

Elevated Temperature Affects Avena sterilis ssp. ludoviciana Reproductive Biology

Abstract
:1. Introduction
2. Materials and Methods
2.1. Biotypes Used
2.2. Experimental Design and Treatment Set Up of Pot Trial
2.3. Data Collection and Spikelet Storage
2.4. Dormancy Tests
2.5. Seed Longevity Determined Using Controlled Ageing Test
2.6. Thermal Time Calculation
2.7. Statistical Analysis
3. Results
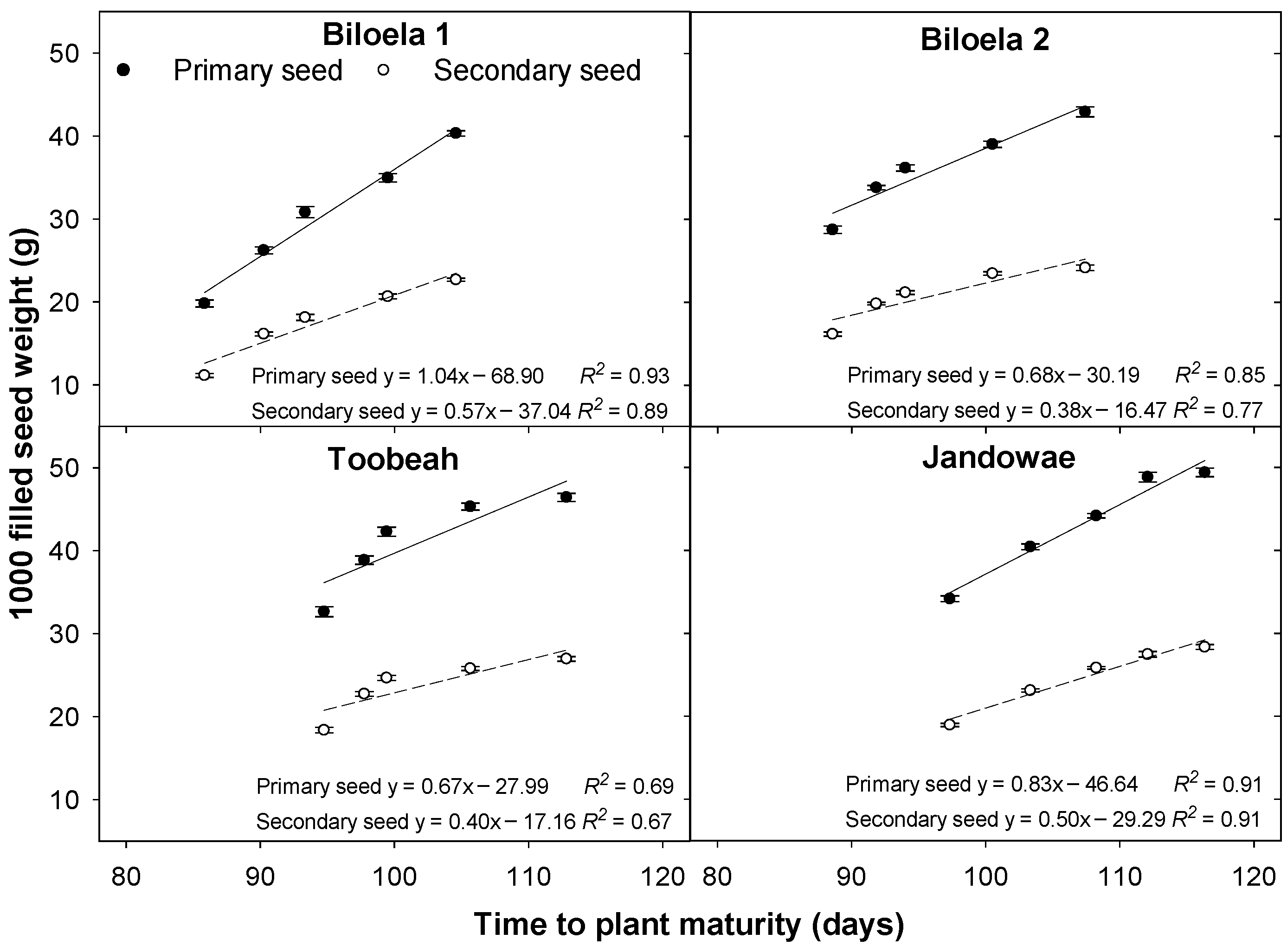
3.1. Time to Plant Maturity
3.2. Seeds Produced
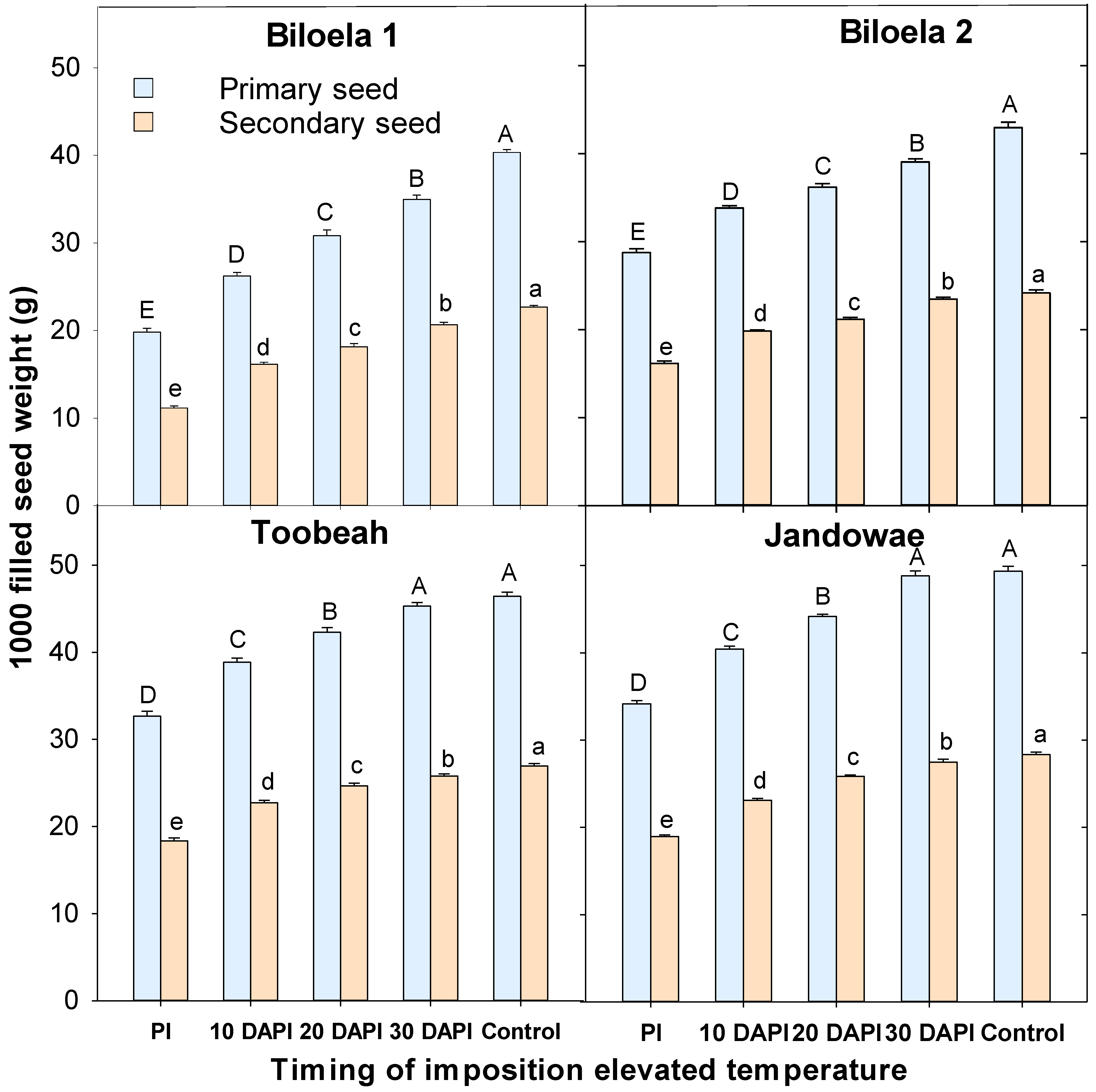
3.3. 1000 Primary and Secondary Seed Weight
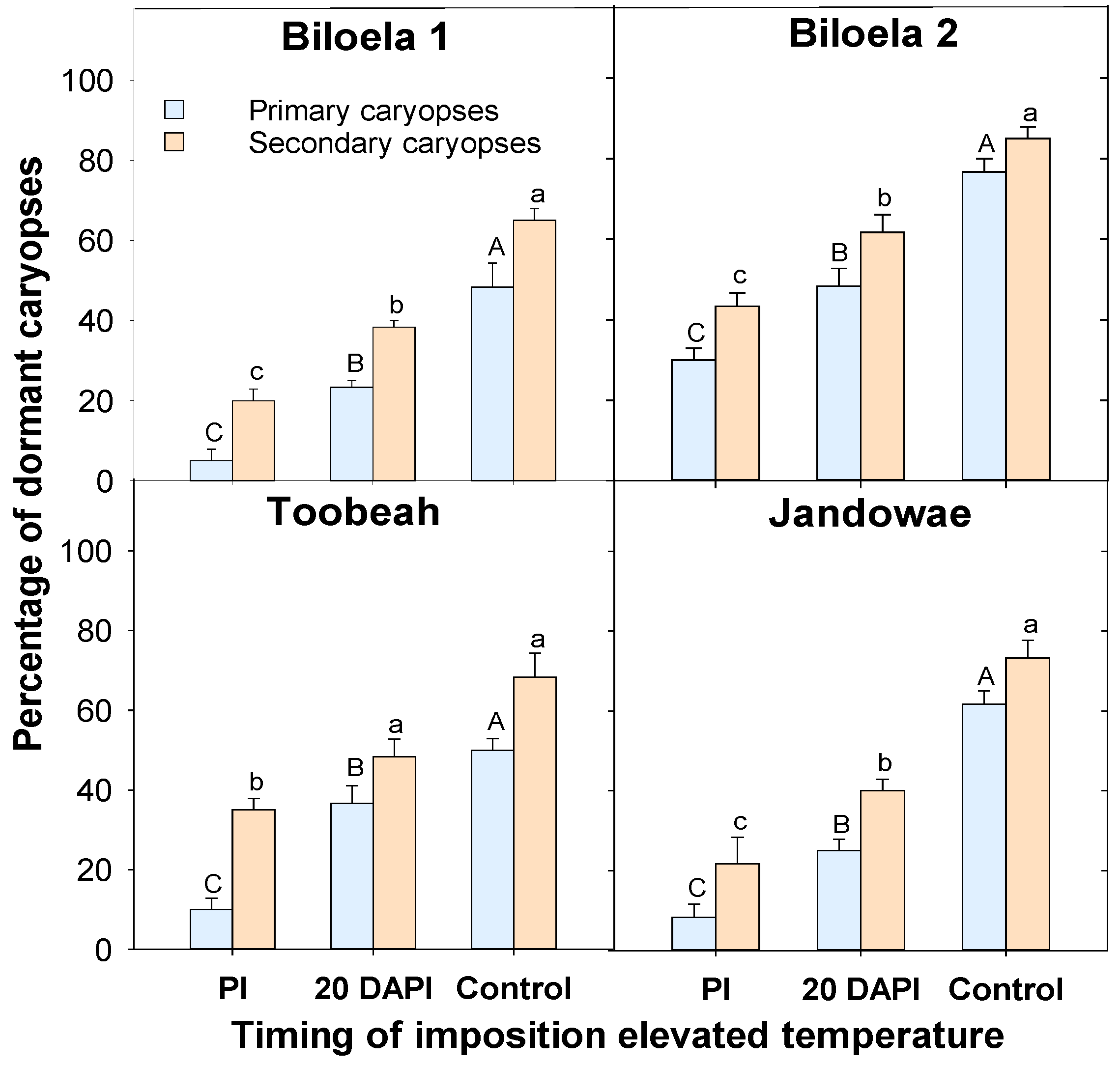
3.4. Dormancy Test of Seeds in the T-Bar
3.5. Dormancy Test of Caryopses in the Germination Incubator
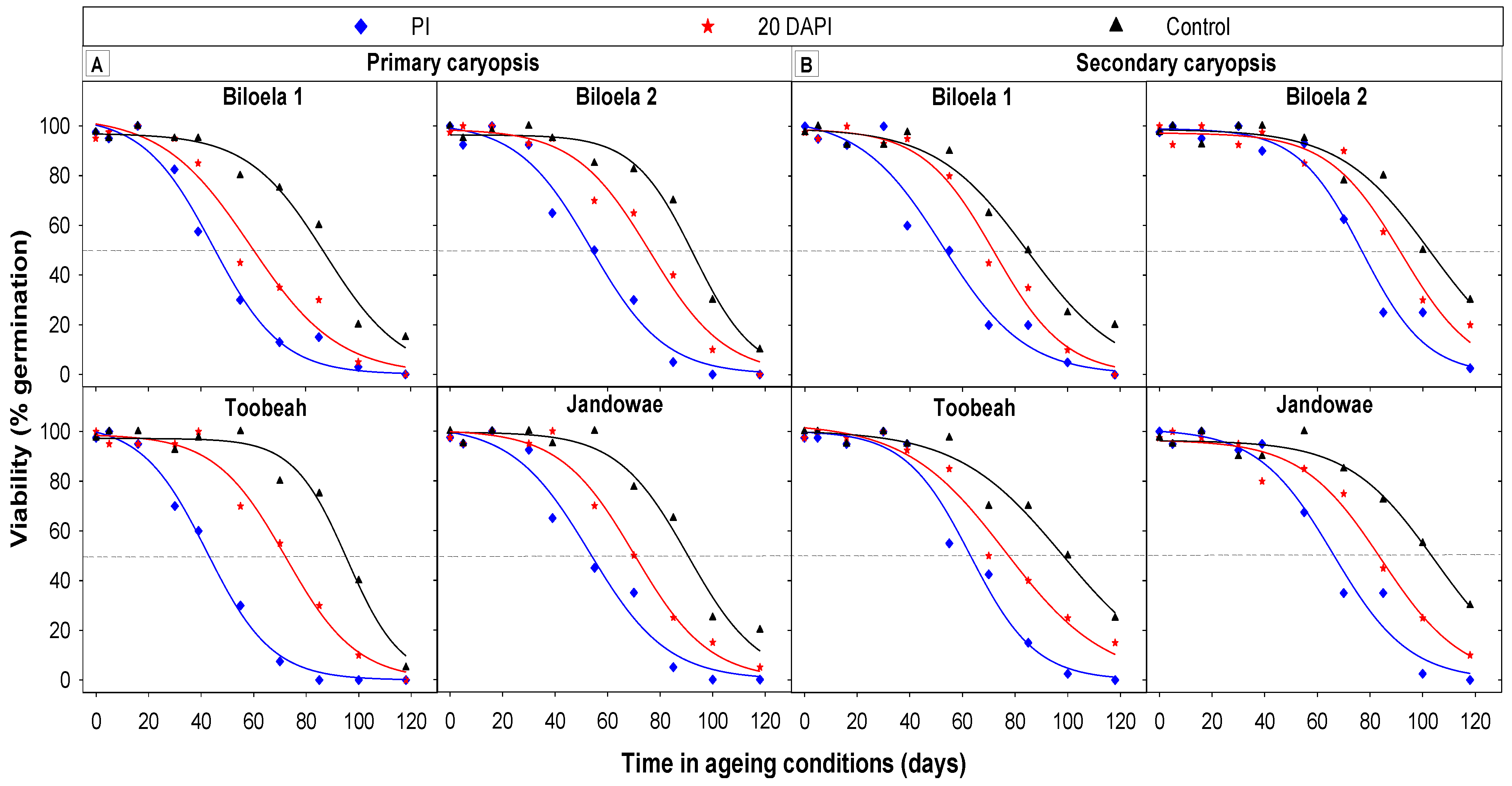
3.6. Seed Longevity Determined by CAT
4. Discussion
4.1. Phenology
4.2. Reproductive Biology
4.3. Dormancy Status
4.4. Longevity Status
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Llewellyn, R.S.; Ronning, D.; Ouzman, J.; Walker, S.; Mayfield, A.; Clarke, M. Impact of Weeds on Australian Grain Production: The Cost of Weeds to Australian Grain Growers and the Adoption of Weed Management and Tillage Practices; Report for GRDC; Grains Research and Development Corporation, and the Commonwealth Scientific and Industrial Research Organization: Canberra, Australia, 2016. [Google Scholar]
- Thomas, G.A.; Titmarsh, G.W.; Freebairn, D.M.; Radford, B.J. No-tillage and conservation farming practices in grain growing areas of Queensland—A review of 40 years of development. Aust. J. Exp. Agric. 2007, 47, 887–898. [Google Scholar] [CrossRef]
- Llewellyn, R.; Ouzman, J. Conservation agriculture in Australia: 30 years. In Australian Agriculture in 2020: From Conservation to Automation; Pratley, J., Kirkegaard, J., Eds.; Agronomy Australia and Charles Sturt University: Wagga Wagga, Australia, 2015; pp. 21–32. [Google Scholar]
- Walsh, M.; Broster, J.; Chauhan, B.; Rebetzke, G.; Pratley, J. Weed control in cropping systems—Past lessons and future opportunities. In Australian Agriculture in 2020: From Conservation to Automation; Pratley, J., Kirkegaard, J., Eds.; Agronomy Australia and Charles Sturt University: Wagga Wagga, Australia, 2015; pp. 153–172. [Google Scholar]
- Dang, Y.P.; Moody, P.W.; Bell, M.J.; Seymour, N.P.; Dalal, R.C.; Freebairn, D.M.; Walker, S.R. Strategic tillage in no-till farming systems in Australia’s north-eastern grains-growing regions: II. Implications for agronomy, soil and environment. Soil Tillage Res. 2015, 152, 115–123. [Google Scholar] [CrossRef]
- Sexsmith, J.J. Dormancy of wild oat seed produced under various temperature and moisture conditions. Weed Sci. 1969, 17, 405–407. [Google Scholar] [CrossRef]
- Peters, N.C.B. Production and dormancy of wild oat (Avena fatua) seed from plants grown under soil water stress. Annals Appl. Biol. 1982, 100, 189–196. [Google Scholar]
- Peters, N.C.B. The dormancy of wild oat seed (Avena fatua L.) from plants grown under various temperature and soil moisture conditions. Weed Res. 1982, 22, 205–212. [Google Scholar] [CrossRef]
- Sawhney, R.; Naylor, J.M. Dormancy studies in seed of Avena fatua. 13. Influence of drought stress during seed development on duration of seed dormancy. Can. J. Bot. 1982, 60, 1016–1020. [Google Scholar] [CrossRef]
- Sawhney, R.; Quick, W.A.; Hsiao, A.L. The effect of temperature during parental vegetative growth on seed germination of wild oats (Avena fatua L.). Ann. Bot. 1985, 55, 25–28. [Google Scholar] [CrossRef]
- Adkins, S.W.; Loewen, M.; Symons, S.J. Variation within pure lines of wild oats (Avena fatua) in relation to temperature of development. Weed Sci. 1987, 35, 169–172. [Google Scholar] [CrossRef]
- O’Donnell, C.; Adkins, S.W. Wild oat and climate change: The effect of CO2 concentration, temperature, and water deficit on the growth and development of wild oat in monoculture. Weed Sci. 2001, 49, 694–702. [Google Scholar] [CrossRef]
- Naylor, J.M.; Jana, S. Genetic adaptation for seed dormancy in Avena fatua. Can. J. Bot. 1976, 54, 306–312. [Google Scholar] [CrossRef]
- Jana, S.; Naylor, J.M. Dormancy studies in seed of Avena fatua. 11. Heritability for seed dormancy. Can. J. Bot. 1980, 58, 91–93. [Google Scholar] [CrossRef]
- Medd, R.W. Ecology of wild oats. Plant Prot. Q. 1996, 11, 185–187. [Google Scholar]
- Gallandt, E.R.; Fuerst, E.P.; Kennedy, A.C. Effect of tillage, fungicide seed treatment, and soil fumigation on seed bank dynamics of wild oat (Avena fatua). Weed Sci. 2004, 52, 597–604. [Google Scholar] [CrossRef]
- Somody, C.N.; Nalewaja, J.D.; Miller, S.D. The response of wild oat (Avena fatua) and Avena sterilis accessions to photoperiod and temperature. Weed Sci. 1984, 32, 206–213. [Google Scholar] [CrossRef]
- Del Arco, M.S.; Torner, C.; Quintanilla, C.F. Seed dynamics in populations of Avena sterilis ssp. ludoviciana. Weed Res. 1995, 35, 477–487. [Google Scholar] [CrossRef]
- Foley, M.E. Genetic model for dormancy in wild oat. In Seed Biology: Advances and Applications; Black, M., Bradford, K.J., Vazquez-Ramos, J., Eds.; CABI Publishing: Wallingford, UK, 1999; pp. 323–327. [Google Scholar]
- Quail, P.H.; Carter, O.G. Survival and seasonal germination of seeds of Avena fatua and A. ludoviciana. Aust. J. Agric. Res. 1968, 19, 721–729. [Google Scholar] [CrossRef]
- BOM and CSIRO (Bureau of Meteorology; Centre for Scientific and Industrial Research Organization). State of the Climate. 2020. Available online: http://www.bom.gov.au/state-of-the-climate/ (accessed on 15 July 2021).
- Lobell, D.B.; Hammer, G.L.; Chenu, K.; Zheng, B.; McLean, G.; Chapman, S.C. The shifting influence of drought and heat stress for crops in northeast Australia. Glob. Chang. Biol. 2015, 21, 4115–4127. [Google Scholar] [CrossRef]
- Whalley, R.D.B.; Burfitt, J.M. Ecotypic variation in Avena fatua L., A. sterilis L. (A. ludoviciana), and A. barbata Pott. in New South Wales and southern Queensland. Aust. J. Agric. Res. 1972, 23, 799–810. [Google Scholar] [CrossRef]
- Nugent, T.; Storrie, A.; Medd, R. Managing Wild Oats; CRC for Weed Management Systems and Grains Research and Development Corporation: Adelaide, SA, Australia; Canberra, ACT, Australia, 1999. [Google Scholar]
- Ali, M.; Suthar, P.C.; Williams, A.; Widderick, M.; Adkins, S.W. Germination behaviour of Avena sterilis ssp. ludoviciana under a range of light and temperature. Crop Pasture Sci. 2022, 73, 1395–1405. [Google Scholar] [CrossRef]
- Soil Survey Staff. Kellogg Soil Survey Laboratory Methods Manual. In Soil Survey Investigations; Burt, R., Ed.; Soil Report No. 42, Version 5.0; Survey Staff; United States Department of Agriculture, Natural Resources Conservation Service: Washington, DC, USA, 2014; pp. 140–144. [Google Scholar]
- Lancashire, P.D.; Bleiholder, H.; Boom, T.V.D.; Langeluddeke, P.; Stauss, R.; Weber, E.; Witzenber, A. A uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Long, R.L.; Panetta, F.D.; Steadman, K.J.; Probert, R.; Bekker, R.M.; Brooks, S.; Adkins, S.W. Seed Persistence in the field may be predicted by Laboratory-Controlled Aging. Weed Sci. 2008, 56, 523–528. [Google Scholar] [CrossRef]
- McMaster, G.S.; Wilhelm, W.W. Growing degree-days: One equation, two interpretations. Agric. For. Meteor. 1997, 87, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Richards, R.A.; Hunt, J.R.; Kirkegaard, J.A.; Passioura, J.B. Yield improvement and adaptation of wheat to water-limited environments in Australia—A case study. Crop Pasture Sci. 2014, 65, 676–689. [Google Scholar] [CrossRef]
- Barnabas, B.; Jager, K.; Feher, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Jagadish, S.V.K.; Septiningsih, E.M.; Kohli, A.; Thomson, M.J.; Ye, C.; Redona, E.; Kumar, A.; Gregorio, G.B.; Wassmann, R.; Ismail, A.M.; et al. Genetic Advances in Adapting Rice to a Rapidly Changing Climate. J. Agron. Crop Sci. 2012, 198, 360–373. [Google Scholar] [CrossRef]
- Shah, F.; Nie, L.; Cui, K.; Tariq, S.; Wei, W.; Chang, C.; Zhu, L.; Ali, F.; Fahad, S.; Huang, J. Rice grain yield and component responses to near 2 °C of warming. Field Crops Res. 2014, 157, 98–110. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Saud, S.; Khan, F.; Hassan, S.; Amanullah; Nasim, W.; Arif, M.; Wang, F.; Huang, J. Exogenously applied plant growth regulators affect heat-stressed rice pollens. J. Agron. Crop Sci. 2016, 202, 139–150. [Google Scholar] [CrossRef]
- Shirtliffe, S.J.; Entz, M.H.; Van Acker, R.C. Avena fatua development and seed shatter as related to thermal time. Weed Sci. 2000, 48, 555–560. [Google Scholar] [CrossRef]
- Ferris, R.; Ellis, R.; Wheeler, T.; Hadley, P. Effect of high temperature stress at anthesis on grain yield and biomass of field-grown crops of wheat. Annals Bot. 1998, 82, 631–639. [Google Scholar] [CrossRef]
- Lobell, D.B.; Gourdji, S.M. The influence of climate change on global crop productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef]
- Bahuguna, R.N.; Solis, C.A.; Shi, W.; Jagadish, K.S.V. Post-flowering night respiration and altered sink activity account for high night temperature-induced grain yield and quality loss in rice (Oryza sativa). Physiol. Plant 2016, 159, 59–73. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Djanaguiraman, M.; Narayanan, S.; Erdayani, E.; Prasad, P.V.V. Effects of high temperature stress during anthesis and grain filling periods on photosynthesis, lipids and grain yield in wheat. BMC Plant Biol. 2020, 20, 268. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Huang, B.; Rachmilevitch, S.; Xu, J. Root carbon and protein metabolism associated with heat tolerance. J. Exp. Bot. 2012, 63, 3455–3465. [Google Scholar] [CrossRef] [PubMed]
- Aisthorpe, D.; McCosker, E. Optimising the phenology and grain yield of wheat genotypes-Emerald. In Queensland Grains Research 2018–19; Regional Research Agronomy, Queensland Department of Agriculture and Fisheries, and Grains Research and Development Corporation, State of Queensland, Australia: Brisbane, Australia, 2019; pp. 7–14. [Google Scholar]
- Shah, F.; Nie, L.; Cui, K.; Shah, T.; Wu, W.; Chen, C.; Zhu, L.; Ali, F.; Fahad, S.; Huang, J. A biochar application protects rice pollen from high-temperature stress. Plant Physiol. Biochem. 2015, 96, 281–287. [Google Scholar]
- Prasad, P.V.V.; Bheemanahalli, R.; Jagdish, S.K. Field crops and the fear of heat stress-opportunities, challenges and future directions. Field Crops Res. 2017, 200, 114–121. [Google Scholar] [CrossRef]
- Naylor, J.M.; Fedec, P. Dormancy studies in seed of Avena fatua. 8. Genetic diversity affecting response to temperature. Can. J. Bot. 1978, 56, 2224–2229. [Google Scholar] [CrossRef]
- Fernandez-Quintanilla, C.; Gonzalez Andujar, J.L.; Appleby, A.P. Characterization of the germination and emergence response to temperature and soil moisture of Avena fatua and A. sterilis. Weed Res. 1990, 30, 289–295. [Google Scholar] [CrossRef]
- Uremis, I.; Uygur, F. Minimum, optimum and maximum germination temperatures of some important weed species in the Çukurova Region of Turkey. Türk. Herbol. Derg. 1999, 2, 1–12. [Google Scholar]
- Gallagher, R.S.; Kristen, L.G.; Lidewij, H.K.; Jairus, R.; Dennis, P.; Sebastian, R.; Burnham, M.; Fuerst, E.P. Shade and drought stress-induced changes in phenolic content of wild oat (Avena fatua L.) seeds. J. Stress Physiol. Biochem. 2010, 6, 90–107. [Google Scholar]
- Jones, R.; Medd, R. Economic analysis of integrated management of wild oats involving fallow, herbicide and crop rotational options. Aust. J. Exp. Agric. 1997, 37, 683–691. [Google Scholar] [CrossRef]
- O’Donnell, C.C. The Response of Avena fatua to the Enhanced Greenhouse Effect. Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2002. [Google Scholar]
- Sharma, A.K.; McBeath, D.K.; Vanden Born, W.H. Studies on the biology of wild oats. I. dormancy, germination and emergence. Can. J. Plant Sci. 1976, 56, 611–618. [Google Scholar] [CrossRef]
- Morrison, I.N.; Dushnicky, L. Structure of the covering layers of the wild oat (Avena fatua) caryopsis. Weed Sci. 1982, 30, 352–359. [Google Scholar] [CrossRef]
- Bekker, R.M.; Bakker, J.P.; Ozinga, W.A.; Thompson, K. Seed traits: Essential for understanding seed longevity. Asp. Appl. Biol. 2003, 69, 1–9. [Google Scholar]
- Martin, R.J.; Felton, W.L. Effect of crop rotation, tillage practice, and herbicides on the population dynamics of wild oats in wheat. Aust. J. Exp. Agric. 1993, 33, 159–165. [Google Scholar] [CrossRef]
- Peltzer, S.C.; Matson, P.T. How fast do the seedbanks of five annual cropping weeds deplete in the absence of weed seed input? In Proceedings of the 13th Australian Weeds Conference, Plant Protection Society of WA, Perth, Australia, 8–13 September 2002; pp. 553–555. [Google Scholar]
- Storrie, A.M. (Ed.) Integrated Weed Management in Australian Cropping Systems, 2nd ed.; Grains Research and Development Corporation: Canberra, Australia, 2014. [Google Scholar]
- Mahajan, G.; Chauhan, B.S. Seed longevity and seedling emergence behavior of wild oat (Avena fatua) and sterile oat (Avena sterilis ssp. ludoviciana) in response to burial depth in eastern Australia. Weed Sci. 2021, 69, 362–371. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevated Temperature Treatments | at Panicle Initiation (PI) | 10 Days after Panicle Initiation (10 DAPI) | 20 Days after Panicle Initiation (20 DAPI) | 30 Days after Panicle Initiation (30 DAPI) | |
|---|---|---|---|---|---|
| Biotypes | Days to Reach These Stages of Development | ||||
| Northern zone | Biloela 1 | 58 | 68 | 78 | 88 |
| Biloela 2 | 58 | 68 | 78 | 88 | |
| Southern zone | Toobeah | 63 | 73 | 83 | 93 |
| Jandowae | 65 | 75 | 85 | 95 | |
| Treatment | Biotype | Caryopsis Type | a | b | P50 | R2 |
|---|---|---|---|---|---|---|
| PI | Biloela 1 | Primary | 102 | 12 | 45 | 0.98 |
| Secondary | 104 | 16 | 53 | 0.95 | ||
| Biloela 2 | Primary | 101 | 14 | 54 | 0.98 | |
| Secondary | 99 | 12 | 77 | 0.97 | ||
| Toobeah | Primary | 103 | 12 | 42 | 0.99 | |
| Secondary | 100 | 12 | 63 | 0.98 | ||
| Jandowae | Primary | 102 | 15 | 53 | 0.97 | |
| Secondary | 101 | 14 | 66 | 0.97 | ||
| 20 DAPI | Biloela 1 | Primary | 97 | 9 | 61 | 0.99 |
| Secondary | 99 | 14 | 72 | 0.98 | ||
| Biloela 2 | Primary | 99 | 14 | 76 | 0.97 | |
| Secondary | 97 | 14 | 92 | 0.96 | ||
| Toobeah | Primary | 99 | 13 | 72 | 0.98 | |
| Secondary | 103 | 19 | 76 | 0.97 | ||
| Jandowae | Primary | 100 | 14 | 70 | 0.98 | |
| Secondary | 97 | 15 | 84 | 0.97 | ||
| Control | Biloela 1 | Primary | 111 | 15 | 87 | 0.97 |
| Secondary | 99 | 18 | 85 | 0.97 | ||
| Biloela 2 | Primary | 96 | 11 | 93 | 0.98 | |
| Secondary | 99 | 17 | 103 | 0.95 | ||
| Toobeah | Primary | 97 | 10 | 96 | 0.97 | |
| Secondary | 100 | 20 | 98 | 0.95 | ||
| Jandowae | Primary | 99 | 13 | 91 | 0.96 | |
| Secondary | 96 | 16 | 105 | 0.96 |
| Biotype | PI | 20 DAPI | Control | |||
|---|---|---|---|---|---|---|
| P50 Value (Days) | Predicted Longevity in the Seedbank (Years) | P50 Value (Days) | Predicted Longevity in the Seedbank (Years) | P50 Value (Days) | Predicted Longevity in the Seedbank (Years) | |
| Primary caryopses/seed | ||||||
| Biloela 1 | 41 | 1 to 2 | 62 | >2 to 4 | 78 | >2 to 4 |
| Biloela 2 | 60 | 1 to 2 | 78 | >2 to 4 | 91 | >4 |
| Toobeah | 48 | 1 to 2 | 71 | >2 to 4 | 82 | >4 |
| Jandowae | 45 | 1 to 2 | 66 | >2 to 4 | 84 | >4 |
| Secondary caryopses/seed | ||||||
| Biloela 1 | 57 | 1 to 2 | 69 | >2 to 4 | 83 | >4 |
| Biloela 2 | 74 | >2 to 4 | 89 | >4 | 102 | >4 |
| Toobeah | 66 | >2 to 4 | 82 | >4 | 95 | >4 |
| Jandowae | 63 | >2 to 4 | 77 | >2 to 4 | 97 | >4 |
| Treatment | Days to Panicle Initiation | Thermal Time until Plants Exposed to Elevated Temperature (Degree-Days) | Days to Maturity | Thermal Time until Maturity (Degree-Days) | ||||
|---|---|---|---|---|---|---|---|---|
| 2018–2019 | 2019–2020 | 2-Years Average | 2018–2019 | 2019–2020 | 2-Years Average | |||
| Biloela 1 | ||||||||
| PI | 58 | 852 | 884 | 868 | 86 | 1580 | 1612 | 1596 |
| 10 DAPI | 1032 | 1063 | 1048 | 90 | 1604 | 1635 | 1620 | |
| 20 DAPI | 1224 | 1236 | 1230 | 93 | 1614 | 1626 | 1620 | |
| 30 DAPI | 1421 | 1437 | 1429 | 100 | 1733 | 1749 | 1741 | |
| Control | 852 | 884 | 868 | 105 | 1795 | 1795 | 1795 | |
| Biloela 2 | ||||||||
| PI | 58 | 852 | 884 | 868 | 89 | 1658 | 1690 | 1674 |
| 10 DAPI | 1032 | 1063 | 1048 | 92 | 1656 | 1687 | 1672 | |
| 20 DAPI | 1224 | 1236 | 1230 | 94 | 1640 | 1652 | 1646 | |
| 30 DAPI | 1421 | 1437 | 1429 | 101 | 1759 | 1775 | 1767 | |
| Control | 852 | 884 | 868 | 107 | 1837 | 1837 | 1837 | |
| Toobeah | ||||||||
| PI | 63 | 946 | 965 | 956 | 95 | 1778 | 1797 | 1788 |
| 10 DAPI | 1124 | 1147 | 1136 | 98 | 1774 | 1797 | 1786 | |
| 20 DAPI | 1315 | 1332 | 1324 | 99 | 1731 | 1748 | 1740 | |
| 30 DAPI | 1515 | 1546 | 1531 | 106 | 1853 | 1884 | 1869 | |
| Control | 946 | 965 | 956 | 113 | 1980 | 1980 | 1980 | |
| Jandowae | ||||||||
| PI | 65 | 978 | 1002 | 990 | 97 | 1810 | 1834 | 1822 |
| 10 DAPI | 1166 | 1180 | 1173 | 103 | 1894 | 1908 | 1901 | |
| 20 DAPI | 1356 | 1375 | 1366 | 108 | 1954 | 1973 | 1964 | |
| 30 DAPI | 1561 | 1583 | 1572 | 112 | 2003 | 2025 | 2014 | |
| Control | 978 | 1002 | 990 | 116 | 2059 | 2039 | 2049 | |
| Location | Year | Monthly Average Temperature (°C) | 3-Month Average Temperature (°C) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| August | September | October | ||||||||
| Max. | Min. | Max. | Min. | Max. | Min. | Max. | Min. | |||
| Northern NGR | Biloela | 2015 | 25.4 | 6.3 | 27.3 | 8.2 | 30.9 | 12.9 | 27.8 | 9.1 |
| 2016 | 24.0 | 7.5 | 27.0 | 13.0 | 30.3 | 11.6 | 27.1 | 10.7 | ||
| 2017 | 26.1 | 6.6 | 30.3 | 9.1 | 29.2 | 17.6 | 28.5 | 11.1 | ||
| Northern NGR’s 3-year average | 25.2 | 6.8 | 28.2 | 10.1 | 30.1 | 14.0 | 27.8 | 10.3 | ||
| Southern NGR | Toobeah | 2015 | 21.9 | 5.8 | 26 | 7.9 | 33.7 | 16.9 | 27.2 | 10.2 |
| 2016 | 21.3 | 5.5 | 23.0 | 10.2 | 27.5 | 11.7 | 23.9 | 9.1 | ||
| 2017 | 23.9 | 3.6 | 28.9 | 8.4 | 29.6 | 15.0 | 27.5 | 9.0 | ||
| 3-year average | 22.4 | 5.0 | 26.0 | 8.8 | 30.3 | 14.5 | 26.2 | 9.4 | ||
| Jandowae | 2015 | 21.7 | 3.9 | 24.5 | 6.8 | 29.7 | 12.3 | 25.3 | 7.7 | |
| 2016 | 20.7 | 5.8 | 21.9 | 9.9 | 27.5 | 9.6 | 23.4 | 8.4 | ||
| 2017 | 23.8 | 5.3 | 29.7 | 8.1 | 27.2 | 15.6 | 26.9 | 9.7 | ||
| 3-year average | 22.1 | 5.0 | 25.4 | 8.3 | 28.1 | 12.5 | 25.2 | 8.6 | ||
| Southern NGR’s 3-year average | 22.2 | 5.0 | 25.8 | 8.7 | 29.2 | 13.5 | 25.7 | 9.0 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.; Williams, A.; Widderick, M.; Adkins, S. Elevated Temperature Affects Avena sterilis ssp. ludoviciana Reproductive Biology. Agronomy 2023, 13, 474. https://doi.org/10.3390/agronomy13020474
Ali M, Williams A, Widderick M, Adkins S. Elevated Temperature Affects Avena sterilis ssp. ludoviciana Reproductive Biology. Agronomy. 2023; 13(2):474. https://doi.org/10.3390/agronomy13020474
Chicago/Turabian StyleAli, Mohammad, Alwyn Williams, Michael Widderick, and Steve Adkins. 2023. "Elevated Temperature Affects Avena sterilis ssp. ludoviciana Reproductive Biology" Agronomy 13, no. 2: 474. https://doi.org/10.3390/agronomy13020474