Soil Microbes from Saline–Alkali Farmland Can Form Carbonate Precipitates

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
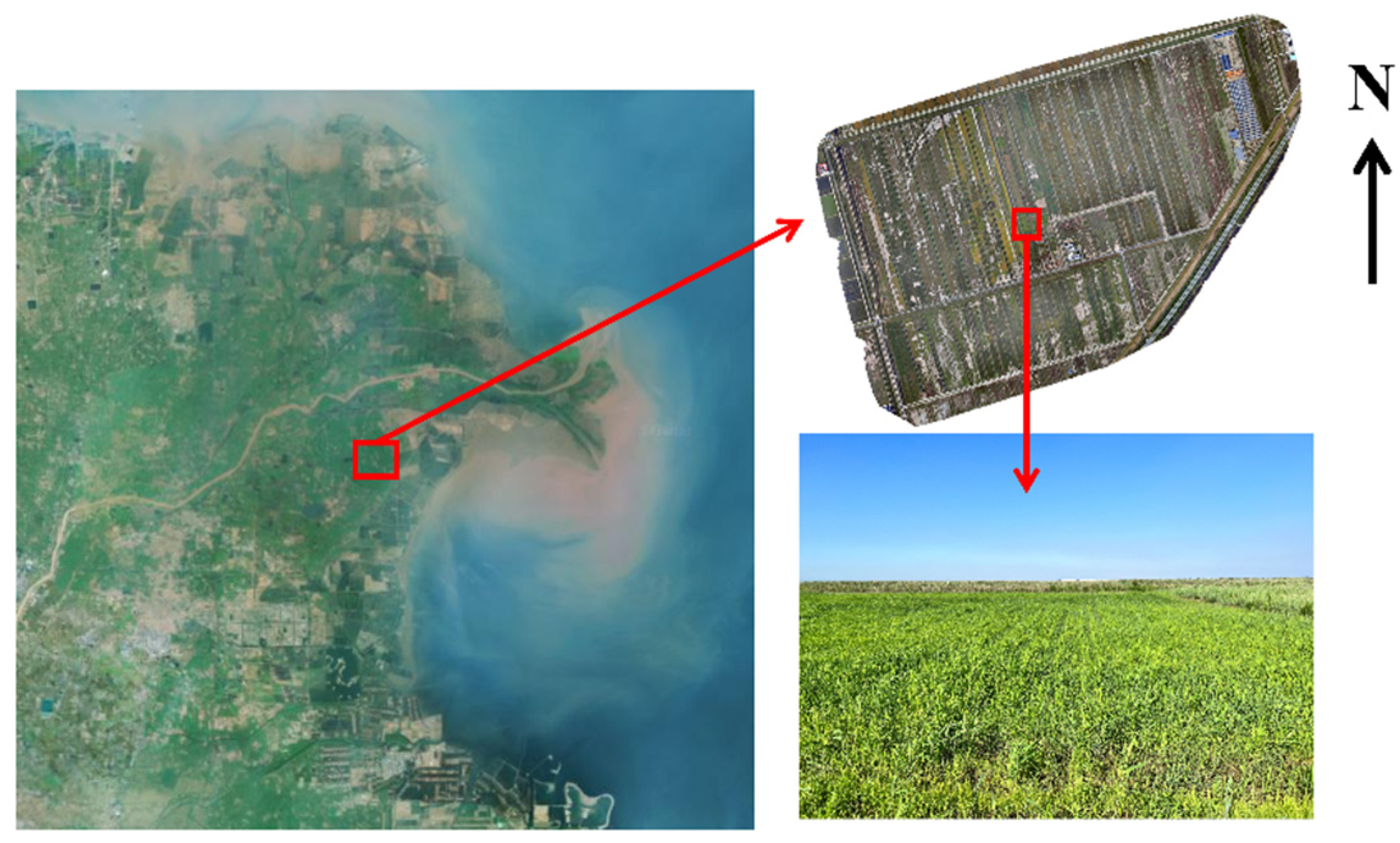
2.1. Site Description and Sampling Collection
2.2. Isolation and Identification of Saline–Alkali Soil Microbes
2.3. Processes of Microbial Carbonate Formation and Identification
2.4. Data Analysis and Processing
3. Results
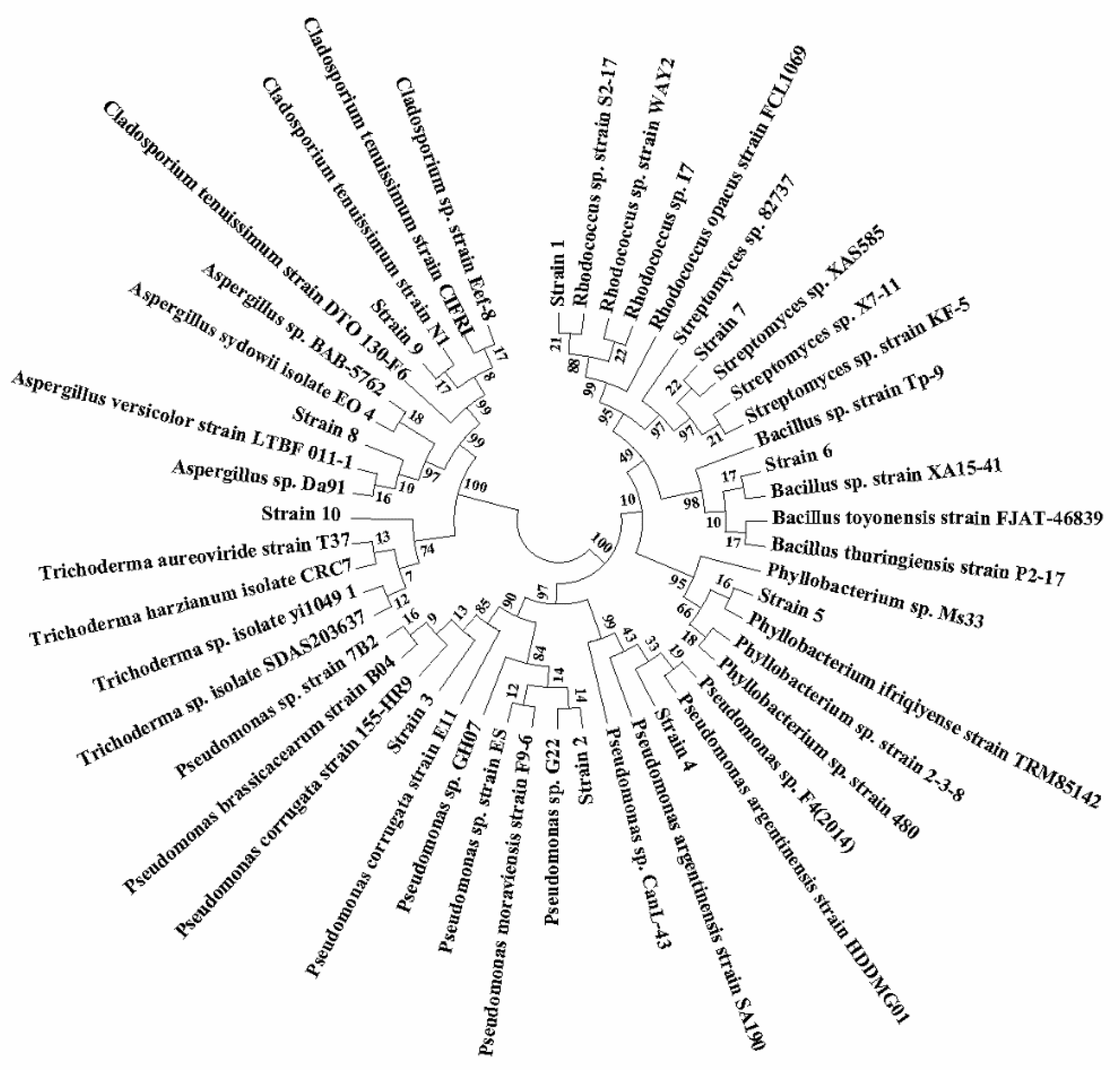
3.1. Isolation and Identification of the Microbial Strains
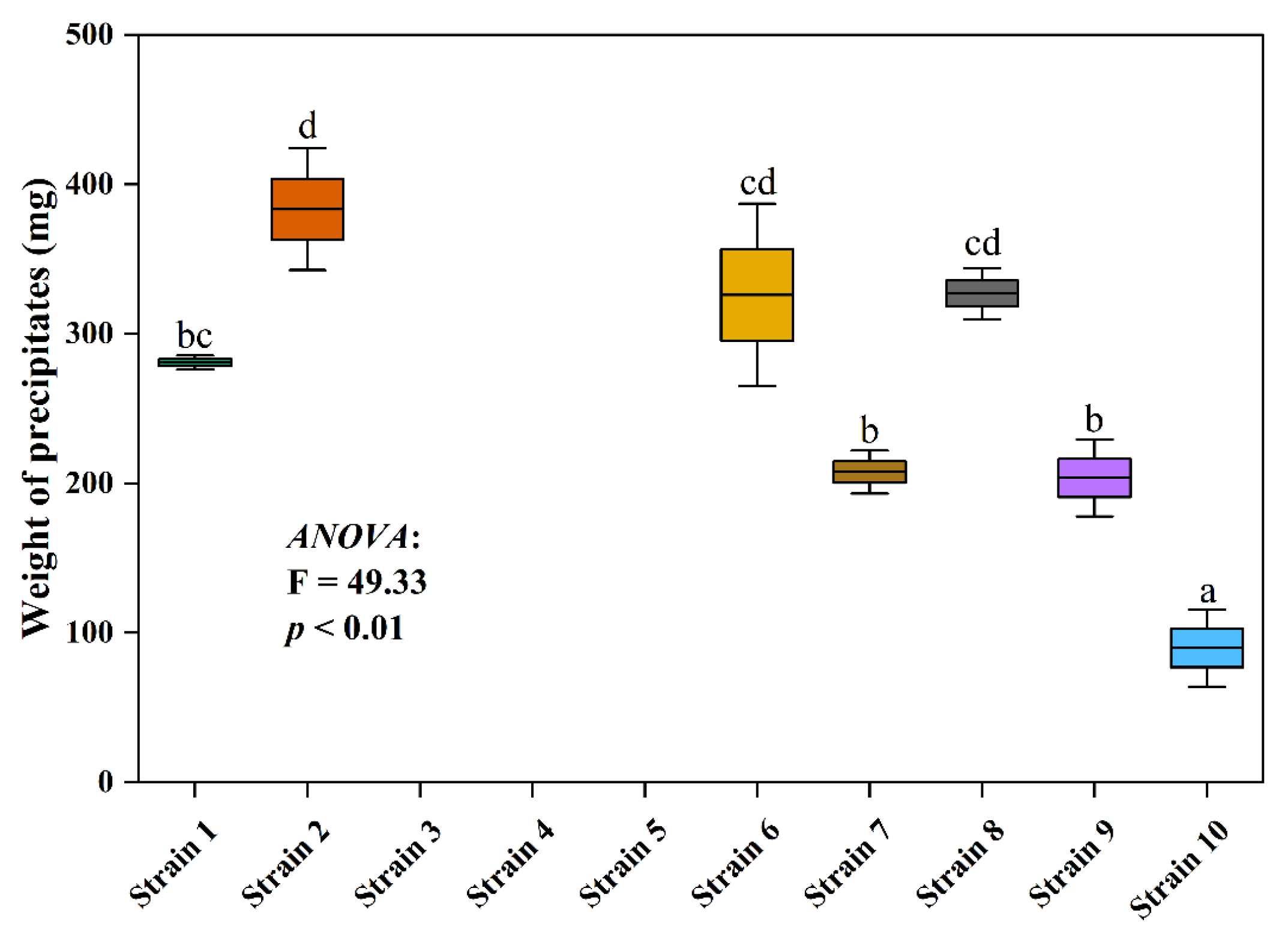
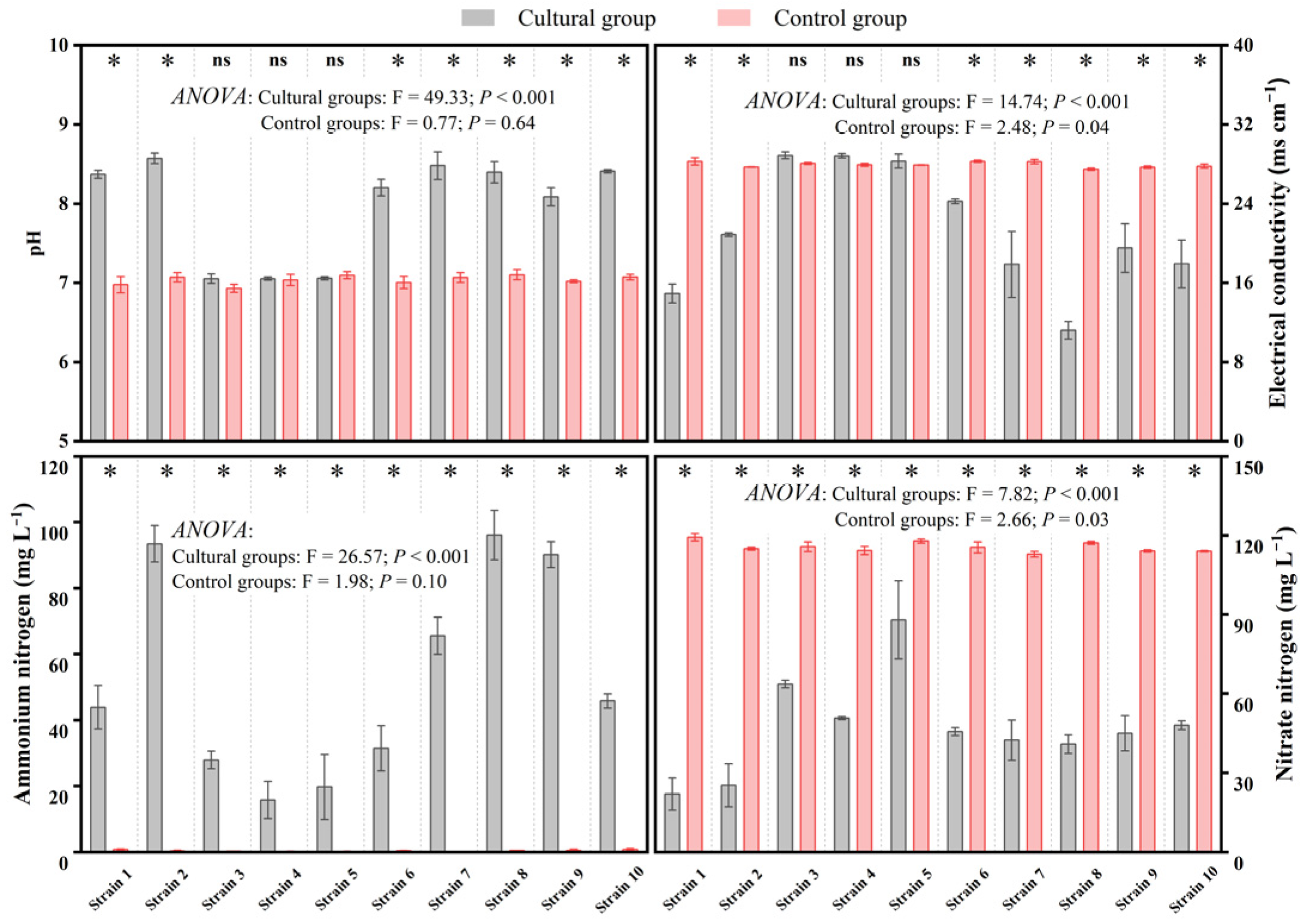
3.2. Precipitate Formation and Physicochemical Properties’ Variation in the Cultural Media
3.3. Identification of the Precipitates Formed by the Microbes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wicke, B.; Smeets, E.; Dornburg, V.; Vashev, B.; Gaiser, T.; Turkenburg, W.; Faaij, A. The global technical and economic potential of bioenergy from salt-affected soils. Energy Environ. Sci. 2011, 4, 2669–2681. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, J.; Xu, M.; Zhang, W.; Fan, T.; Zhang, J. Carbon accumulation in arid croplands of northwest China: Pedogenic carbonate exceeding organic carbon. Sci. Rep. 2015, 5, 11439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Wang, X.; Li, X.; Wang, J.; Xu, M.; Li, D. Dynamics of soil organic and inorganic carbon in the cropland of upper Yellow River Delta, China. Sci. Rep. 2016, 6, 36105. [Google Scholar] [CrossRef] [PubMed]
- Monger, H.C.; Kraimer, R.A.; Khresat, S.E.; Cole, D.R.; Wang, X.; Wang, J. Sequestration of inorganic carbon in soil and groundwater. Geology 2015, 43, 375–378. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Y.; Guo, C.; Xue, D.; Li, J.; Chen, Q.; Song, Z.; Lou, L.; Kuzyakov, Y.; Wang, Z.L.; et al. Conversion of coastal marshes to croplands decreases organic carbon but increases inorganic carbon in saline soils. Land Degrad. Dev. 2020, 31, 1099–1109. [Google Scholar] [CrossRef]
- Kim, J.H.; Jobbágy, E.G.; Richter, D.D.; Trumbore, S.E.; Jackson, R.B. Agricultural acceleration of soil carbonate weathering. Glob. Change Biol. 2020, 26, 5988–6002. [Google Scholar] [CrossRef]
- Hasinger, O.; Spangenberg, J.E.; Millière, L.; Bindschedler, S.; Cailleau, G.; Verrecchia, E.P. Carbon dioxide in scree slope deposits: A pathway from atmosphere to pedogenic carbonate. Geoderma 2015, 247, 129–139. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, Z.; Li, Y.; Kong, F.; Xi, M. Inorganic carbon sequestration and its mechanism of coastal saline-alkali wetlands in Jiaozhou Bay, China. Geoderma 2019, 351, 221–234. [Google Scholar] [CrossRef]
- Sanderman, J. Can management induced changes in the carbonate system drive soil carbon sequestration? A review with particular focus on Australia. Agric. Ecosyst. Environ. 2012, 155, 70–77. [Google Scholar] [CrossRef]
- Sánchez-Cañete, E.P.; Barron-Gafford, G.A.; Chorover, J. A considerable fraction of soil-respired CO2 is not emitted directly to the atmosphere. Sci. Rep. 2018, 8, 13518. [Google Scholar] [CrossRef] [PubMed]
- Boyce, C.K.; Ibarra, D.E.; D’Antonio, M.P. What we talk about when we talk about the long-term carbon cycle. New Phytol. 2022; advance online publication. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, Y.; Zhang, Y.; Qin, S.; Sun, Y.; Mao, H.; Miao, L. Desert soil sequesters atmospheric CO2 by microbial mineral formation. Geoderma 2020, 361, 114104. [Google Scholar] [CrossRef]
- Banks, E.D.; Taylor, N.M.; Gulley, J.; Lubbers, B.R.; Giarrizzo, J.G.; Bullen, H.A.; Hoehler, T.M.; Barton, H.A. Bacterial calcium carbonate precipitation in cave environments: A function of calcium homeostasis. Geomicrobiol. J. 2010, 27, 444–454. [Google Scholar] [CrossRef]
- Glunk, C.; Dupraz, C.; Braissant, O.; Gallagher, K.L.; Verrecchia, E.P.; Visscher, P.T. Microbially mediated carbonate precipitation in a hypersaline lake, Big Pond (Eleuthera, Bahamas). Sedimentology 2011, 58, 720–736. [Google Scholar] [CrossRef]
- Shi, S.; Tian, L.; Nasir, F.; Bahadur, A.; Batool, A.; Luo, S.; Yang, F.; Wang, Z.; Tian, C. Response of microbial communities and enzyme activities to amendments in saline-alkaline soils. Appl. Soil Ecol. 2019, 135, 16–24. [Google Scholar] [CrossRef]
- Yang, C.; Lv, D.; Jiang, S.; Lin, H.; Sun, J.; Li, K.; Sun, J. Soil salinity regulation of soil microbial carbon metabolic function in the Yellow River Delta, China. Sci. Total Environ. 2021, 790, 148258. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Y.; Fa, K.; Zhao, H.; Qin, S.; Yan, R.; Wu, B. Desert soil bacteria deposit atmospheric carbon dioxide in carbonate precipitates. Catena 2018, 170, 64–72. [Google Scholar] [CrossRef]
- Fang, H.; Liu, G.; Kearney, M. Georelational analysis of soil type, soil salt content, landform, and land use in the Yellow River Delta, China. Environ. Manag. 2005, 35, 72–83. [Google Scholar] [CrossRef]
- Bai, J.; Yu, Z.; Yu, L.; Wang, D.; Guan, Y.; Liu, X.; Gu, C.; Cui, B. In-situ organic phosphorus mineralization in sediments in coastal wetlands with different flooding periods in the Yellow River Delta, China. Sci. Total Environ. 2019, 682, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization (FAO); UNESCO. Soil Map of the World; FAO: Rome, Italy, 1978; Volume 1–10. [Google Scholar]
- Zhao, Q.Q.; Bai, J.H.; Zhang, G.L.; Jia, J.; Wang, W.; Wang, X. Effects of water and salinity regulation measures on soil carbon sequestration in coastal wetlands of the Yellow River Delta. Geoderma 2018, 319, 219–229. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. A Language and Environment for Statistical Computing; R Ver. 4.0. 3.; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org (accessed on 10 October 2022).
- Cao, C.; Jiang, J.; Sun, H.; Huang, Y.; Tao, F.; Lian, B. Carbonate mineral formation under the influence of limestone-colonizing actinobacteria: Morphology and polymorphism. Front. Microbiol. 2016, 7, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smythe, W.F.; McAllister, S.M.; Hager, K.W.; Hager, K.R.; Tebo, B.M.; Moyer, C.L. Silica biomineralization of Calothrix-dominated biofacies from Queen’s Laundry hot-spring, Yellowstone National Park, USA. Front. Environ. Sci. 2016, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Andrei, A.Ş.; Păuşan, M.R.; Tămaş, T.; Har, N.; Barbu-Tudoran, L.; Leopold, N.; Banciu, H.L. Diversity and biomineralization potential of the epilithic bacterial communities inhabiting the oldest public stone monument of Cluj-Napoca (Transylvania, Romania). Front. Microbiol. 2017, 8, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Kumar, A.; Zhang, Y. Microbial-induced carbonate precipitation prevents Cd2+ migration through the soil profile. Sci. Total Environ. 2022, 844, 157167. [Google Scholar] [CrossRef]
- Ronholm, J.; Schumann, D.; Sapers, H.M.; Izawa, M.; Applin, D.; Berg, B.; Mann, P.; Vali, H.; Flemming, R.L.; Cloutis, E.A.; et al. A mineralogical characterization of biogenic calcium carbonates precipitated by heterotrophic bacteria isolated from cryophilic polar regions. Geobiology 2014, 12, 542–556. [Google Scholar] [CrossRef]
- Balci, N.; Menekşe, M.; Karagüler, N.G.; Şeref Sönmez, M.; Meister, P. Reproducing authigenic carbonate precipitation in the hypersaline Lake Acıgöl (Turkey) with microbial cultures. Geomicrobiol. J. 2016, 33, 758–773. [Google Scholar] [CrossRef]
- Meier, A.; Kastner, A.; Harries, D.; Wierzbicka-Wieczorek, M.; Majzlan, J.; Büchel, G.; Kothe, E. Calcium carbonates: Induced biomineralization with controlled macromorphology. Biogeosciences 2017, 14, 4867–4878. [Google Scholar] [CrossRef] [Green Version]
- Montaño-Salazar, S.M.; Lizarazo-Marriaga, J.; Brandão, P.F.B. Isolation and potential biocementation of calcite precipitation inducing bacteria from Colombian buildings. Curr. Microbiol. 2018, 75, 256–265. [Google Scholar] [CrossRef]
- Okyay, T.O.; Nguyen, H.N.; Castro, S.L.; Rodrigues, D.F. CO2 sequestration by ureolytic microbial consortia through microbially-induced calcite precipitation. Sci. Total Environ. 2016, 572, 671–680. [Google Scholar] [CrossRef]
- Zhu, T.; Dittrich, M. Carbonate precipitation through microbial activities in natural environment, and their potential in biotechnology: A review. Front. Bioeng. Biotechnol. 2016, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Bindschedler, S.; Cailleau, G.; Verrecchia, E. Role of fungi in the biomineralization of calcite. Minerals 2016, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Uren, N.C. Calcium oxalate in soils, its origins and fate—A review. Soil Res. 2018, 56, 443–450. [Google Scholar] [CrossRef]
- Hervé, V.; Simon, A.; Randevoson, F.; Cailleau, G.; Rajoelison, G.; Razakamanarivo, H.; Bindschedler, S.; Verrecchia, E.; Junier, P. Functional Diversity of the Litter-Associated Fungi from an Oxalate-Carbonate Pathway Ecosystem in Madagascar. Microorganisms 2021, 9, 985. [Google Scholar] [CrossRef] [PubMed]
- Syed, S.; Buddolla, V.; Lian, B. Oxalate Carbonate Pathway-Conversion and Fixation of Soil Carbon-A Potential Scenario for Sustainability. Front. Plant Sci. 2020, 11, 591297. [Google Scholar] [CrossRef]
- Paiva, E.A.S. Are calcium oxalate crystals a dynamic calcium store in plants? New Phytol. 2019, 223, 1707–1711. [Google Scholar] [CrossRef] [Green Version]
- Tian, K.; Wu, Y.; Zhang, H.; Li, D.; Nie, K.; Zhang, S. Increasing wind erosion resistance of aeolian sandy soil by microbially induced calcium carbonate precipitation. Land Degrad. Dev. 2018, 29, 4271–4281. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weight of Precipitates | pH | EC | ||
|---|---|---|---|---|
| pH | 0.82 *** | |||
| EC | −0.63 *** | −0.81 *** | ||
| 0.69 *** | 0.70 *** | −0.66 *** | ||
| −0.68 *** | −0.72 *** | 0.59 *** | 0.55 ** |
| Strain Types | Element Composition | Crystal Types | ||||||
|---|---|---|---|---|---|---|---|---|
| Atom Percentage (%) | Quality Percentage (%) | Calcite | Vaterite | |||||
| Carbon | Oxygen | Calcium | Carbon | Oxygen | Calcium | % | % | |
| 1 | 25.52 | 59.64 | 14.83 | 16.52 | 51.43 | 32.05 | 18.90 | 81.10 |
| 2 | 26.54 | 61.61 | 11.84 | 17.92 | 55.41 | 26.67 | 46.70 | 53.30 |
| 6 | 25.87 | 60.84 | 13.29 | 17.11 | 53.58 | 29.31 | 100 | - |
| 7 | 23.17 | 60.60 | 16.24 | 14.66 | 51.07 | 34.28 | 10.70 | 89.30 |
| 8 | 19.53 | 63.26 | 17.21 | 12.12 | 52.27 | 35.62 | 55.90 | 44.10 |
| 9 | 26.80 | 58.90 | 14.30 | 17.52 | 51.29 | 31.19 | 42.70 | 57.30 |
| 10 | 23.88 | 61.17 | 14.96 | 15.38 | 52.47 | 32.15 | 100 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Li, J.; Zhang, Y.; Gong, H.; Hou, R.; Sun, Z.; Ouyang, Z. Soil Microbes from Saline–Alkali Farmland Can Form Carbonate Precipitates. Agronomy 2023, 13, 372. https://doi.org/10.3390/agronomy13020372
Liu Z, Li J, Zhang Y, Gong H, Hou R, Sun Z, Ouyang Z. Soil Microbes from Saline–Alkali Farmland Can Form Carbonate Precipitates. Agronomy. 2023; 13(2):372. https://doi.org/10.3390/agronomy13020372
Chicago/Turabian StyleLiu, Zhen, Jing Li, Yitao Zhang, Huarui Gong, Ruixing Hou, Zhigang Sun, and Zhu Ouyang. 2023. "Soil Microbes from Saline–Alkali Farmland Can Form Carbonate Precipitates" Agronomy 13, no. 2: 372. https://doi.org/10.3390/agronomy13020372