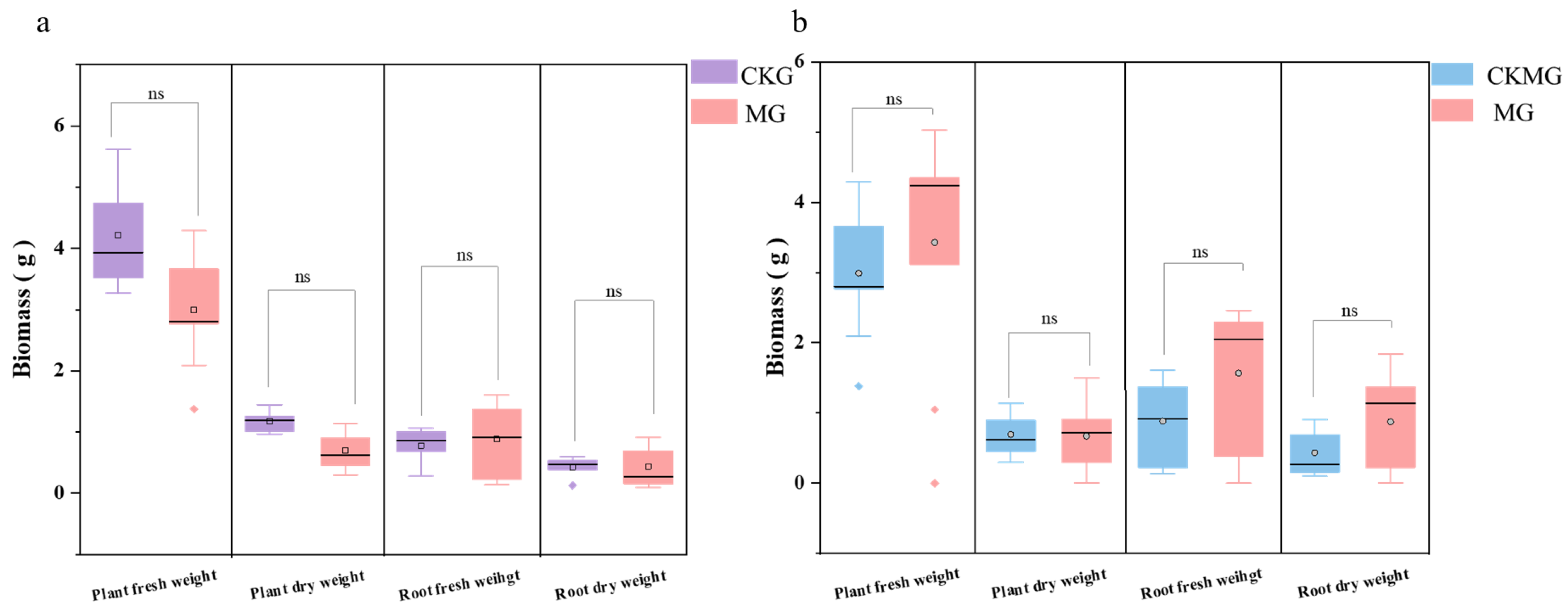
3.1. Effects of Organic Fertilization on Physical and Chemical Properties of Soil, and Forage Biomass of Grass–Legume Mixtures
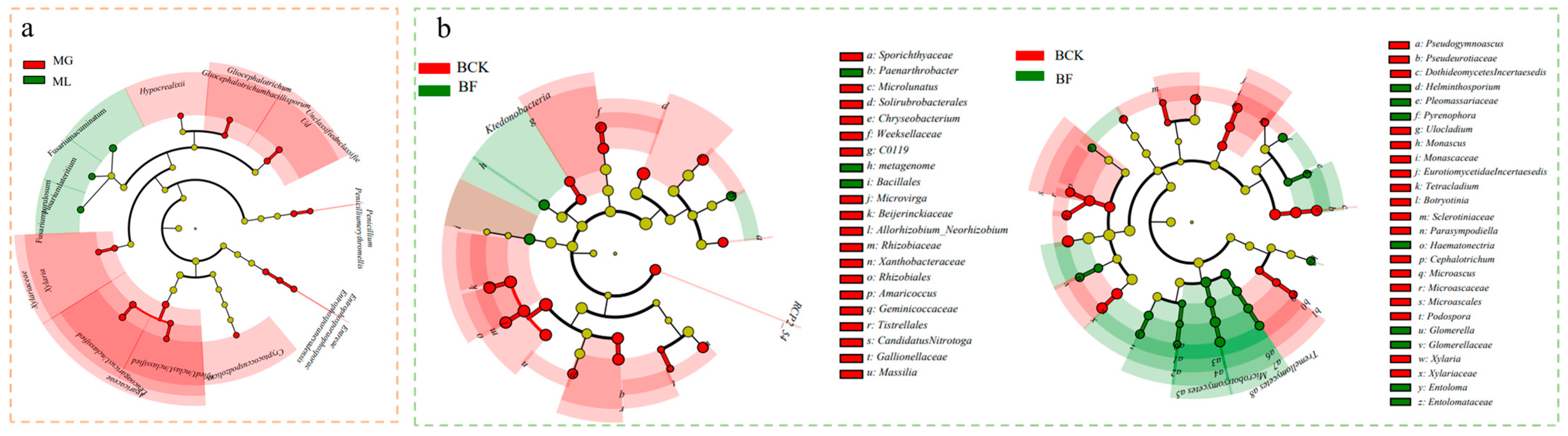
3.2. Effect of Organic Fertilization on Microbial Community Diversity, Structure, and Composition
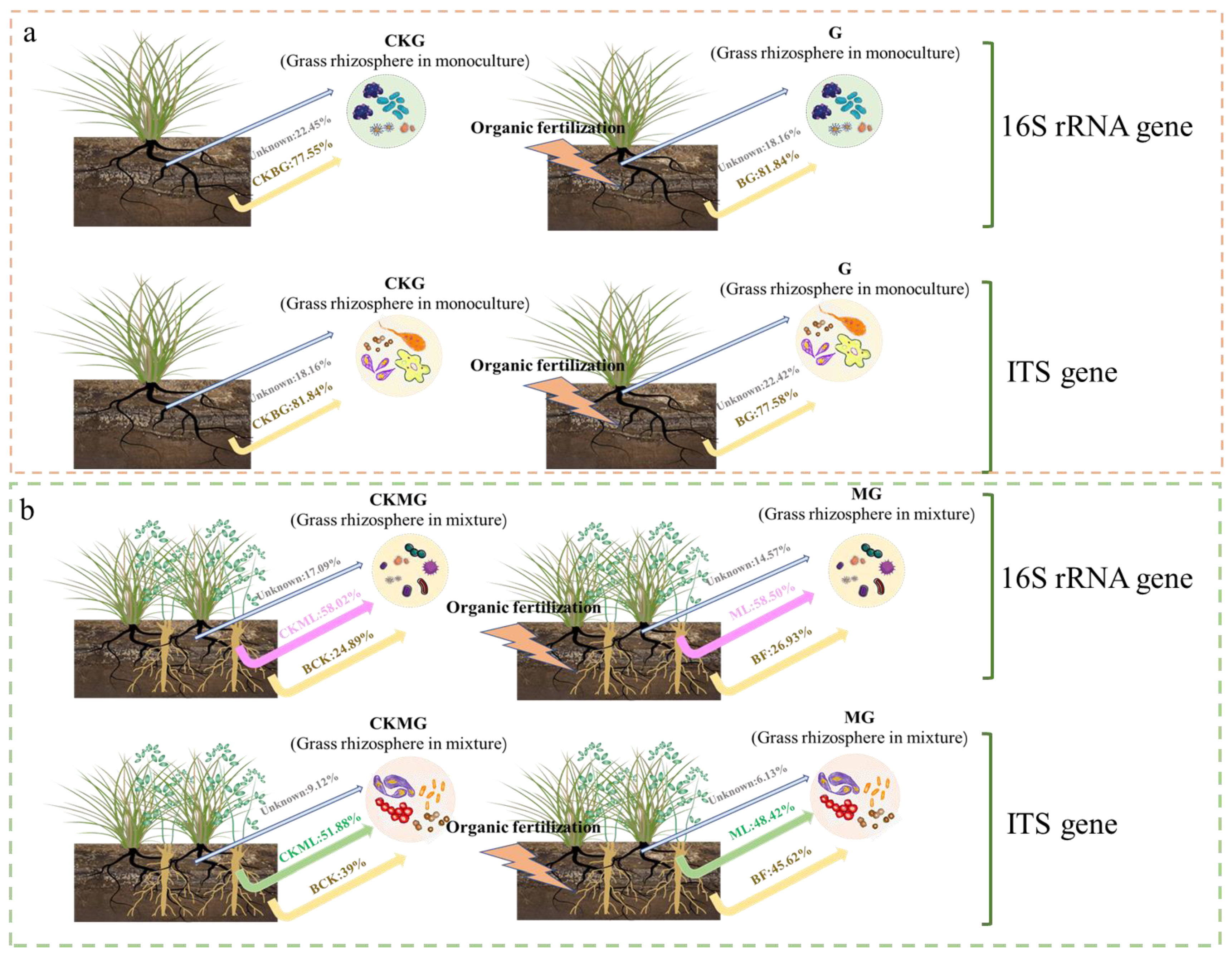
3.3. Source Tracker Analyzes the Main Source of Rhizosphere Prokaryote and Fungal Communities after Application of Organic Fertilizer
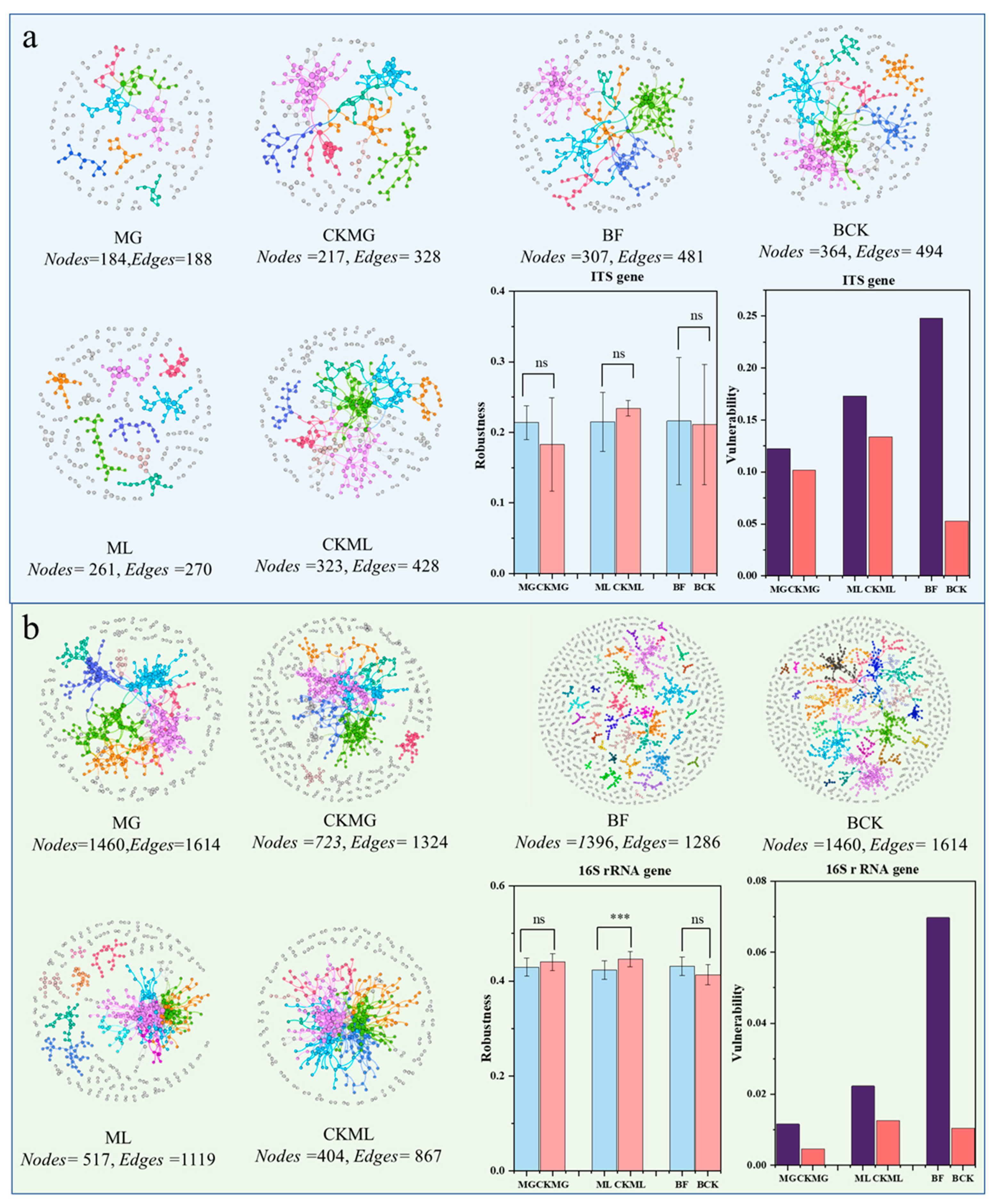
3.4. Molecular Ecological Network Analysis on Soil Communities
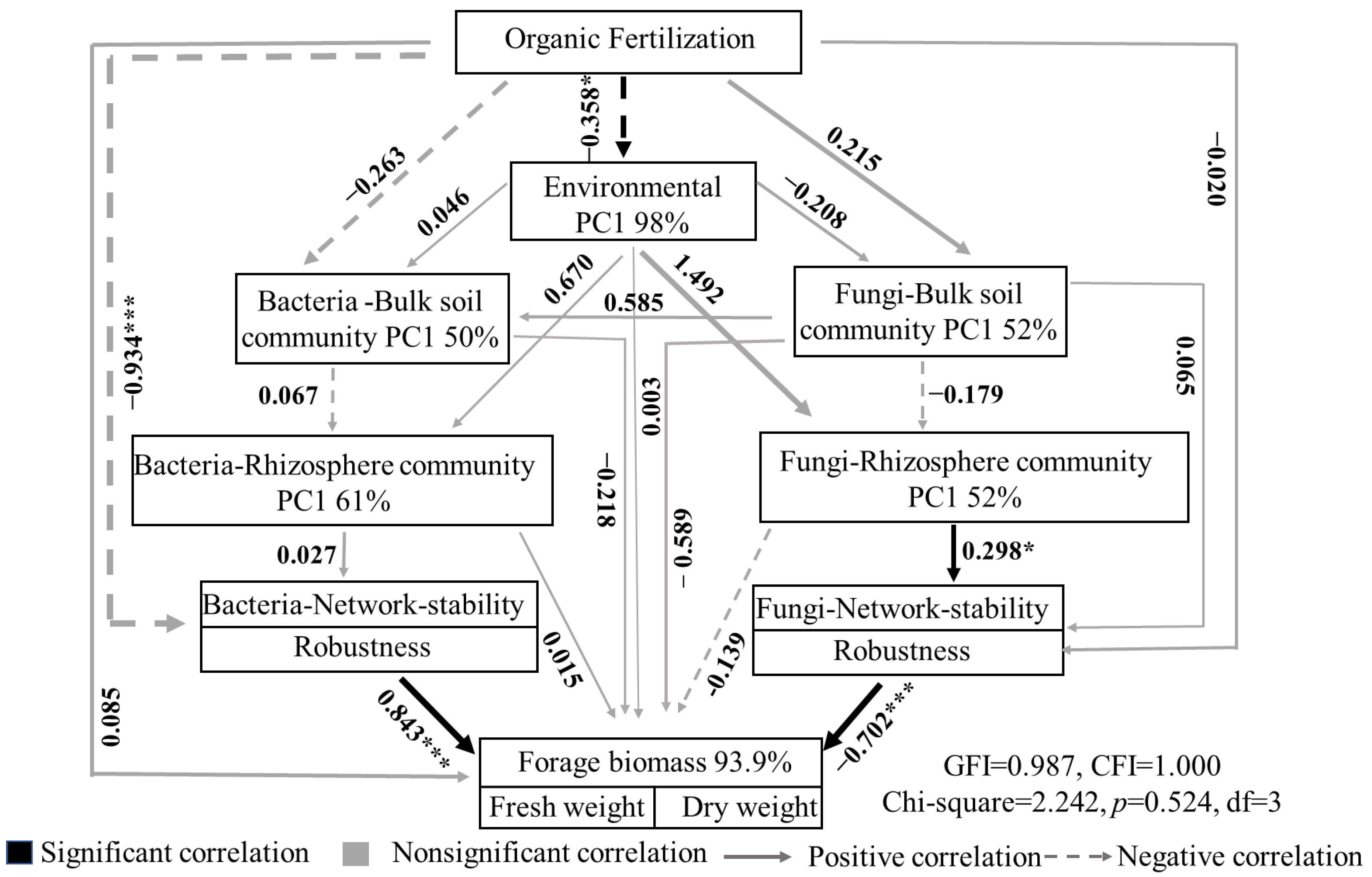
3.5. Structural Equation Model Analysis of Relationship between Soil Microbial Community and Forage Biomass under Organic Fertilizer Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gu, C.; Liang, Y.; Li, J.; Shao, H.; Jiang, Y.; Zhou, X.; Gao, C.; Li, X.; Zhang, W.; Guo, C.; et al. Saline lakes on the Qinghai-Tibet Plateau harbor unique viral assemblages mediating microbial environmental adaption. iScience 2021, 24, 103439. [Google Scholar] [CrossRef]
- Ma, K.; Wang, Y.; Jin, X.; Zhao, Y.; Yan, H.; Zhang, H.; Zhou, X.; Lu, G.; Deng, Y. Application of Organic Fertilizer Changes the Rhizosphere Microbia Communities of a Gramineous Grass on Qinghai Tibet Plateau. Microorganisms 2022, 10, 1148. [Google Scholar] [CrossRef]
- Zhang, H.; Yao, Z.; Wang, K.; Zheng, X.; Ma, L.; Wang, R.; Liu, C.; Zhang, W.; Zhu, B.; Tang, X.; et al. Annual N2O emissions from conventionally grazed typical alpine grass meadows in the eastern Qinghai–Tibetan Plateau. Sci. Total. Environ. 2018, 625, 885–899. [Google Scholar] [CrossRef]
- Lang, M.; Li, P.; Long, G.; Yuan, F.; Yu, Y.; Ma, E.; Shan, J.; Müller, C.; Zhu, T. Grazing rest versus no grazing stimulates soil inorganic N turnover in the alpine grasslands of the Qinghai-Tibet plateau. Catena 2021, 204, 105382. [Google Scholar] [CrossRef]
- Yan, H.; Gu, S.; Li, S.; Shen, W.; Zhou, X.; Yu, H.; Ma, K.; Zhao, Y.; Wang, Y.; Zheng, H.; et al. Grass-legume mixtures enhance forage production via the bacterial community. Agric. Ecosyst. Environ. 2022, 338, 108087. [Google Scholar] [CrossRef]
- Bao, G.; Suetsugu, K.; Wang, H.; Yao, X.; Liu, L.; Ou, J.; Li, C. Effects of the hemiparasitic plant Pedicularis kansuensis on plant community structure in a degraded grassland. Ecol. Res. 2015, 30, 507–515. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, X.; Zhang, L.; Zeng, L.; Liu, Y.; Wang, X.; He, P.; Li, S.; Liang, G.; Zhou, W.; et al. The stronger impact of inorganic nitrogen fertilization on soil bacterial community than organic fertilization in short-term condition. Geoderma 2021, 382, 114752. [Google Scholar] [CrossRef]
- Suleiman, A.K.; Harkes, P.; Elsen, S.V.D.; Holterman, M.; Korthals, G.W.; Helder, J.; Kuramae, E.E. Organic amendment strengthens interkingdom associations in the soil and rhizosphere of barley (Hordeum vulgare). Sci. Total. Environ. 2019, 695, 133885. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Li, J.; Wang, Q.; Guo, M.; Yan, D.; Fang, W.; Ren, Z.; Wang, Q.; Ouyang, C.; Li, Y.; et al. Effect of soil fumigants on degradation of abamectin and their combination synergistic effect to root-knot nematode. PLoS ONE 2018, 13, e0188245. [Google Scholar] [CrossRef]
- Wu, Y. Periphyton: A Promising Bio-Organic Fertilizer Source in Agricultural Ecosystems; Elsevier: Boston, MA, USA, 2017; pp. 225–249. [Google Scholar]
- Wołejko, E.; Jabłońska-Trypuć, A.; Wydro, U.; Butarewicz, A.; Łozowicka, B. Soil biological activity as an indicator of soil pollution with pesticides—A review. Appl. Soil Ecol. 2020, 147, 103356. [Google Scholar] [CrossRef]
- Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Bei, S.; Zhang, Y.; Li, T.; Christie, P.; Li, X.; Zhang, J. Response of the soil microbial community to different fertilizer inputs in a wheat-maize rotation on a calcareous soil. Agric. Ecosyst. Environ. 2018, 260, 58–69. [Google Scholar] [CrossRef]
- Yang, Y.; Li, X.; Liu, J.; Zhou, Z.; Zhang, T.; Wang, X. Fungal community structure in relation to manure rate in red soil in southern China. Appl. Soil Ecol. 2020, 147, 103442. [Google Scholar] [CrossRef]
- Ling, N.; Deng, K.; Song, Y.; Wu, Y.; Zhao, J.; Raza, W.; Huang, Q.; Shen, Q. Variation of rhizosphere bacterial community in watermelon continuous mono-cropping soil by long-term application of a novel bioorganic fertilizer. Microbiol. Res. 2014, 169, 570–578. [Google Scholar] [CrossRef]
- Ji, L.; Si, H.; He, J.; Fan, L.; Li, L. The shifts of maize soil microbial community and networks are related to soil properties under different organic fertilizers. Rhizosphere 2021, 19, 100388. [Google Scholar] [CrossRef]
- Deng, Y.; Jiang, Y.-H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Thébault, E.; Fontaine, C. Stability of Ecological Communities and the Architecture of Mutualistic and Trophic Networks. Science 2010, 329, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Yang, T.; Friman, V.-P.; Xu, Y.; Shen, Q.; Jousset, A. Trophic network architecture of root-associated bacterial communities determines pathogen invasion and plant health. Nat. Commun. 2015, 6, 8413. [Google Scholar] [CrossRef]
- Li, C.; Wang, L.; Ji, S.; Chang, M.; Wang, L.; Gan, Y.; Liu, J. The ecology of the plastisphere: Microbial composition, function, assembly, and network in the freshwater and seawater ecosystems. Water Res. 2021, 202, 117428. [Google Scholar] [CrossRef]
- Taylor, D.L.; Walters, W.A.; Lennon, N.J.; Bochicchio, J.; Krohn, A.; Caporaso, J.G.; Pennanen, T. Accurate Estimation of Fungal Diversity and Abundance through Improved Lineage-Specific Primers Optimized for Illumina Amplicon Sequencing. Appl. Environ. Microbiol. 2016, 82, 7217–7226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, K.; Zhang, Z.; Cai, W.; Liu, W.; Xu, M.; Yin, H.; Wang, A.; He, Z.; Deng, Y. Biodiversity and species competition regulate the resilience of microbial biofilm community. Mol. Ecol. 2017, 26, 6170–6182. [Google Scholar] [CrossRef]
- Kong, Y. Btrim: A fast, lightweight adapter and quality trimming program for next-generation sequencing technologies. Genomics 2011, 98, 152–153. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Deng, Y.; Lian, S.; Dai, C.; Ma, Q.; Qu, Y. Succession of diversity, functions, and interactions of the fungal community in activated sludge under aromatic hydrocarbon stress. Environ. Res. 2022, 204, 112143. [Google Scholar] [CrossRef]
- Edgar, R.C. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. bioRxiv 2016, 15, 081257. [Google Scholar]
- Lin, T.J. Exploring the Differences in Taiwanese University Students’ Online Learning Task Value, Goal Orientation, and Self-Efficacy Before and After the COVID-19 Outbreak. Asia-Pacific Edu. Res. 2021, 30, 191–203. [Google Scholar] [CrossRef]
- Satorra, A.; Bentler, P.M. Scaling Corrections for Statistics in Covariance Structure Analysis; UCLA: Los Angeles, CA, USA, 1988. [Google Scholar]
- Knights, D.; Kuczynski, J.; Charlson, E.S.; Zaneveld, J.; Mozer, M.C.; Collman, R.G.; Bushman, F.D.; Knight, R.T.; Kelley, S.T. Bayesian community-wide culture-independent microbial source tracking. Nat. Methods 2011, 8, 761–763. [Google Scholar] [CrossRef]
- Feng, K.; Peng, X.; Zhang, Z.; Gu, S.; He, Q.; Shen, W.; Wang, Z.; Wang, D.; Hu, Q.; Li, Y.; et al. iNAP: An integrated network analysis pipeline for microbiome studies. iMeta 2022, 1, e13. [Google Scholar] [CrossRef]
- Montesinos-Navarro, A.; Hiraldo, F.; Tella, J.L.; Blanco, G. Network structure embracing mutualism–antagonism continuums increases community robustness. Nat. Ecol. Evol. 2017, 1, 1661–1669. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, P.; Qin, Y.; Tu, Q.; Yang, Y.; He, Z.; Schadt, C.W.; Zhou, J. Network succession reveals the importance of competition in response to emulsified vegetable oil amendment for uranium bioremediation. Environ. Microbiol. 2016, 18, 205–218. [Google Scholar] [CrossRef]
- Tao, C.; Li, R.; Xiong, W.; Shen, Z.; Liu, S.; Wang, B.; Ruan, Y.; Geisen, S.; Shen, Q.; Kowalchuk, G.A. Bio-organic fertilizers stimulate indigenous soil Pseudomonas populations to enhance plant disease suppression. Microbiome 2020, 8, 137. [Google Scholar] [CrossRef]
- Wang, J.; Song, Y.; Ma, T.; Raza, W.; Li, J.; Howland, J.G.; Huang, Q.; Shen, Q. Impacts of inorganic and organic fertilization treatments on bacterial and fungal communities in a paddy soil. Appl. Soil Ecol. 2017, 112, 42–50. [Google Scholar] [CrossRef]
- Li, W.; Zhang, F.; Cui, G.; Wang, Y.; Yang, J.; Cheng, H.; Liu, H.; Zhang, L. Effects of bio-organic fertilizer on soil fertility, microbial community composition, and potato growth. Scienceasia 2021, 47, 347. [Google Scholar] [CrossRef]
- Liu, Z.-L.; Gao, J.-J.; Gu, D.-Y.; Yan, W.-Q. [Effects of organic fertilizer on soil environment and yield of tomato under year-round cultivation]. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2020, 31, 929–934. [Google Scholar]
- Liang, J.-P.; Xue, Z.-Q.; Yang, Z.-Y.; Chai, Z.; Niu, J.-P.; Shi, Z.-Y. Effects of microbial organic fertilizers on Astragalus membranaceus growth and rhizosphere microbial community. Ann. Microbiol. 2021, 71, 11. [Google Scholar] [CrossRef]
- Li, D.; Davis, J.E.; Sun, Y.; Wang, G.; Nabi, G.; Wingfield, J.C.; Lei, F. Coping with extremes: Convergences of habitat use, territoriality, and diet in summer but divergences in winter between two sympatric snow finches on the Qinghai-Tibet Plateau. Integr. Zool. 2020, 15, 533–543. [Google Scholar] [CrossRef]
- Oberson, A.; Frossard, E.; Bühlmann, C.; Mayer, J.; Mäder, P.; Lüscher, A. Nitrogen fixation and transfer in grass-clover leys under organic and conventional cropping systems. Plant Soil 2013, 371, 237–255. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, G.; Yu, H.; Du, X.; He, Q.; Yao, S.; Zhao, L.; Huang, C.; Wen, X.; Deng, Y. Meadow degradation increases spatial turnover rates of the fungal community through both niche selection and dispersal limitation. Sci. Total. Environ. 2021, 798, 149362. [Google Scholar] [CrossRef]
- Cai, Y.; Shen, J.-P.; Di, H.J.; Zhang, L.-M.; Zhang, C.; He, J.-Z. Variation of soil nitrate and bacterial diversity along soil profiles in manure disposal maize field and adjacent woodland. J. Soils Sediments 2020, 20, 3557–3568. [Google Scholar] [CrossRef]
- Zhong, W.; Gu, T.; Wang, W.; Zhang, B.; Lin, X.; Huang, Q.; Shen, W. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 2010, 326, 511–522. [Google Scholar] [CrossRef]
- Tan, Y.; Wang, J.; He, Y.; Yu, X.; Chen, S.; Penttinen, P.; Liu, S.; Yang, Y.; Zhao, K.; Zou, L. Organic Fertilizers Shape Soil Microbial Communities and Increase Soil Amino Acid Metabolites Content in a Blueberry Orchard. Microb. Ecol. 2022, 85, 232–246. [Google Scholar] [CrossRef]
- Tahat, M.M.; Alananbeh, K.M.; Othman, Y.A.; Leskovar, D.I. Soil Health and Sustainable Agriculture. Sustainability 2020, 12, 4859. [Google Scholar] [CrossRef]
- Doley, K.; Borde, M. Chapter 9—Arbuscular mycorrhizal (AM) fungi: Potential role in sustainable agriculture. Environ. Technol. 2021, 1, 203–225. [Google Scholar]
- Cheng, X.; Wu, H.; Guo, Q.; Hou, B.; Xia, L.; Wang, H.; Diao, Z.; Huo, F.; Ji, W.; Li, R.; et al. Paleomagnetic results of Late Paleozoic rocks from northern Qiangtang Block in Qinghai-Tibet Plateau, China. Sci. China Earth Sci. 2012, 55, 67–75. [Google Scholar] [CrossRef]
- Trisilawati, O.; Hartoyo, B.; Bermawie, N.; Pribadi, E.R. Application of AMF (Arbuscular Mycorrhizal Fungi) and Organic Fertilizer to Increase the Growth, Biomass and Bioactive Content of Centella. IOP Conf. Ser. Earth Environ. Sci. 2019, 292, 012067. [Google Scholar] [CrossRef]
- Dowarah, B.; Gill, S.S.; Agarwala, N. Arbuscular Mycorrhizal Fungi in Conferring Tolerance to Biotic Stresses in Plants. J. Plant Growth Regul. 2022, 41, 1429–1444. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef]
- El Omari, B.; El Ghachtouli, N. Arbuscular mycorrhizal fungi-weeds interaction in cropping and unmanaged ecosystems: A review. Symbiosis 2021, 83, 279–292. [Google Scholar] [CrossRef]
- Kumar, U.; Nayak, A.K.; Shahid, M.; Gupta, V.V.; Panneerselvam, P.; Mohanty, S.; Kaviraj, M.; Kumar, A.; Chatterjee, D.; Lal, B.; et al. Continuous application of inorganic and organic fertilizers over 47 years in paddy soil alters the bacterial community structure and its influence on rice production. Agric. Ecosyst. Environ. 2018, 262, 65–75. [Google Scholar] [CrossRef]
- Lekberg, Y.; Arnillas, C.A.; Borer, E.T.; Bullington, L.S.; Fierer, N.; Kennedy, P.G.; Leff, J.W.; Luis, A.D.; Seabloom, E.W.; Henning, J.A. Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils. Nat. Commun. 2021, 12, 3484. [Google Scholar] [CrossRef]
- Castañares, E.; Stenglein, S.A.; Dinolfo, M.I.; Moreno, M.V. Fusarium tricinctum Associated with Head Blight on Wheat in Argentina. Plant Dis. 2011, 95, 496. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, J.; Gao, J.; Yang, L. First Report of Leaf Spot Disease on Schisandra chinensis Caused by Phoma glomerata in China. Plant Dis. 2012, 96, 289. [Google Scholar] [CrossRef]
- Chen, Q.; Jiang, J.R.; Zhang, G.Z.; Cai, L.; Crous, P. Resolving the Phoma enigma. Stud. Mycol. 2015, 82, 137–217. [Google Scholar] [CrossRef]
- Sun, W.; Fan, J.; Fang, A.; Li, Y.; Tariqjaveed, M.; Li, D.; Hu, D.; Wang, W.-M. Ustilaginoidea virens: Insights into an Emerging Rice Pathogen. Annu. Rev. Phytopathol. 2020, 58, 363–385. [Google Scholar] [CrossRef]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Li, R.; Tao, R.; Ling, N.; Chu, G. Chemical, organic and bio-fertilizer management practices effect on soil physicochemical property and antagonistic bacteria abundance of a cotton field: Implications for soil biological quality. Soil Tillage Res. 2017, 167, 30–38. [Google Scholar] [CrossRef]
- Wang, B.; Yuan, J.; Zhang, J.; Shen, Z.; Zhang, M.; Li, R.; Ruan, Y.; Shen, Q. Effects of novel bioorganic fertilizer produced by Bacillus amyloliquefaciens W19 on antagonism of Fusarium wilt of banana. Biol. Fertil. Soils 2013, 49, 435–446. [Google Scholar] [CrossRef]
- Lazcano, C.; Zhu-Barker, X.; Decock, C. Effects of Organic Fertilizers on the Soil Microorganisms Responsible for N2O Emissions: A Review. Microorganisms 2021, 9, 983. [Google Scholar] [CrossRef]
- Cheng, H.; Zhang, D.; Huang, B.; Song, Z.; Ren, L.; Hao, B.; Liu, J.; Zhu, J.; Fang, W.; Yan, D.; et al. Organic fertilizer improves soil fertility and restores the bacterial community after 1,3-dichloropropene fumigation. Sci. Total. Environ. 2020, 738, 140345. [Google Scholar] [CrossRef]
- Bai, L.; Zhang, X.; Li, B.; Sun, F.; Zhao, X.; Wang, Y.; Lu, Z.; Zhang, D.; Fang, J. Fungal communities are more sensitive to nitrogen fertilization than bacteria in different spatial structures of silage maize under short-term nitrogen fertilization. Appl. Soil Ecol. 2021, 170, 104275. [Google Scholar] [CrossRef]
- Semenov, M.V.; Krasnov, G.S.; Semenov, V.M.; van Bruggen, A. Mineral and Organic Fertilizers Distinctly Affect Fungal Communities in the Crop Rhizosphere. J. Fungi 2022, 8, 251. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, J.; Manuel, D.-B.; de Beeck, M.O.; Shahbaz, M.; Chen, Y.; Deng, X.; Xu, Z.; Li, J.; Liu, Z. Rotation cropping and organic fertilizer jointly promote soil health and crop production. J. Environ. Manag. 2022, 315, 115190. [Google Scholar] [CrossRef]
- Zhang, Y.; Cong, J.; Lu, H.; Yang, C.; Yang, Y.; Zhou, J.; Li, D. An Integrated Study to Analyze Soil Microbial Community Structure and Metabolic Potential in Two Forest Types. PLoS ONE 2014, 9, e93773. [Google Scholar] [CrossRef]
- Röttjers, L.; Faust, K. From hairballs to hypotheses–biological insights from microbial networks. FEMS Microbiol. Rev. 2018, 42, 761–780. [Google Scholar] [CrossRef] [Green Version]
- Li, B.-B.; Roley, S.S.; Duncan, D.S.; Guo, J.; Quensen, J.F.; Yu, H.-Q.; Tiedje, J.M. Long-term excess nitrogen fertilizer increases sensitivity of soil microbial community to seasonal change revealed by ecological network and metagenome analyses. Soil Biol. Biochem. 2021, 160, 108349. [Google Scholar] [CrossRef]
- van der Heijden, M.G.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Huang, G.; Liao, J.; Han, Z.; Li, J.; Zhu, L.; Lyu, G.; Lu, L.; Xie, Y.; Ma, J. Interaction between Fungal Communities, Soil Properties, and the S vival of Invading E. coli O157:H7 in Soils. Int. J. Environ. Res. Public Health 2020, 17, 3516. [Google Scholar] [CrossRef]
- Li, X.; Zhao, Y.; Sun, G.; Jin, W.; Sun, M.; Zhang, H.; Xu, N.; Cai, D.; Li, D. Linking soil bacterial community and crop yield in a wheat (triticum aestivum L.)—Alfalfa (medicago sativa L.) intercropping system. Appl. Ecol. Environ. Res. 2020, 18, 4487–4505. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Community | Area | Planting Pattern | Group | MRPP | ANOSIM | PERMANOVA | |||
|---|---|---|---|---|---|---|---|---|---|
| Delta | p | r | p | Pesudo-F | p | ||||
| Monoculture | G vs. CKG | 0.7194 | 0.025 | 0.288 | 0.041 | 2.2664 | 0.028 | ||
| MG vs. CKMG | 0.6453 | 0.493 | −0.0034 | 0.41 | 0.9052 | 0.607 | |||
| 16S rRNA gene | Rhizosphere | ML vs. CKML | 0.6563 | 0.567 | −0.0232 | 0.569 | 1.0384 | 0.236 | |
| Grass-legume mixture | MG vs. ML | 0.6588 | 0.34 | 0.0400 | 0.183 | 0.9679 | 0.401 | ||
| CKMG vs. CKML | 0.6440 | 0.258 | 0.0078 | 0.31 | 1.0504 | 0.259 | |||
| Bulk soil | Monoculture | BG vs. CKBG | 0.8167 | 0.129 | 0.2533 | 0.073 | 1.7621 | 0.112 | |
| Grass-legume mixture | BF vs. BCK | 0.5278 | 0.01 | 0.0712 | 0.026 | 1.1398 | 0.015 | ||
| Monoculture | G vs. CKG | 0.7660 | 0.79 | −0.0666 | 0.734 | 0.9781 | 0.786 | ||
| MG vs. CKMG | 0.7587 | 0.094 | 0.0545 | 0.139 | 1.0905 | 0.095 | |||
| ITS gene | Rhizphere | ML vs. CKML | 0.7640 | 0.648 | −0.0307 | 0.762 | 0.9800 | 0.452 | |
| Grass-legume mixture | MG vs. ML | 0.7418 | 0.03 | 0.1390 | 0.037 | 1.2312 | 0.026 | ||
| CKMG vs. CKML | 0.7406 | 0.655 | −0.0331 | 0.692 | 0.9629 | 0.626 | |||
| Bulk soil | Monoculture | BG vs. CKBG | 0.7800 | 0.973 | −0.136 | 0.935 | 0.9437 | 0.958 | |
| Grass-legume mixture | BF vs. BCK | 0.7626 | 0.001 | 0.3685 | 0.001 | 1.6687 | 0.001 | ||
| Group | Molecular Ecological Network | Random Network | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Similarity Threshold | Nodes | Links | Average Degree (avgK) | Average Clustering Coefficient (avgCC) | Average Path Distance (GD) | Centralization of Degree (CD) | Density (D) | Transitivity (Trans) | Modularity | R2 | Average Clustering Coefficient (avgCC) | Average Path Distance (GD) | Modularity | |
| MG | 0.970 | 488 | 944 | 3.869 | 0.293 a | 7.16 b | 0.025 | 0.008 | 0.402 | 0.788 c | 0.85 | 0.012 ± 0.003 | 4.324 ± 0.045 | 0.517 ± 0.006 |
| CKMG | 0.970 | 723 | 1324 | 3.663 | 0.259 a | 6.231 b | 0.021 | 0.005 | 0.449 | 0.794 c | 0.838 | 0.008 ± 0.002 | 4.564 ± 0.040 | 0.542 ± 0.004 |
| ML | 0.960 | 517 | 1119 | 4.329 | 0.259 a | 4.394 b | 0.046 | 0.008 | 0.445 | 0.645 c | 0.923 | 0.027 ± 0.004 | 3.869 ± 0.043 | 0.462 ± 0.005 |
| CKML | 0.960 | 404 | 867 | 4.292 | 0.217 a | 4.694 b | 0.049 | 0.011 | 0.364 | 0.626 c | 0.895 | 0.025 ± 0.005 | 3.839 ± 0.047 | 0.465 ± 0.006 |
| BF | 0.890 | 1396 | 1286 | 1.842 | 0.104 a | 16.612 b | 0.007 | 0.001 | 0.265 | 0.959 c | 0.93 | 0.001 ± 0.001 | 9.374 ±0.332 | 0.899 ± 0.004 |
| BCK | 0.890 | 1460 | 1614 | 2.211 | 0.128 a | 14.309 b | 0.011 | 0.002 | 0.294 | 0.925 c | 0.947 | 0.002 ± 0.001 | 7.309 ± 0.119 | 0.806 ± 0.004 |
| MG | 0.890 | 184 | 188 | 2.043 | 0.149 a | 4.924 b | 0.033 | 0.011 | 0.313 | 0.873 c | 0.85 | 0.007 ± 0.006 | 6.188 ± 0.408 | 0.782 ± 0.011 |
| CKMG | 0.890 | 217 | 328 | 3.023 | 0.227 a | 8.712 b | 0.037 | 0.014 | 0.382 | 0.809 c | 0.806 | 0.015 ± 0.007 | 4.649 ± 0.102 | 0.601 ± 0.010 |
| ML | 0.860 | 261 | 270 | 2.069 | 0.188 a | 3.577 b | 0.023 | 0.008 | 0.381 | 0.937 c | 0.852 | 0.005 ± 0.004 | 7.173 ± 0.401 | 0.802 ± 0.009 |
| CKML | 0.860 | 323 | 488 | 3.022 | 0.26 a | 6.732 b | 0.034 | 0.009 | 0.323 | 0.8 c | 0.87 | 0.011 ± 0.004 | 4.925± 0.073 | 0.616 ± 0.007 |
| BF | 0.830 | 307 | 481 | 3.134 | 0.235 a | 10.505 b | 0.059 | 0.01 | 0.376 | 0.811 c | 0.864 | 0.013 ± 0.005 | 4.575 ± 0.076 | 0.594 ± 0.007 |
| BN | 0.830 | 346 | 494 | 2.855 | 0.205 a | 6.815 b | 0.035 | 0.008 | 0.296 | 0.802 c | 0.865 | 0.009 ± 0.004 | 4.972 ± 0.094 | 0.640 ± 0.007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, H.; Zhou, X.; Zheng, K.; Gu, S.; Yu, H.; Ma, K.; Zhao, Y.; Wang, Y.; Zheng, H.; Liu, H.; et al. Response of Organic Fertilizer Application to Soil Microorganisms and Forage Biomass in Grass–Legume Mixtures. Agronomy 2023, 13, 481. https://doi.org/10.3390/agronomy13020481
Yan H, Zhou X, Zheng K, Gu S, Yu H, Ma K, Zhao Y, Wang Y, Zheng H, Liu H, et al. Response of Organic Fertilizer Application to Soil Microorganisms and Forage Biomass in Grass–Legume Mixtures. Agronomy. 2023; 13(2):481. https://doi.org/10.3390/agronomy13020481
Chicago/Turabian StyleYan, Huilin, Xueli Zhou, Kaifu Zheng, Songsong Gu, Hao Yu, Kun Ma, Yangan Zhao, Yingcheng Wang, Hua Zheng, Hanjiang Liu, and et al. 2023. "Response of Organic Fertilizer Application to Soil Microorganisms and Forage Biomass in Grass–Legume Mixtures" Agronomy 13, no. 2: 481. https://doi.org/10.3390/agronomy13020481