
Management Effect on the Weed Control Efficiency in Double Cropping Systems
 , , , and
, , , and
Abstract
:1. Introduction
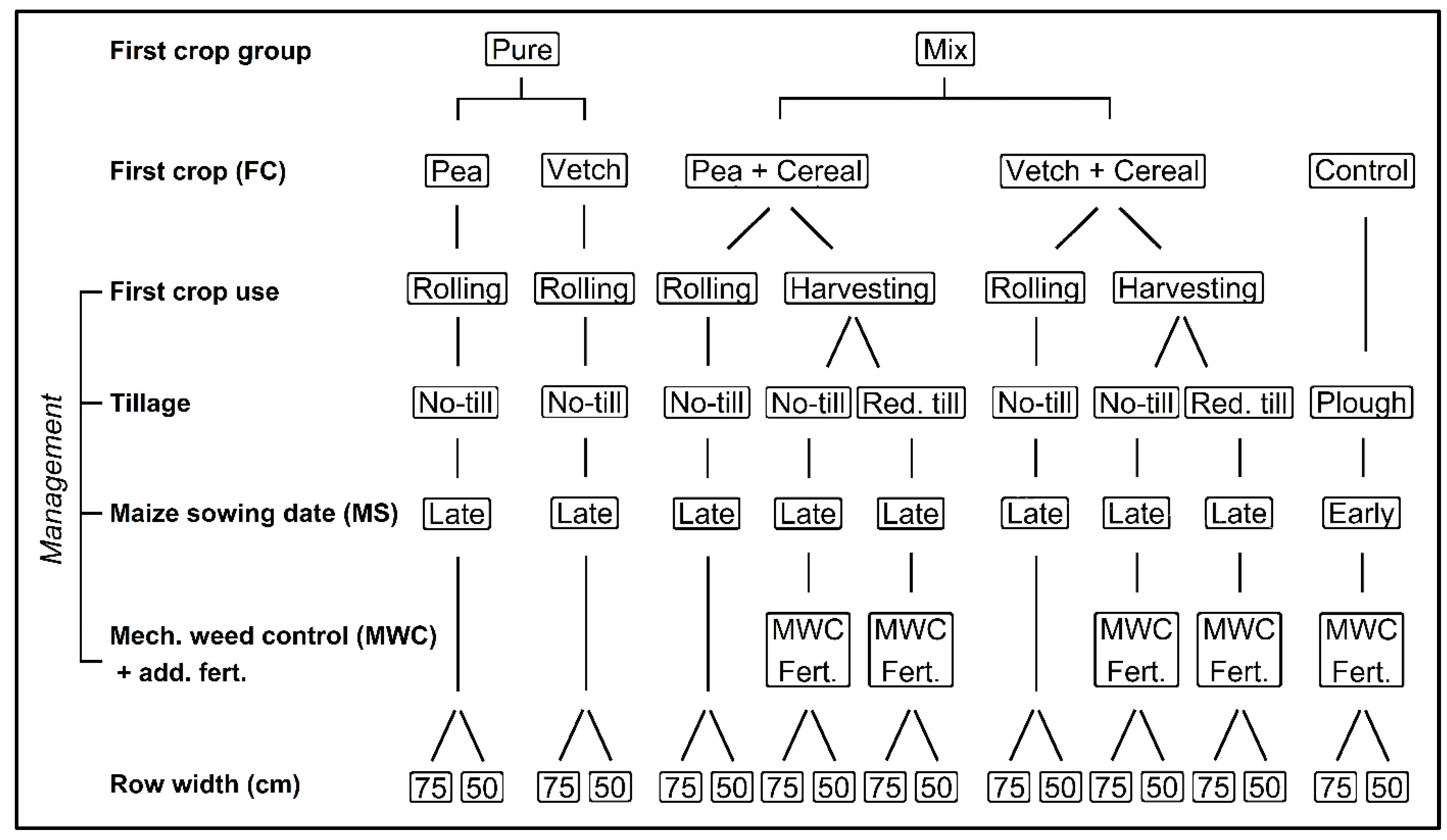
2. Materials and Methods
3. Results
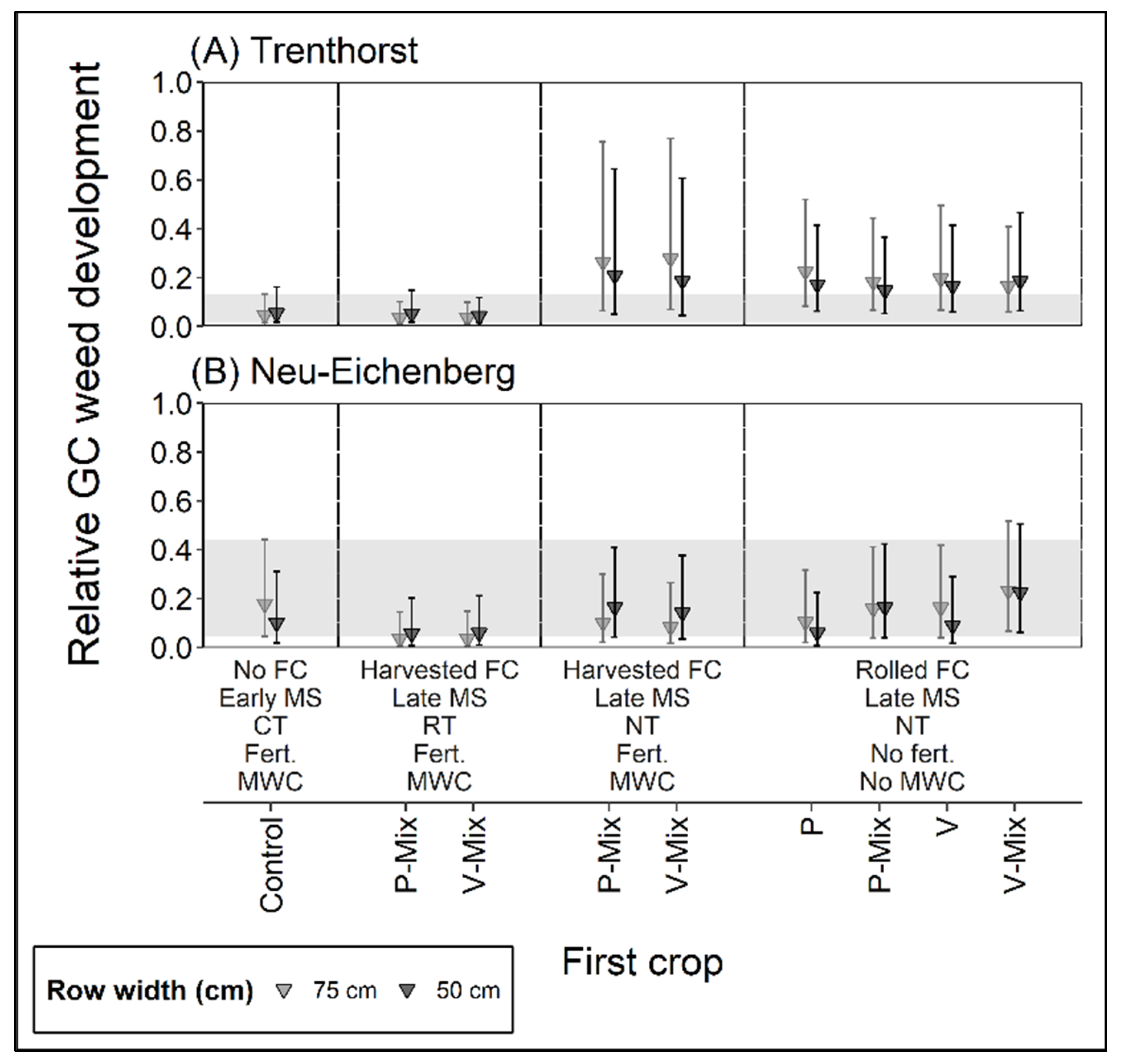
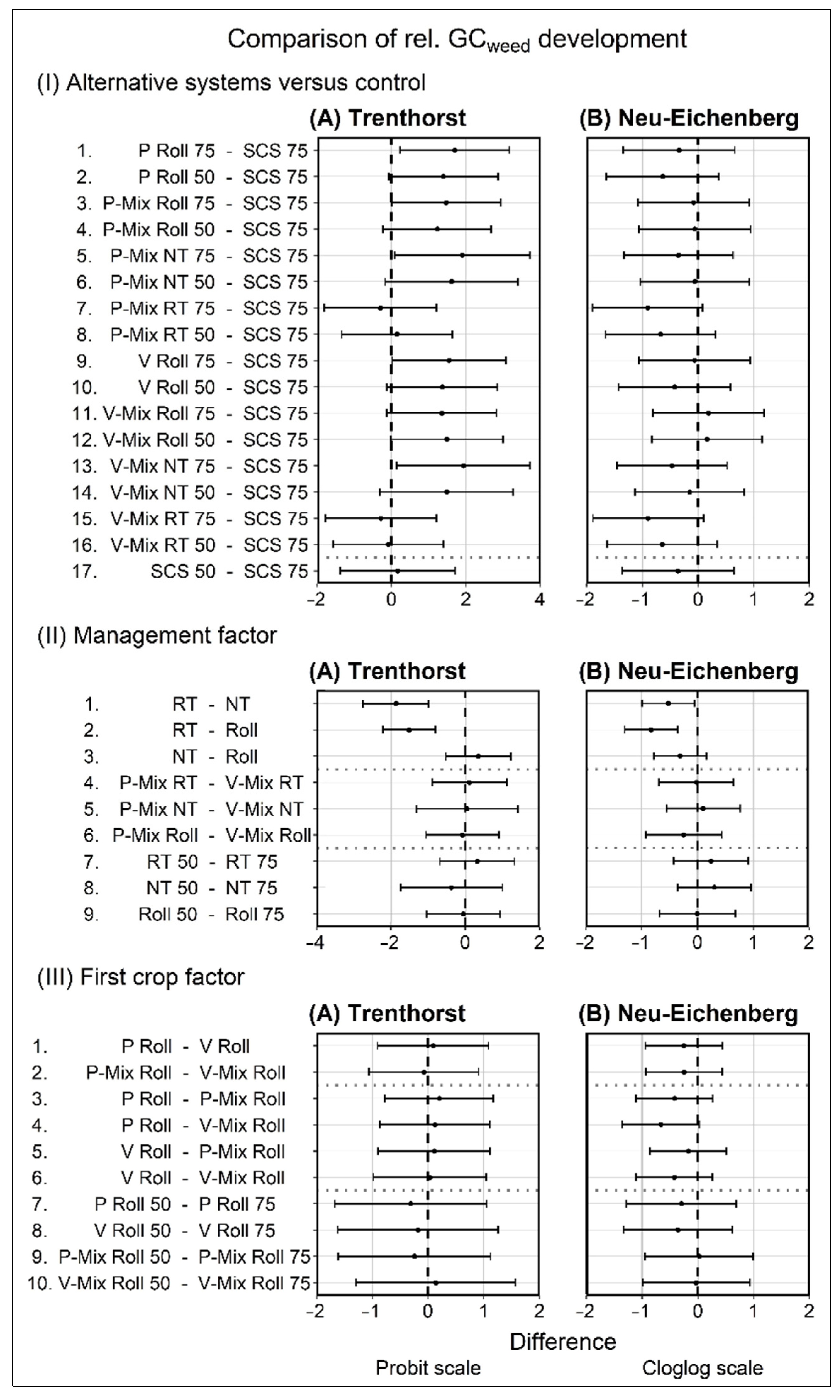
3.1. Relative Weed Groundcover Development
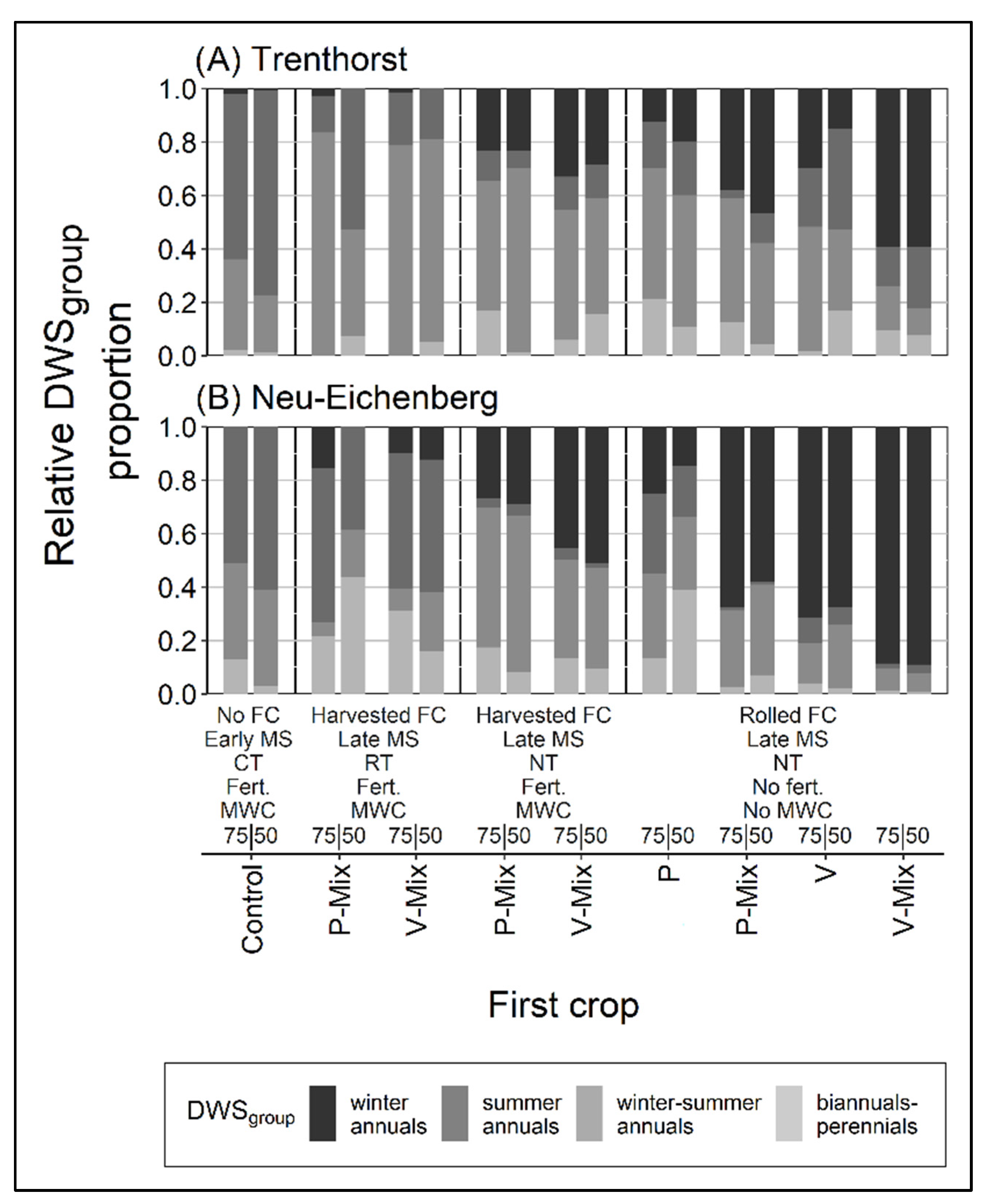
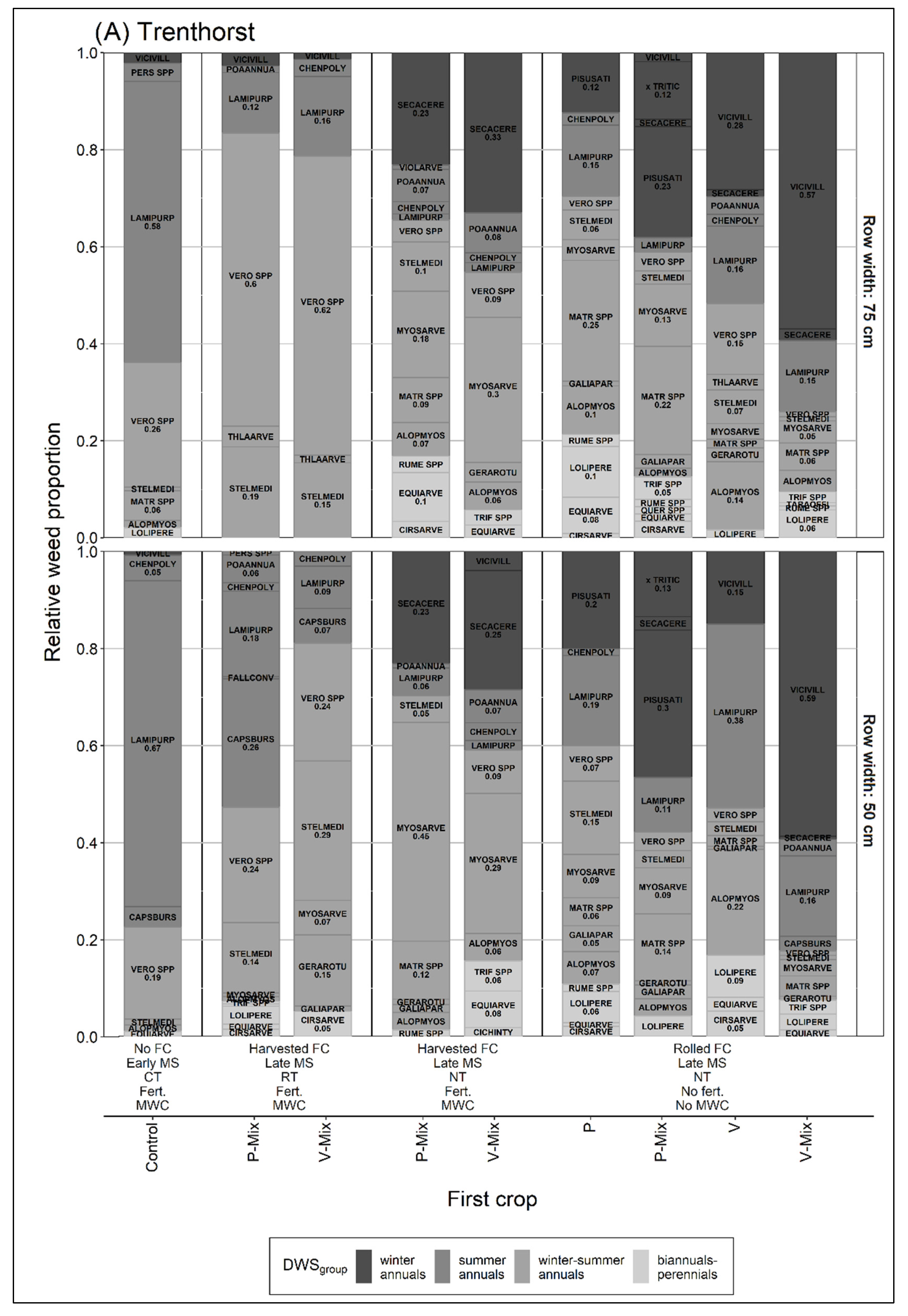
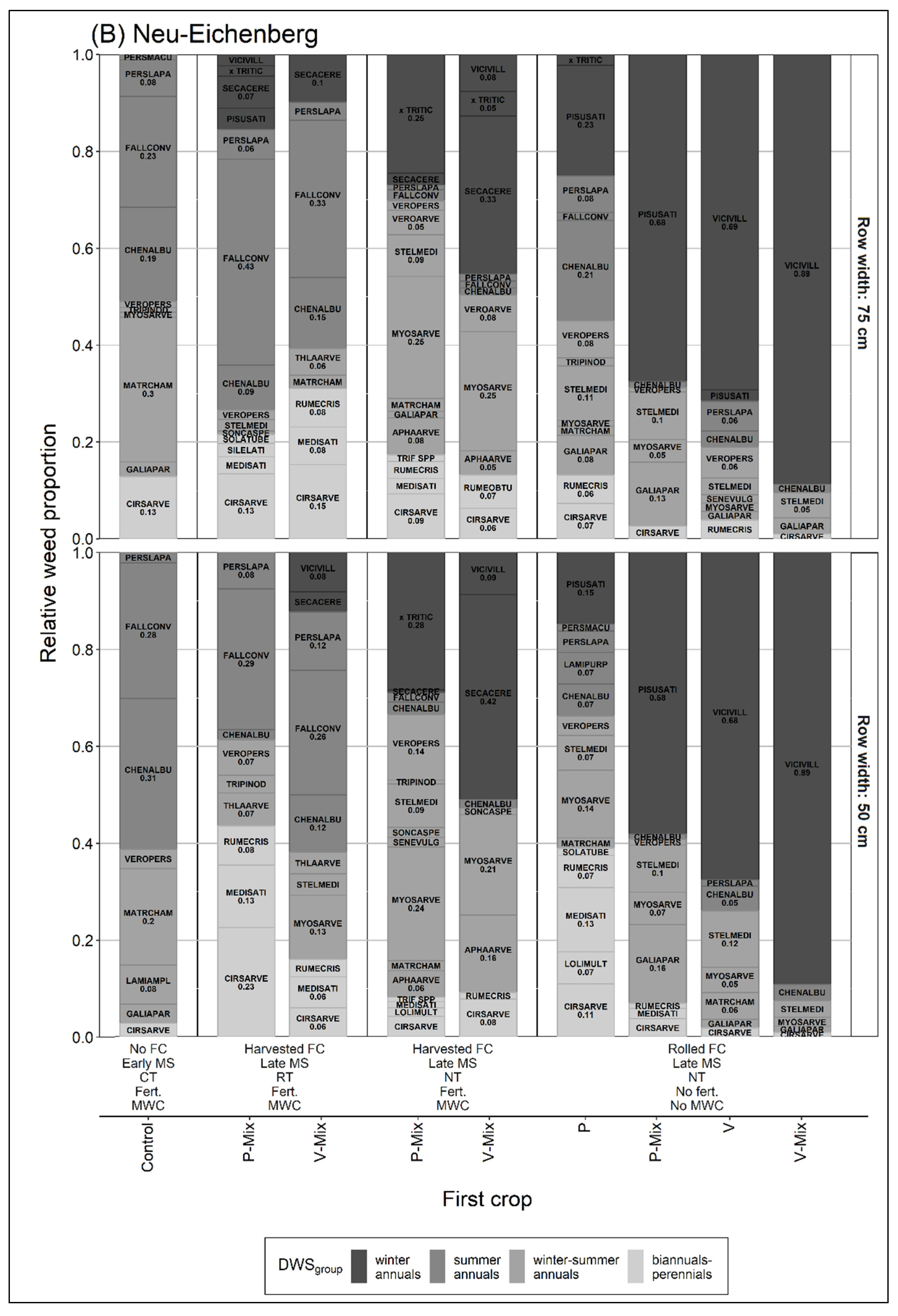
3.2. Relative Proportion of Dominant Weed Species Groups over the Season
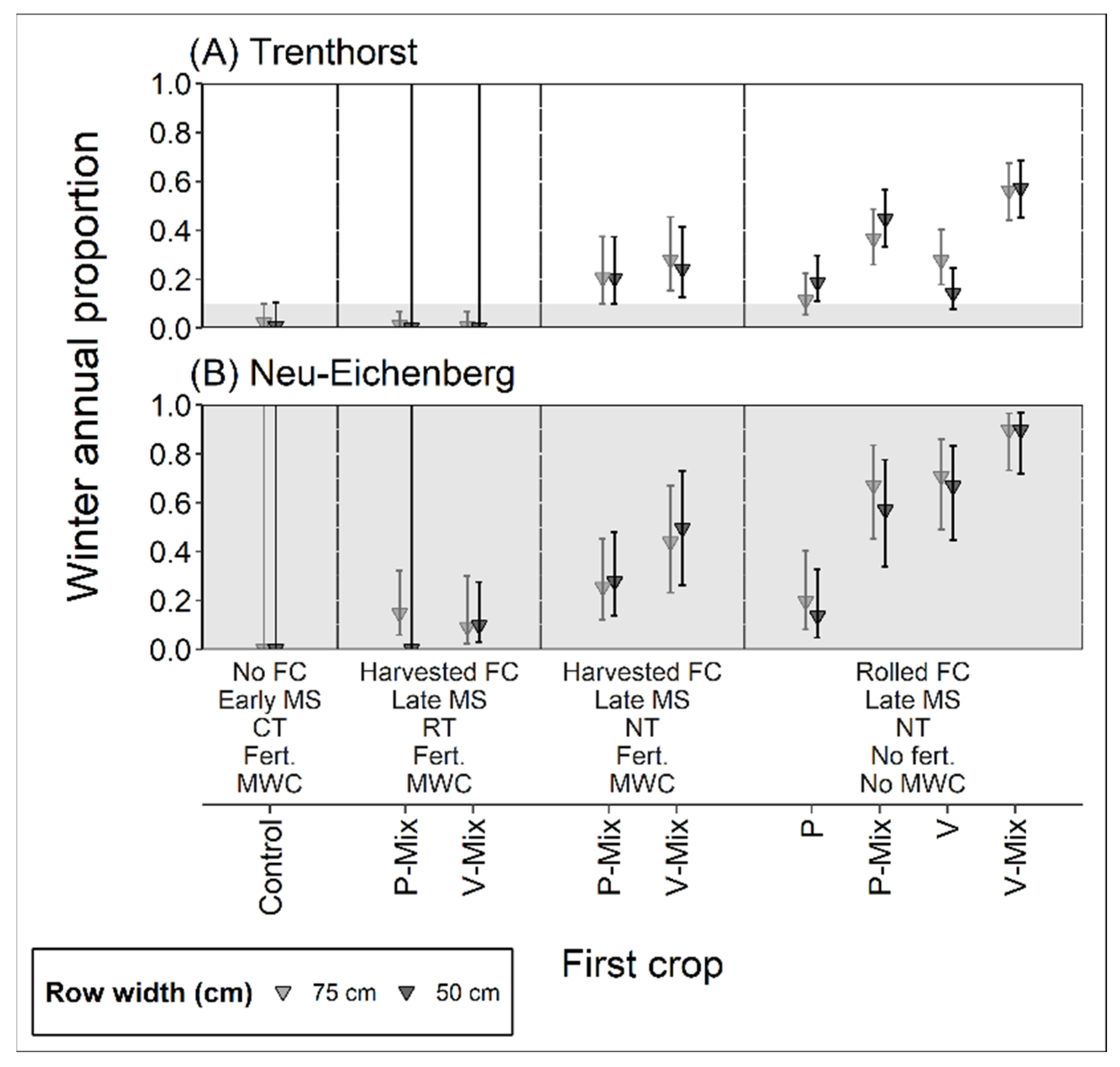
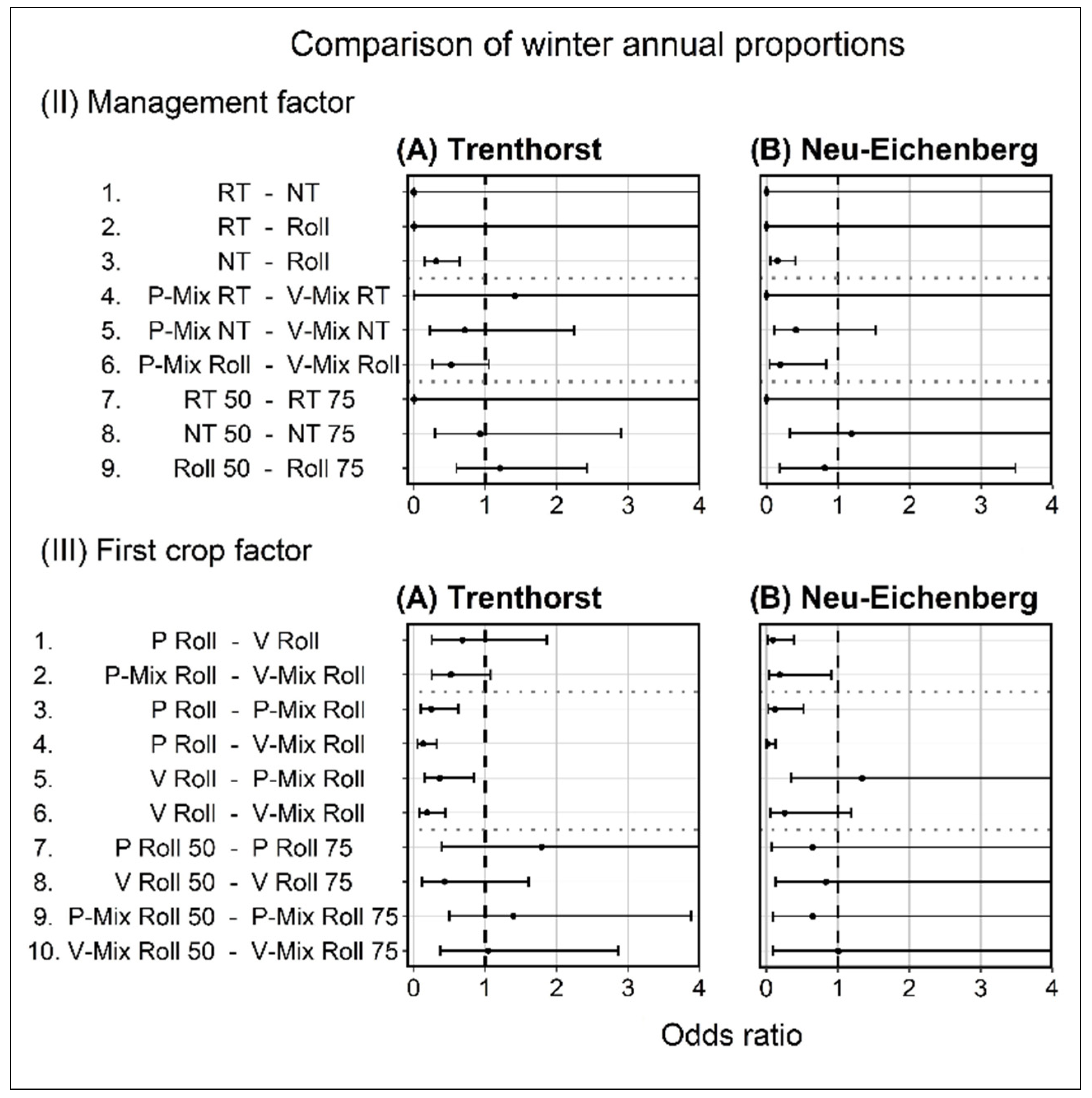
3.2.1. Relative Proportion of Winter Annuals
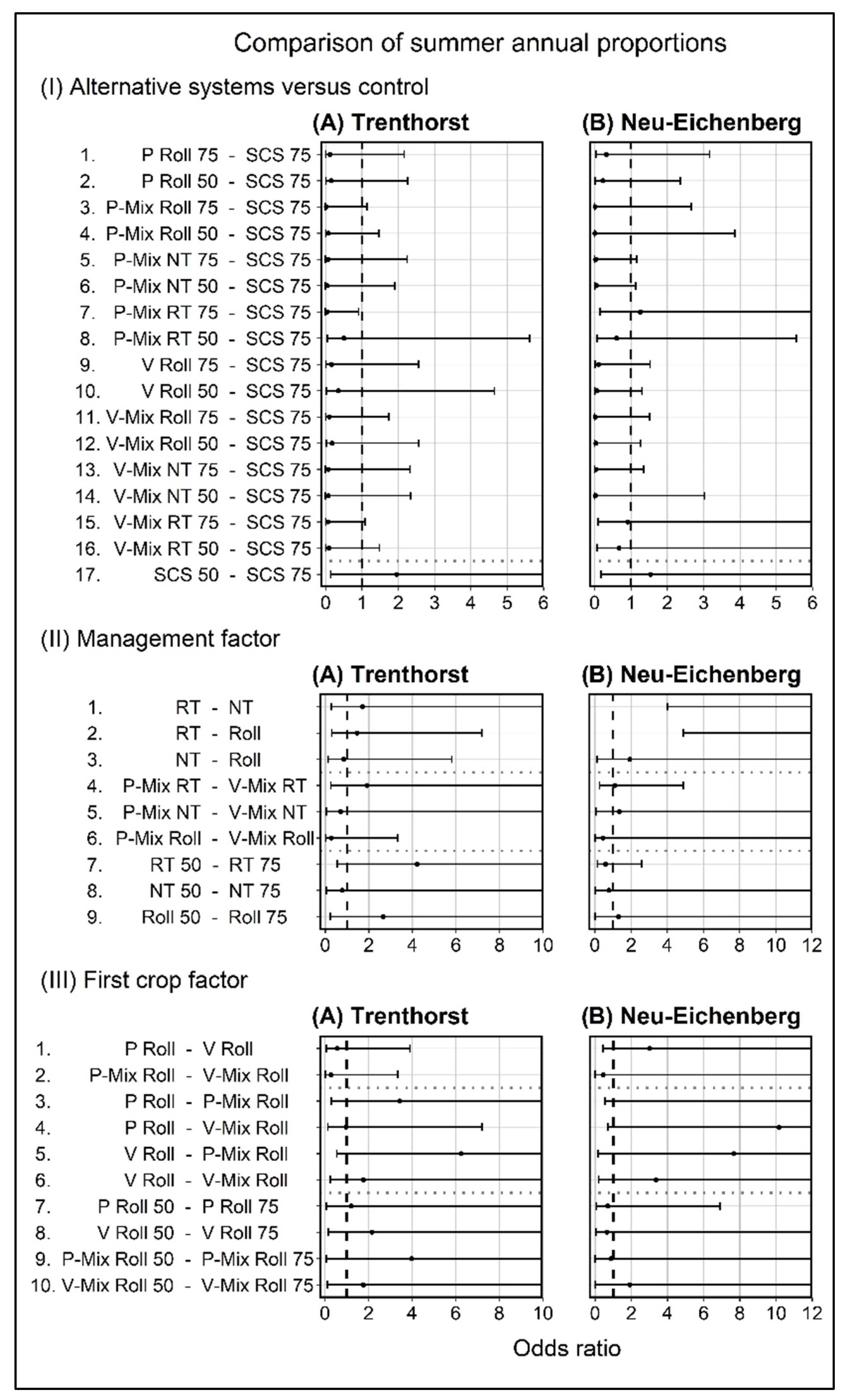
3.2.2. Relative Proportion of Summer and Winter-Summer Annuals
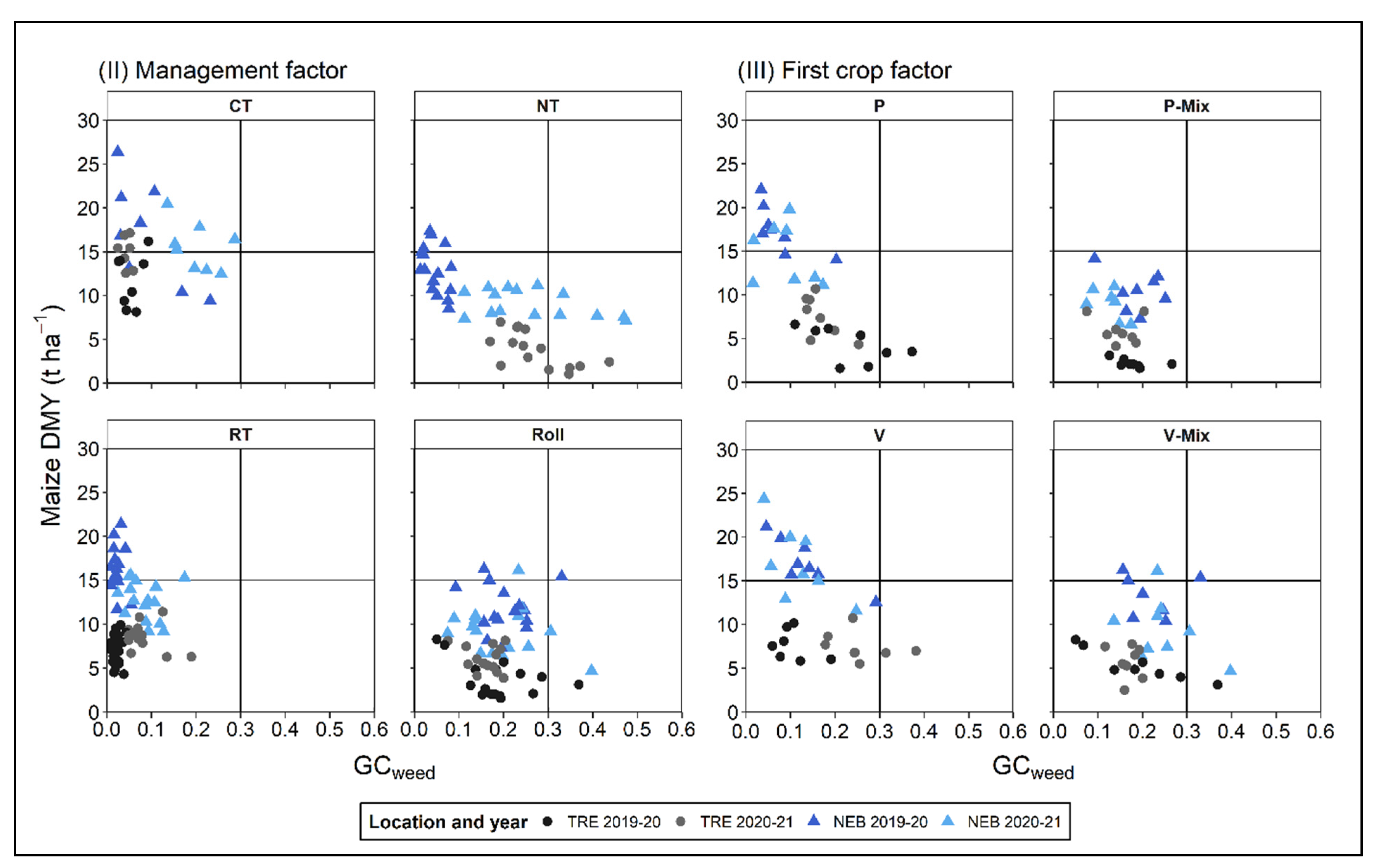
3.3. Relationship between Weed Groundcover and Maize Dry Matter Yield
4. Discussion
4.1. Relative Weed Groundcover Development
4.2. Relative Proportion of Dominant Weed Species
4.2.1. Relative Proportion of Winter Annuals
4.2.2. Relative Proportion of Summer and Winter-Summer Annuals
4.3. Relationship between Weed Groundcover and Maize Dry Matter Yield
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Year | Distinct Factors | t0 | t1 | t2 | t3 | t4 | t5 | t6 |
|---|---|---|---|---|---|---|---|---|---|
| TRE | 2019–20 | SCS | 0 | 57 | 71 | 85 | |||
| DCS, DCMS | 0 | 10 | 24 | 38 | 52 | 66 | 80 | ||
| 2020–21 | SCS | 0 | 14 | 29 | 40 | 54 | 70 | 85 | |
| DCS | 0 | 8 | 23 | 34 | 48 | 64 | 79 | ||
| DCMS | 0 | 9 | 20 | 34 | 50 | 65 | |||
| NEB | 2019–20 | SCS | 0 | 36.5 | 50 | ||||
| SCS, row 1 | 0 | 13.5 | 28 | 41.5 | 55.5 | ||||
| DCS, DCMS | 0 | 13.5 | 28 | 41.5 | 55.5 | 75.5 | |||
| 2020–21 | SCS | 0 | 48 | 63 | 76.5 | 90.5 | |||
| DCS 75 cm | 0 | 23 | 38 | 51.5 | 65.5 | ||||
| DCMS | 0 | 15 | 30 | 43.5 | 57.5 | ||||
| DCS 50 cm | 0 | 10 | 25 | 38.5 | 52.5 |
Appendix B
| Location | DWSgroup | Proportion of Zero Values (%) | Proportion of One Values (%) | Model Iterations |
|---|---|---|---|---|
| TRE | winter annuals | 50 | 0 | 980 |
| summer annuals | 38 | 7 | 4 | |
| winter-summer annuals | 31 | 4 | 4 | |
| biannuals-perennials | 70 | 0 | 980 | |
| NEB | winter annuals | 45 | 7 | 981 |
| summer annuals | 46 | 5 | 3 | |
| winter-summer annuals | 42 | 3 | 981 | |
| biannuals-perennials | 58 | 1 | 895 |
Appendix C
| Life form (DWSgroup) | Location (Total Species) | DWS (%) | Percent of Total (%) |
|---|---|---|---|
| winter annuals | TRE (4) | Vicia villosa Roth (10), Pisum sativum L. (6), × Triticosecale Wittm. ex A. Camus (4), Secale cereale L. (3) | 23 |
| NEB (4) | Vi. villosa (16), Pi. sativum (9), Se. cereale (7), × Triticosecale (5) | 37 | |
| summer annuals | TRE (5 + 2) | Lamium purpureum L. (19), Poa annuaa L. (2), Chenopodium polyspermum L. (1), Persicaria spp. b (1), Capsella bursa-pastoris c (L.) Medik. (1), Fallopia convolvulus (L.) Á. Löve (<1), Viola arvensis Murray (<1) | 24 |
| NEB (3 + 4) | Fa. convolvulus (10), Chenopodium album L. (5), Persicaria lapathifolia (L.) Gray (4), Pe. maculosa Gray (<1),Ca. bursa-pastorisc (<1), La. purpureum (<1), Vi. arvensis (<1) | 19 | |
| winter-summer annuals | TRE (6 + 2) | Veronica spp. d (10), Matricaria spp. e (10), Myosotis arvensis f (L.) Hill (8), Stellaria media (L.) Vill. (7), Alopecurus myosuroides Huds. (5), Galium aparine L. (1),Geranium rotundifolium L. (<1), Thlaspi arvense L. (<1) | 41 |
| NEB (9 + 4) | My. Arvensisf (10), St. media (5), Aphanes arvensis L. (3), Ga. aparine (3), Matricaria chamomilla L. (2), Veronica persica Poir. (2), Ve. arvensis L. (1), Tripleurospermum inodorum (L.) Sch. Bip. (1), Th. arvense (1), Sonchus asper (L.) Hill (<1), Senecio vulgaris L. (<1), Lamium amplexicaule L. (<1), So. oleraceus L. (<1) | 28 | |
| biannuals-perennials | TRE (4 + 4) | Lolium perenne L. (6), Equisetum arvense L. (1), Trifolium spp. g (1), Cirsium arvense (L.) Scop. (1), Rumex spp. (<1), Quercus spp. (<1), Cichorium intybus L. (<1), Taraxacum officinale F.H. Wigg. (<1) | 9 |
| NEB (3 + 6) | Ci. arvense (8), Medicago sativa h L. (2),Rumex crispus L. (2), Solanum tuberosum L. (<1), Trifolium spp. (<1), Lolium multiflorum i Lam. (<1), Ru. obtusifolius L. (<1), Silene latifolia Poir. (<1), Ta. officinale (<1) | 12 |


Appendix D


Appendix E

References
- Graß, R.; Heuser, F.; Stülpnagel, R.; Piepho, H.P.; Wachendorf, M. Energy crop production in double-cropping systems: Results from an experiment at seven sites. Eur. J. Agron. 2013, 51, 120–129. [Google Scholar] [CrossRef]
- FNR. Bioenergy in Germany: Facts and Figures 2020; Fachagentur Nachwachsende Rohstoffe e.V. (FNR): Gülzow-Prüzen, Germany, 2019. [Google Scholar]
- Schmidt, F.; Böhm, H.; Piepho, H.; Urbatzka, P.; Wachendorf, M.; Graß, R. Management Effects on the Performance of Double Cropping Systems—Results from a Multi-Site Experiment. Agronomy 2022, 12, 2104. [Google Scholar] [CrossRef]
- Reckleben, Y. Cultivation of maize—Which sowing row distance is needed? Landtechnik 2011, 66, 370–372. [Google Scholar]
- Carr, P.M.; Mäder, P.; Creamer, N.G.; Beeby, J.S. Editorial: Overview and comparison of conservation tillage practices and organic farming in Europe and North America. Renew. Agric. Food Syst. 2012, 27, 2–6. [Google Scholar] [CrossRef]
- Graß, R.; Scheffer, K. Direkt- und Spätsaat von Silomais nach Wintererbsenvorfrucht—Erfahrungen aus Forschung und Praxis. In Proceedings of the 7. Wissenschaftstagung zum Ökologischen Landbau, Wien, Austria, 23–26 February 2003; pp. 45–48. [Google Scholar]
- Peigné, J.; Lefèvre, V.; Vian, J.F.; Fleury, P. Conservation agriculture in organic farming: Experiences, challenges and opportunities in Europe. In Conservation Agriculture; Farooq, M., Siddique, K.H.M., Eds.; Springer: New York, NY, USA; London, UK, 2015; pp. 559–578. [Google Scholar]
- Herrmann, A. Biogas Production from Maize: Current State, Challenges and Prospects. 2. Agronomic and Environmental Aspects. Bioenergy Res. 2013, 6, 372–387. [Google Scholar] [CrossRef]
- Reicosky, D.C.; Sauer, T.J.; Hatfield, J.L. Challenging Balance between Productivity and Environmental Quality: Tillage Impacts. In Soil Management: Building a Stable Base for Agriculture; Hatfield, J.L., Sauer, T.J., Eds.; American Society of Agronomy Soil Science Society of America: Madison, WI, USA, 2011; pp. 13–37. ISBN 9780891181958. [Google Scholar]
- Finckh, M.R. Integration of breeding and technology into diversification strategies for disease control in modern agriculture. Eur. J. Plant Pathol. 2008, 121, 399–409. [Google Scholar] [CrossRef]
- MEA Food. Ecosystems and Human Well-Being: Current State and Trends; Balisacan, A.M., Gardiner, P., Eds.; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Döring, T.F.; Vieweger, A.; Pautasso, M.; Vaarst, M.; Finckh, M.R.; Wolfe, M.S. Resilience as a universal criterion of health. J. Sci. Food Agric. 2015, 95, 455–465. [Google Scholar] [CrossRef]
- Wolfe, M.S.; Baresel, J.P.; Desclaux, D.; Goldringer, I.; Kovács, G.; Löschenberger, F.; Miedaner, T.; Østergård, H.; Lammerts van Bueren, E.T. Developments in breeding cereals for organic agriculture. Euphytica 2008, 163, 323–346. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Marín, C.; Weiner, J. Effects of density and sowing pattern on weed suppression and grain yield in three varieties of maize under high weed pressure. Weed Res. 2014, 54, 467–474. [Google Scholar] [CrossRef]
- Mhlanga, B.; Chauhan, B.S.; Thierfelder, C. Weed management in maize using crop competition: A review. Crop Prot. 2016, 88, 28–36. [Google Scholar] [CrossRef]
- Peigné, J.; Ball, B.C.; Roger-Estrade, J.; David, C. Is conservation tillage suitable for organic farming? A review. Soil Use Manag. 2007, 23, 129–144. [Google Scholar] [CrossRef]
- Snapp, S.S.; Swinton, S.M.; Labarta, R.; Mutch, D.; Black, J.R.; Leep, R.; Nyiraneza, J.; O’Neil, K. Evaluating cover crops for benefits, costs and performance within cropping system niches. Agron. J. 2005, 97, 322–332. [Google Scholar] [CrossRef]
- Dabney, S.M.; Delgado, J.A.; Reeves, D.W. Using winter cover crops to improve soil and water quality. Commun. Soil Sci. Plant Anal. 2001, 32, 1221–1250. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C.; Bailey, B.A. Role of cover crops in improving soil and row crop productivity. Commun. Soil Sci. Plant Anal. 2005, 36, 2733–2757. [Google Scholar] [CrossRef]
- Holderbaum, J.F.; Decker, A.M.; Messinger, J.J.; Mulford, F.R.; Vough, L.R. Fall-Seeded Legume Cover Crops for No-Tillage Corn in the Humid East. Agron. J. 1990, 82, 117–124. [Google Scholar] [CrossRef]
- Parr, M.; Grossman, J.M.; Reberg-Horton, S.C.; Brinton, C.; Crozier, C. Nitrogen delivery from legume cover crops in no-till organic corn production. Agron. J. 2011, 103, 1578–1590. [Google Scholar] [CrossRef]
- Videnović, Ž.; Simić, M.; Srdić, J.; Dumanović, Z. Long term effects of different soil tillage systems on maize (Zea mays L.) yields. Plant Soil Environ. 2011, 57, 186–192. [Google Scholar] [CrossRef]
- Krauss, M.; Berner, A.; Burger, D.; Wiemken, A.; Niggli, U.; Mäder, P. Reduced tillage in temperate organic farming: Implications for crop management and forage production. Soil Use Manag. 2010, 26, 12–20. [Google Scholar] [CrossRef]
- Nichols, V.; Verhulst, N.; Cox, R.; Govaerts, B. Weed dynamics and conservation agriculture principles: A review. F. Crops Res. 2015, 183, 56–68. [Google Scholar] [CrossRef]
- Liebman, M.; Gallandt, E.R. Many Little Hammers: Ecological Management of Crop-Weed Interactions. In Ecology in Agriculture; Jackson, L.E., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 291–343. [Google Scholar]
- Bhatt, R. Zero tillage for mitigating global warming consequences and improving livelihoods in South Asia. In Environmental Sustainability and Climate Change Adaptation Strategies; Information Science Reference: Hershey, PA, USA, 2016; pp. 126–161. [Google Scholar] [CrossRef]
- Dierauer, H.; Hegglin, D.; Böhler, D. Direktsaat von Mais im Biolandbau; FiBL: Frick, Switzerland, 2015. [Google Scholar]
- Baraibar, B.; Hunter, M.C.; Schipanski, M.E.; Hamilton, A.; Mortensen, D.A. Weed Suppression in Cover Crop Monocultures and Mixtures. Weed Sci. 2018, 66, 121–133. [Google Scholar] [CrossRef]
- Drinkwater, L.E.; Wagoner, P.; Sarrantonio, M. Legume-based cropping systems have reduced carbon and nitrogen losses. Nature 1998, 396, 262–265. [Google Scholar] [CrossRef]
- Shilling, D.G.; Brecke, B.J.; Hiebsch, C.; MacDonald, G. Effect of Soybean (Glycine max) Cultivar, Tillage, and Rye (Secale cereale) Mulch on Sicklepod (Senna obtusifolia). Weed Technol. 1995, 9, 339–342. [Google Scholar] [CrossRef]
- Teasdale, J.R.; Beste, C.E.; Potts, W.E. Response of Weeds to Tillage and Cover Crop Residue. Weed Sci. 1991, 39, 195–199. [Google Scholar] [CrossRef]
- Mischler, R.; Duiker, S.W.; Curran, W.S.; Wilson, D. Hairy vetch management for no-till organic corn production. Agron. J. 2010, 102, 355–362. [Google Scholar] [CrossRef]
- Teasdale, J.R.; Mohler, C.L. The quantitative relationship between weed emergence and the physical properties of mulches. Weed Sci. 2000, 48, 385–392. [Google Scholar] [CrossRef]
- Ashford, D.L.; Reeves, D.W. Use of a mechanical roller-crimper as an alternative kill method for cover crops. Am. J. Altern. Agric. 2003, 18, 37–45. [Google Scholar] [CrossRef]
- Böhler, D.; Dierauer, H. Direktsaat von Mais in überwinternde Begrünungen unter Biobedingungen: Messerwalze statt Glyphosat. Landwirtschaft Ohne Pflug. 2017, 5, 39–43. [Google Scholar]
- Wells, M.S.; Reberg-Horton, S.C.; Smith, A.N.; Grossman, J.M. The Reduction of Plant-Available Nitrogen by Cover Crop Mulches and Subsequent Effects on Soybean Performance and Weed Interference. Agron. J. 2013, 105, 539–545. [Google Scholar] [CrossRef]
- Booth, B.D.; Swanton, C.J. Assembly theory applied to weed communities. Weed Sci. 2002, 50, 2–13. [Google Scholar] [CrossRef]
- de Mol, F.; von Redwitz, C.; Gerowitt, B. Weed species composition of maize fields in Germany is influenced by site and crop sequence. Weed Res. 2015, 55, 574–585. [Google Scholar] [CrossRef]
- von Redwitz, C.; Gerowitt, B. Maize-dominated crop sequences in northern Germany: Reaction of the weed species communities. Appl. Veg. Sci. 2018, 21, 431–441. [Google Scholar] [CrossRef]
- Mehrtens, J.; Schulte, M.; Hurle, K. Unkrautflora in Mais: Ergebnisse eines monitorings in Deutschland. Gesunde Pflanz. 2005, 57, 206–218. [Google Scholar] [CrossRef]
- Pannwitt, H.; Krato, C.; Gerowitt, B. Unkraut-Monitoring 2.0—Erste Ergebnisse zur aktuellen Unkrautvegetation im Mais. In Proceedings of the 28. Deutsche Arbeitsbesprechung über Fragen der Unkrautbiologie und -Bekämpfung, Braunschweig, Germany, 27 February–1 March 2018; pp. 24–29. [Google Scholar]
- Pannwitt, H.; Krato, C.; Gerowitt, B. Unkräuter im Mais—Veränderung der Eigenschaften der Unkrautzusammensetzung durch Bodenbearbeitung und Fruchtfolge. In Proceedings of the 29. Deutsche Arbeitsbesprechung über Fragen der Unkrautbiologie und -Bekämpfung, Braunschweig, Germany, 3–5 March 2020; pp. 186–191. [Google Scholar]
- Armengot, L.; Blanco-Moreno, J.M.; Bàrberi, P.; Bocci, G.; Carlesi, S.; Aendekerk, R.; Berner, A.; Celette, F.; Grosse, M.; Huiting, H.; et al. Tillage as a driver of change in weed communities: A functional perspective. Agric. Ecosyst. Environ. 2016, 222, 276–285. [Google Scholar] [CrossRef]
- Streit, B.; Rieger, S.B.; Stamp, P.; Richner, W. The effect of tillage intensity and time of herbicide application on weed communities and populations in maize in central Europe. Agric. Ecosyst. Environ. 2002, 92, 211–224. [Google Scholar] [CrossRef]
- Froud-Williams, R.J.; Chancellor, R.J.; Drennan, D.S.H. The Effects of Seed Burial and Soil Disturbance on Emergence and Survival of Arable Weeds in Relation to Minimal Cultivation. J. Appl. Ecol. 1984, 21, 629–641. [Google Scholar] [CrossRef]
- Egley, G.H.; Williams, R.D. Decline of Weed Seeds and Seedling Emergence over Five Years as Affected by Soil Disturbances. Weed Sci. 1990, 38, 504–510. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; Version 4.0.4; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R. Version 1.4.1106; RStudio: Boston, MA, USA, 2021. [Google Scholar]
- Simko, I.; Piepho, H.P. The area under the disease progress stairs: Calculation, advantage, and application. Phytopathology 2012, 102, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Jäger, E.J. Exkursionsflora von Deutschland 3; Elsevier GmbH.: München, Germany, 2007; ISBN 978-3-8274-1842-5. [Google Scholar]
- Schaarschmidt, F.; Vaas, L. Analysis of trials with complex treatment structure using multiple contrast tests. HortScience 2009, 44, 188–195. [Google Scholar] [CrossRef]
- Piepho, H.P.; Büchse, A.; Emrich, K. A Hitchhiker’s Guide to Mixed Models for Randomized Experiments. J. Agron. Crop Sci. 2003, 189, 310–322. [Google Scholar] [CrossRef]
- Bretz, F.; Hothorn, T.; Westfall, P. Multiple Comparisons Using R; CRC Press: New York, NY, USA, 2011; ISBN 9781584885740. [Google Scholar]
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. Version 1.3–5. 2021. Available online: https://cran.r-project.org/web//packages/agricolae/agricolae.pdf (accessed on 20 November 2022).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springler: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. Version 0.4.1. 2021. Available online: https://cran.microsoft.com/snapshot/2021-09-26/web/packages/DHARMa/vignettes/DHARMa.html (accessed on 20 November 2022).
- Lenth, R.V. emmeans: Estimated Marginal Means, Aka Least-Squares Means. Version 1.6.1. 2021. Available online: https://github.com/rvlenth/emmeans#readme (accessed on 20 November 2022).
- Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. Version 0.9–79. 2021. Available online: https://cran.uni-muenster.de/web/packages/RVAideMemoire/RVAideMemoire.pdf (accessed on 20 November 2022).
- Sierra, J. Temperature and soil moisture dependence of N mineralization in intact soil cores. Soil Biol. Biochem. 1997, 29, 1557–1563. [Google Scholar] [CrossRef]
- Schwartz, R.C.; Baumhardt, R.L.; Evett, S.R. Tillage effects on soil water redistribution and bare soil evaporation throughout a season. Soil Tillage Res. 2010, 110, 221–229. [Google Scholar] [CrossRef]
- Dahiya, R.; Ingwersen, J.; Streck, T. The effect of mulching and tillage on the water and temperature regimes of a loess soil: Experimental findings and modeling. Soil Tillage Res. 2007, 96, 52–63. [Google Scholar] [CrossRef]
- FAO. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements—FAO Irrigation and Drainage Paper 56; Allen, R.G., Pereira, L.S., Raes, D., Smith, M., Eds.; FAO: Rome, Italy, 1998; ISBN 92-5-104219-5. [Google Scholar]
- Teasdale, J.R.; Mohler, C.L. Light Transmittance, Soil Temperature, and Soil Moisture under Residue of Hairy Vetch and Rye. Agron. J. 1993, 85, 673–680. [Google Scholar] [CrossRef]
- Boscutti, F.; Sigura, M.; Gambon, N.; Lagazio, C.; Krüsi, B.O.; Bonfanti, P. Conservation Tillage Affects Species Composition But Not Species Diversity: A Comparative Study in Northern Italy. Environ. Manage. 2015, 55, 443–452. [Google Scholar] [CrossRef]
- Davis, A.S.; Dixon, P.M.; Liebman, M. Using matrix models to determine cropping system effects on annual weed demography. Ecol. Appl. 2004, 14, 655–668. [Google Scholar] [CrossRef]
- Wickham, H.; Bryan, J. Readxl: Read Excel Files. Version 1.3.1. 2019. Available online: https://mran.microsoft.com/web/packages/readxl/readxl.pdf (accessed on 20 November 2022).
- Wickham, H.; François, R.; Henry, L.; Müller, K. Dplyr: A Grammar of Data Manipulation. Version 1.0.6. 2021. Available online: https://mran.microsoft.com/web/packages/dplyr/dplyr.pdf (accessed on 20 November 2022).
- Firke, S. janitor: Simple Tools for Examining and Cleaning Dirty Data. Version 2.1.0. 2021. Available online: https://cran.r-project.org/web//packages/janitor/janitor.pdf (accessed on 20 November 2022).
- Kowarik, A.; Templ, M. Imputation with the R Package VIM. J. Stat. Softw. 2016, 74, 1–16. [Google Scholar] [CrossRef]
- Wickham, H. Reshaping data with the reshape package. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]
- Wright, K. Desplot: Plotting Field Plans for Agricultural Experiments. Version 1.8. 2020. Available online: https://cran.r-project.org/web/packages/desplot/desplot.pdf (accessed on 20 November 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springler: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Pedersen, T.L. Patchwork: The Composer of Plots. Version 1.1.1. 2020. Available online: https://cloud.r-project.org/web/packages/patchwork/patchwork.pdf (accessed on 20 November 2022).
- Schloerke, B.; Cook, D.; Larmarange, J.; Briatte, F.; Marbach, M.; Thoen, E.; Elberg, A.; Crowley, J. GGally: Extension to “ggplot2”. Version 2.1.1. 2021. Available online: https://mode.com/blog/r-ggplot-extension-packages/ (accessed on 20 November 2022).






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, F.; Böhm, H.; Graß, R.; Wachendorf, M.; Piepho, H.-P. Management Effect on the Weed Control Efficiency in Double Cropping Systems. Agronomy 2023, 13, 467. https://doi.org/10.3390/agronomy13020467
Schmidt F, Böhm H, Graß R, Wachendorf M, Piepho H-P. Management Effect on the Weed Control Efficiency in Double Cropping Systems. Agronomy. 2023; 13(2):467. https://doi.org/10.3390/agronomy13020467
Chicago/Turabian StyleSchmidt, Fruzsina, Herwart Böhm, Rüdiger Graß, Michael Wachendorf, and Hans-Peter Piepho. 2023. "Management Effect on the Weed Control Efficiency in Double Cropping Systems" Agronomy 13, no. 2: 467. https://doi.org/10.3390/agronomy13020467