
The Yield-Forming Role of Nitrogen in Rice in the Growing Seasons with Variable Thermal Conditions
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Design
2.2. Field Experiment
2.3. Measurements and Data Analysis
2.4. Statistical Analysis
3. Results
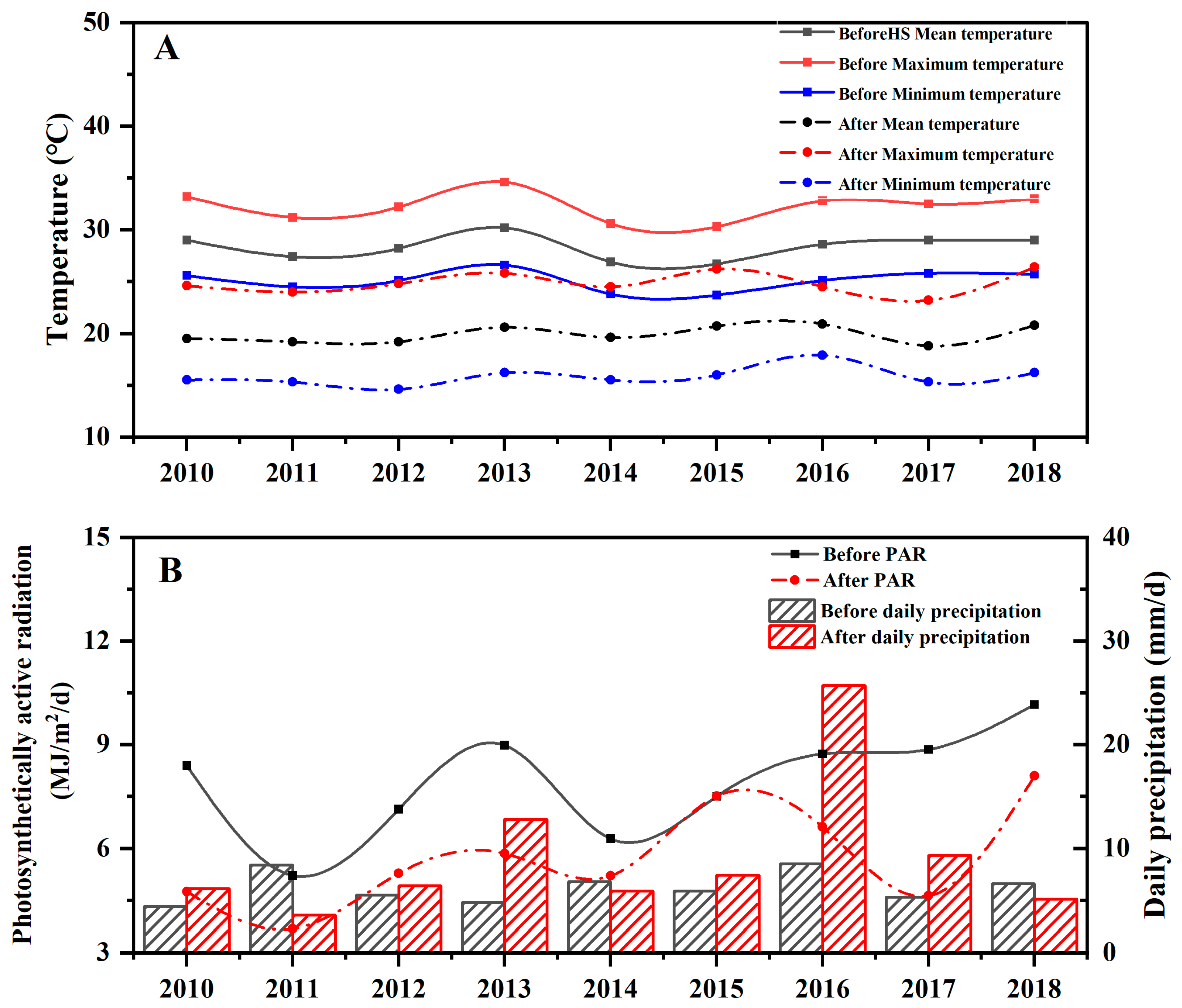
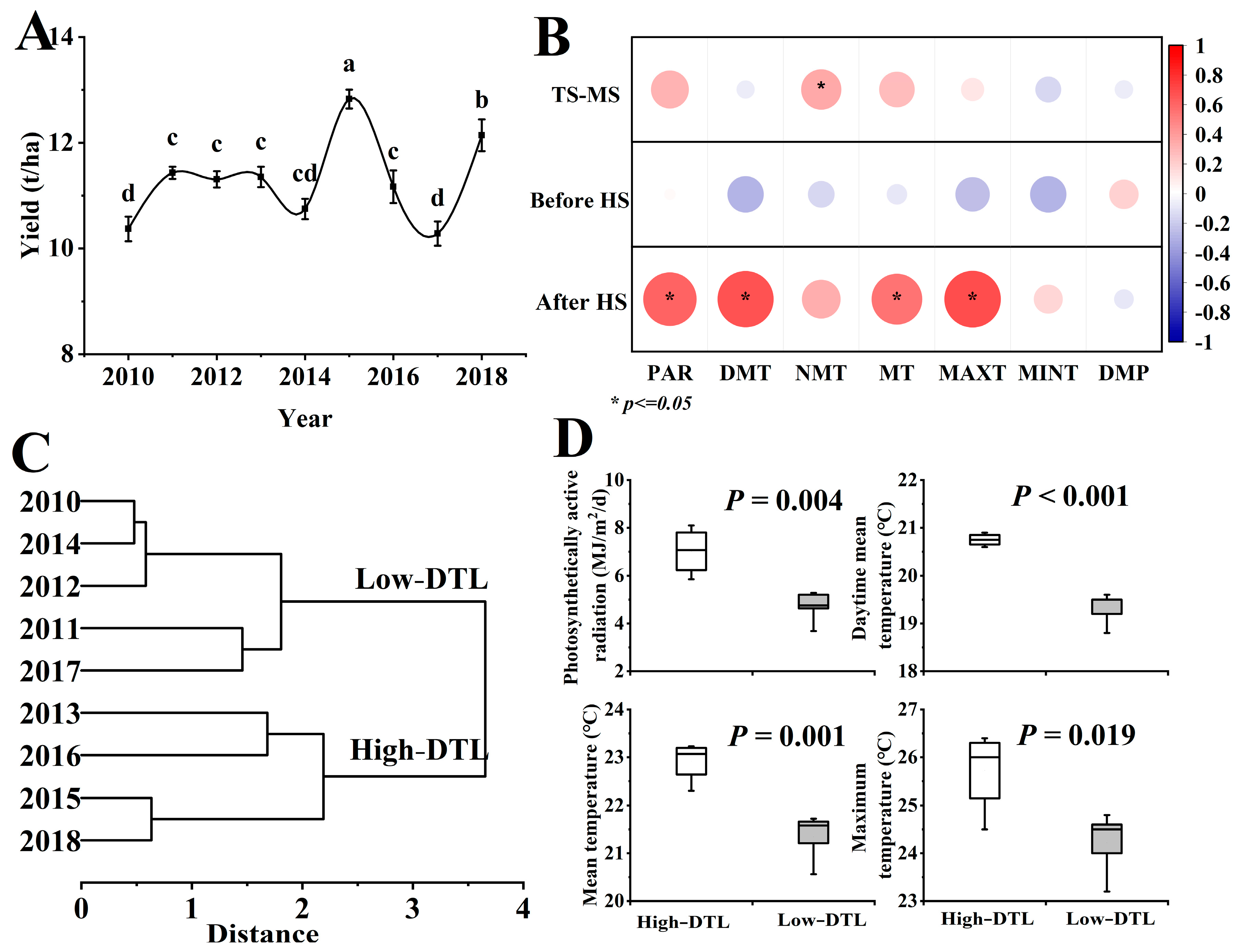
3.1. Relationship between Rice Yield and Meteorological Factors
3.2. Effect of Fertilization Management on Rice Yield and Yield Components under Two Weather Types
3.3. Effect of Fertilization Management on Biomass and HI under Two Weather Types
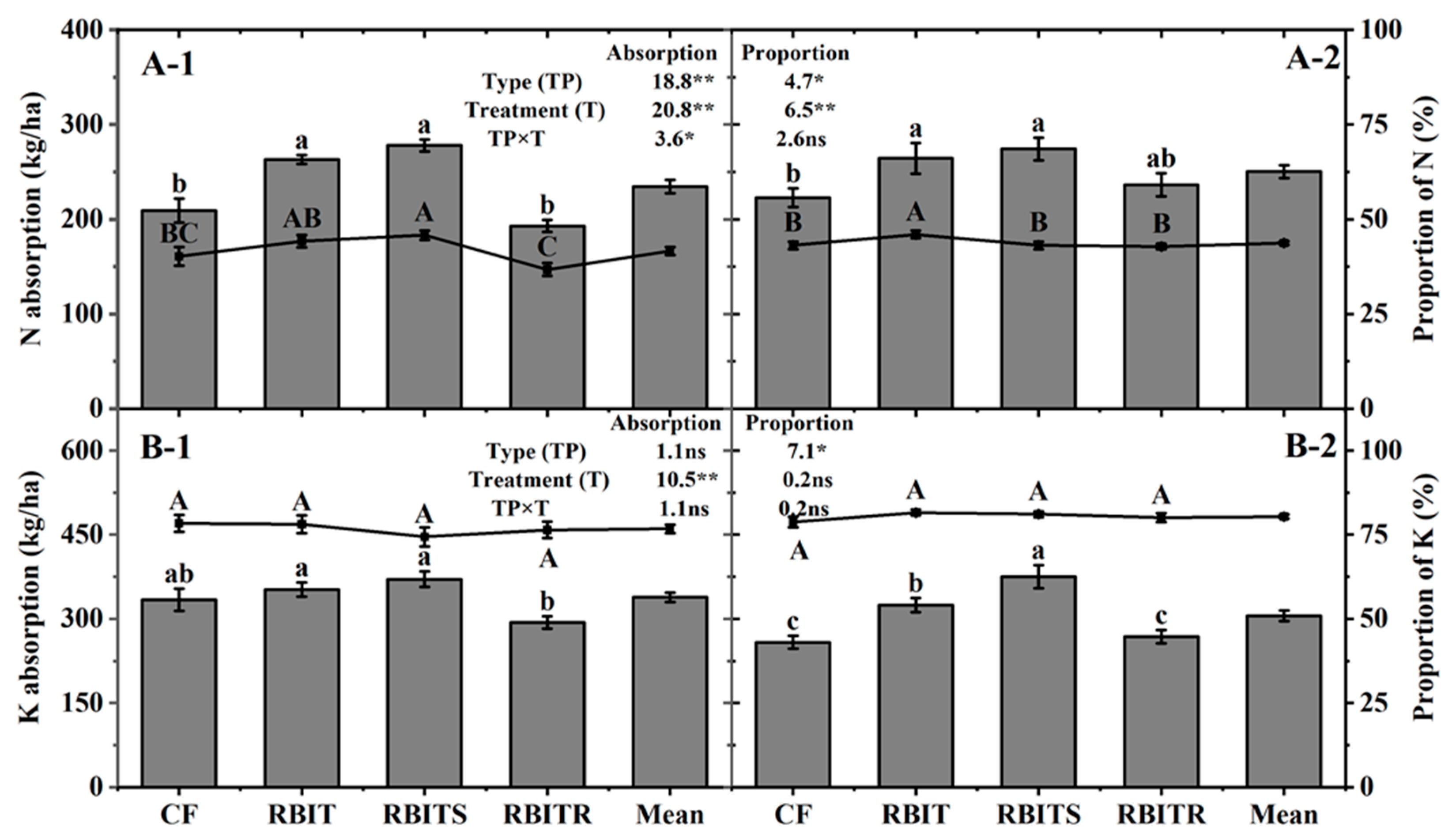
3.4. Effect of Fertilization Management on Nutrient Uptake under Two Weather Types
3.4.1. Absorption and Proportion of N and K in Rice
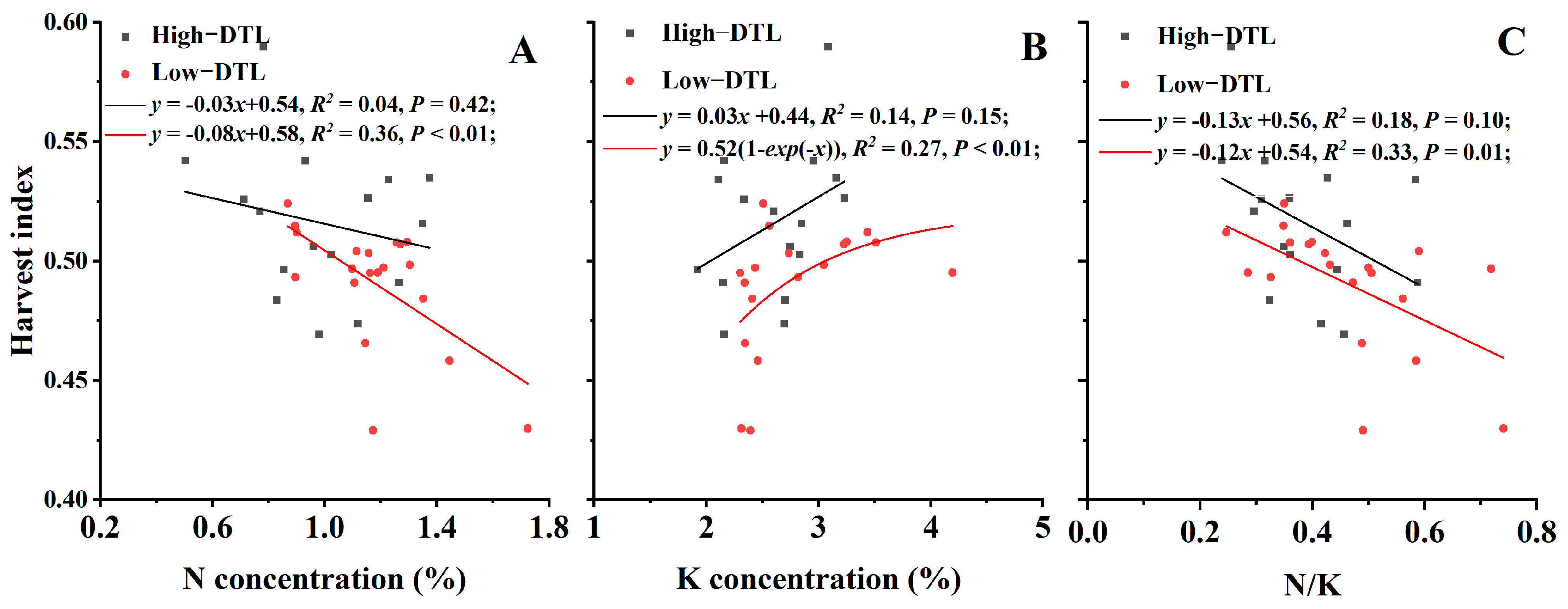
3.4.2. Nutrient Concentrations and N/K in Different Rice Parts
3.4.3. Regression Analysis
4. Discussion
4.1. Effect of Daytime Temperature and Light Change after Heading on Rice Production
4.2. Adaptation Strategies of Fertilization Management under Weather Changes
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- FAO. Food and Agriculture Organization of the United Nations Data. Statistical Databases 2022. Available online: http://www.fao.org/faostat/zh/#data/ (accessed on 30 August 2022).
- Zhang, Z.; Wang, P.; Chen, Y.; Song, X.; Wei, X.; Shi, P. Global warming over 1960–2009 did increase heat stress and reduce cold stress in the major rice-planting areas across China. Eur. J. Agron. 2014, 59, 49–56. [Google Scholar] [CrossRef]
- Espe, M.B.; Hill, J.E.; Hijmans, R.J.; McKenzie, K.; Mutters, R.; Espino, L.A.; Leinfelder-Miles, M.; Van Kessel, C.; Linquist, B.A. Point stresses during reproductive stage rather than warming seasonal temperature determine yield in temperate rice. Glob. Chang. Biol. 2017, 23, 4386–4395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tao, F.; Xiao, D.; Shi, W.; Liu, F.; Zhang, S.; Liu, Y.; Wang, M.; Bai, H. Contributions of climate, varieties, and agronomic management to rice yield change in the past three decades in China. Front. Earth Sci. 2015, 10, 315–327. [Google Scholar] [CrossRef]
- Ladha, J.K.; Dawe, D.; Pathak, H.; Padre, A.T.; Yadav, R.L.; Singh, B.; Hobbs, P.R. How extensive are yield declines in long-term rice-wheat experiments in Asia? Field Crops Res. 2003, 81, 159–180. [Google Scholar] [CrossRef]
- Ray, D.K.; Ramankutty, N.; Mueller, N.D.; West, P.; Foley, J.A. Recent patterns of crop yield growth and stagnation. Nat. Commun. 2012, 3, 1293. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2014 Synthesis Report; RK Pachauri, L.M., Ed.; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014. [Google Scholar]
- Stanhill, G.; Cohen, S. Global dimming: A review of the evidence for a widespread and significant reduction in global radiation with discussion of its probable causes and possible agricultural consequences. Agric. For. Meteorol. 2001, 107, 255–278. [Google Scholar] [CrossRef]
- Bai, H.; Tao, F.; Xiao, D.; Liu, F.; Zhang, H. Attribution of yield change for rice-wheat rotation system in China to climate change, cultivars and agronomic management in the past three decades. Clim. Chang. 2015, 135, 539–553. [Google Scholar] [CrossRef]
- Ye, C.; Ma, H.; Huang, X.; Xu, C.; Chen, S.; Chu, G.; Zhang, X.; Wang, D. Effects of increasing panicle-stage N on yield and N use efficiency of indica rice and its relationship with soil fertility. Crop J. 2022, 10, 1784–1797. [Google Scholar] [CrossRef]
- Kamiji, Y.; Yoshida, H.; Palta, J.A.; Sakuratani, T.; Shiraiwa, T. N applications that increase plant N during panicle development are highly effective in increasing spikelet number in rice. Field Crops Res. 2011, 122, 242–247. [Google Scholar] [CrossRef]
- Sui, B.; Feng, X.; Tian, G.; Hu, X.; Shen, Q.; Guo, S. Optimizing nitrogen supply increases rice yield and nitrogen use efficiency by regulating yield formation factors. Field Crops Res. 2013, 150, 99–107. [Google Scholar] [CrossRef]
- Zhang, Z.; Chu, G.; Liu, L.; Wang, Z.; Wang, X.; Zhang, H.; Yang, J.; Zhang, J. Mid-season nitrogen application strategies for rice varieties differing in panicle size. Field Crops Res. 2013, 150, 9–18. [Google Scholar] [CrossRef]
- Xia, L.; Lam, S.K.; Chen, D.; Wang, J.; Tang, Q.; Yan, X. Can knowledge-based N management produce more staple grain with lower greenhouse gas emission and reactive nitrogen pollution? A meta-analysis. Glob. Chang. Biol. 2017, 23, 1917–1925. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Clarke, V.C. The nitrogen cost of photosynthesis. J. Exp. Bot. 2019, 70, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Danso, I.; Webber, H.; Bourgault, M.; Ewert, F.; Naab, J.B.; Gaiser, T. Crop management adaptations to improve and stabilize crop yields under low-yielding conditions in the Sudan Savanna of West Africa. Eur. J. Agron. 2018, 101, 1–9. [Google Scholar] [CrossRef]
- Wang, L.; Deng, F.; Ren, W.-J. Shading tolerance in rice is related to better light harvesting and use efficiency and grain filling rate during grain filling period. Field Crops Res. 2015, 180, 54–62. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, J.; Qu, Z.; Zou, D.; Sha, H.; Liu, H.; Zhao, H. Effects of low water temperature during reproductive growth on photosynthetic production and nitrogen accumulation in rice. Field Crops Res. 2019, 242, 107587. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Liang, Y.; Han, Y.; Han, Y.; Tan, J. High Potassium Application Rate Increased Grain Yield of Shading-Stressed Winter Wheat by Improving Photosynthesis and Photosynthate Translocation. Front. Plant Sci. 2020, 11, 134. [Google Scholar] [CrossRef]
- Tsay, Y.-F.; Ho, C.-H.; Chen, H.-Y.; Lin, S.-H. Integration of Nitrogen and Potassium Signaling. Annu. Rev. Plant Biol. 2011, 62, 207–226. [Google Scholar] [CrossRef]
- Ou, X.; Cui, X.; Zhu, D.; Guo, L.; Liu, D.; Yang, Y. Lowering Nitrogen and Increasing Potassium Application Level Can Improve the Yield and Quality of Panax notoginseng. Front. Plant Sci. 2020, 11, 595095. [Google Scholar] [CrossRef]
- Weih, M.; Karlsson, P.S. Growth response of Mountain birch to air and soil temperature: Is increasing leaf-nitrogen content an acclimation to lower air temperature? New Phytol. 2001, 150, 147–155. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Wu, L.-M.; Ding, Y.-F.; Weng, F.; Wu, X.-R.; Li, G.-H.; Liu, Z.-H.; Tang, S.; Ding, C.-Q.; Wang, S.-H. Top-dressing nitrogen fertilizer rate contributes to decrease culm physical strength by reducing structural carbohydrate content in japonica rice. J. Integr. Agric. 2016, 15, 992–1004. [Google Scholar] [CrossRef] [Green Version]
- Webster, D.; Ebdon, J. Effects of Nitrogen and Potassium Fertilization on Perennial Ryegrass Cold Tolerance During Deacclimation in Late Winter and Early Spring. Hortscience 2005, 40, 842–849. [Google Scholar] [CrossRef]
- Cong, R.; Liu, T.; Lu, P.; Ren, T.; Li, X.; Lu, J. Nitrogen fertilization compensation the weak photosynthesis of Oilseed rape (Brassca napus L.) under haze weather. Sci. Rep. 2020, 10, 4047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qaswar, M.; Jing, H.; Ahmed, W.; Dongchu, L.; Shujun, L.; Lu, Z.; Cai, A.; Lisheng, L.; Yongmei, X.; Jusheng, G.; et al. Yield sustainability, soil organic carbon sequestration and nutrients balance under long-term combined application of manure and inorganic fertilizers in acidic paddy soil. Soil Tillage Res. 2020, 198, 104569. [Google Scholar] [CrossRef]
- Zhang, J.; Li, W.; Zhou, Y.; Ding, Y.; Xu, L.; Jiang, Y.; Li, G. Long-term straw incorporation increases rice yield stability under high fertilization level conditions in the rice–wheat system. Crop J. 2021, 9, 1191–1197. [Google Scholar] [CrossRef]
- Xu, J.; Han, H.; Ning, T.; Li, Z.; Lal, R. Long-term effects of tillage and straw management on soil organic carbon, crop yield, and yield stability in a wheat-maize system. Field Crops Res. 2019, 233, 33–40. [Google Scholar] [CrossRef]
- Sánchez, B.; Rasmussen, A.; Porter, J.R. Temperatures and the growth and development of maize and rice: A review. Glob. Chang. Biol. 2014, 20, 408–417. [Google Scholar] [CrossRef]
- Lodhi, N.; Kumar, N.; Singh, S.; Rani, D.S.; Panjwani, S.; Lata, S. Influence of Aerosols and Clouds on Surface Solar Radiation Effects on Potential Yields of Rice and Wheat in Indo-Gangetic Plains (Preprint). Social Science Research Network 2022. Available online: https://ssrn.com/abstract=4086681 (accessed on 20 October 2022).
- Cantagallo, J.; Medan, D.; Hall, A. Grain number in sunflower as affected by shading during floret growth, anthesis and grain setting. Field Crops Res. 2004, 85, 191–202. [Google Scholar] [CrossRef]
- Li, Q.; Deng, F.; Chen, H.; Zeng, Y.; Li, B.; Zhong, X.; Wang, L.; Zhou, W.; Chen, Y.; Ren, W. Shading decreases rice yield by impeding grain-filling progress after heading. Agron. J. 2020, 112, 4018–4030. [Google Scholar] [CrossRef]
- Wei, H.-Y.; Zhu, Y.; Qiu, S.; Han, C.; Hu, L.; Xu, D.; Zhou, N.-B.; Xing, Z.-P.; Hu, Y.-J.; Cui, P.-Y.; et al. Combined effect of shading time and nitrogen level on grain filling and grain quality in japonica super rice. J. Integr. Agric. 2018, 17, 2405–2417. [Google Scholar] [CrossRef]
- Cai, Z. Shade delayed flowering and decreased photosynthesis, growth and yield of Sacha Inchi (Plukenetia volubilis) plants. Ind. Crops Prod. 2011, 34, 1235–1237. [Google Scholar] [CrossRef]
- Shu, S.; Tang, Y.; Yuan, Y.; Sun, J.; Zhong, M.; Guo, S. The role of 24-epibrassinolide in the regulation of photosynthetic characteristics and nitrogen metabolism of tomato seedlings under a combined low temperature and weak light stress. Plant Physiol. Biochem. 2016, 107, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Liu, Z.; Zhao, B.; Liu, P.; Zhang, J.-W. Physiological and comparative proteomic analysis provides new insights into the effects of shade stress in maize (Zea mays L.). BMC Plant Biol. 2020, 20, 60. [Google Scholar] [CrossRef] [PubMed]
- Kyveryga, P.M.; Caragea, P.C.; Kaiser, M.S.; Blackmer, T.M. Predicting Risk from Reducing Nitrogen Fertilization Using Hierarchical Models and On-Farm Data. Agron. J. 2013, 105, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Hamoud, Y.A.; Shaghaleh, H.; Wang, R.; Gouertoumbo, W.F.; Hamad, A.A.A.; Sheteiwy, M.S.; Wang, Z.; Xiangping, G. Wheat Straw Burial Improves Physiological Traits, Yield and Grain Quality of Rice by Regulating Antioxidant System and Nitrogen Assimilation Enzymes under Alternate Wetting and Drying Irrigation. Rice Sci. 2022, 29, 473–488. [Google Scholar] [CrossRef]
- Cui, H.; Luo, Y.; Chen, J.; Jin, M.; Li, Y.; Wang, Z. Straw return strategies to improve soil properties and crop productivity in a winter wheat-summer maize cropping system. Eur. J. Agron. 2022, 133, 126436. [Google Scholar] [CrossRef]
- Ramteke, P.R.; Vashisht, B.B.; Sharma, S.; Jalota, S.K. Assessing and Ranking Influence of Rates of Rice (Oryza sativa L.) Straw Incorporation and N Fertilizer on Soil Physicochemical Properties and Wheat (Triticum aestivum L.) Yield in Rice-Wheat System. J. Soil. Sci. Plant Nut. 2022, 22, 515–526. [Google Scholar] [CrossRef]
- Hou, S.; Thiergart, T.; Vannier, N.; Mesny, F.; Ziegler, J.; Pickel, B.; Hacquard, S. A microbiota–root–shoot circuit favours Arabidopsis growth over defence under suboptimal light. Nat. Plants 2021, 7, 1078–1092. [Google Scholar] [CrossRef]
- Haro, R.; Benito, B. The Role of Soil Fungi in K+ Plant Nutrition. Int. J. Mol. Sci. 2019, 20, 3169. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Samo, N.; Zhao, C.; Wang, H.; Yang, G.; Hu, Y.; Peng, Y.; Rasul, F. Negative and Positive Impacts of Rape Straw Returning on the Roots Growth of Hybrid Rice in the Sichuan Basin Area. Agronomy 2019, 9, 690. [Google Scholar] [CrossRef]
- Hou, W.; Yan, J.; Jákli, B.; Lu, J.; Ren, T.; Cong, R.; Li, X. Synergistic Effects of Nitrogen and Potassium on Quantitative Limitations to Photosynthesis in Rice (Oryza sativa L.). J. Agric. Food Chem. 2018, 66, 5125–5132. [Google Scholar] [CrossRef] [PubMed]
- Tao, F.; Zhang, Z.; Shi, W.; Liu, Y.; Xiao, D.; Zhang, S.; Zhu, Z.; Wang, M.; Liu, F. Single rice growth period was prolonged by cultivars shifts, but yield was damaged by climate change during 1981–2009 in China, and late rice was just opposite. Glob. Chang. Biol. 2013, 19, 3200–3209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, Q. Identification of current research intensity and influence factors of agricultural nitrogen loss from cropping systems. J. Clean. Prod. 2020, 276, 123308. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fertilizer Period | CF | RBIT | RBITS | RBITR |
|---|---|---|---|---|
| Total nitrogen | 300 | 300 | 300 | 198 |
| Basal | 150 | 120 | 120 | 79.2 |
| Tilling | 75 | 60 | 60 | 39.6 |
| Panicle initiation | 75 | 60 | 60 | 39.6 |
| Spikelet differentiation | – | 60 | 60 | 39.6 |
| Total phosphorous (P2O5) | 150 | 150 | 150 | 120 |
| Basal | 150 | 75 | 75 | 60 |
| Jointing | – | 75 | 75 | 60 |
| Total potassium (K2O) | 240 | 240 | 240 | 180 |
| Basal | 240 | 120 | 120 | 90 |
| Jointing | – | 120 | 120 | 90 |
| Type | Treatment | Yield (t/ha) | No. of Panicles (/m2) | No. of Spikelets Per Panicle | Grain Weight (mg) | Seed Setting Rate (%) |
|---|---|---|---|---|---|---|
| High-DTL | CF | 11.6 ab | 337.7 ab | 123.4 c | 31.2 b | 90.0 a |
| RBIT | 12.4 a | 360.7 a | 130.9 b | 31.4 ab | 84.3 bc | |
| RBITS | 12.2 ab | 344.4 ab | 138.9 a | 31.2 b | 82.9 c | |
| RBITR | 11.4 b | 313.1 b | 126.8 bc | 32.6 a | 89.7 ab | |
| Mean | 11.9 | 339.6 | 130.1 | 31.6 | 86.6 | |
| Low-DTL | CF | 10.5 B | 319.2 A | 123.4 C | 29.9 AB | 89.9 A |
| RBIT | 10.8 B | 331.6 A | 129.8 BC | 29.3 B | 86.2 A | |
| RBITS | 11.4 A | 331.9 A | 138.5 A | 29.4 AB | 85.3 A | |
| RBITR | 10.6 B | 304.2 B | 130.8 B | 30.1 A | 88.7 A | |
| Mean | 10.5 | 321.7 | 130.6 | 29.7 | 87.5 | |
| Type (TP) | 59.2 ** | 9.9 ** | 0.0 ns | 66.3 ** | 1.0 ns | |
| Treatment (T) | 3.9 * | 3.9 * | 11.1 ** | 3.8 * | 3.0 * | |
| TP × T | 1.0 ns | 0.2 ns | 0.5 ns | 1.0 ns | 0.3 ns | |
| Type | Treatment | Biomass (t/ha) | Harvest Index | ||
|---|---|---|---|---|---|
| HS | HS–MS | MS | |||
| High-DTL | CF | 12.2 a | 8.0 ab | 20.7 a | 0.509 a |
| RBIT | 12.2 a | 9.2 a | 21.0 a | 0.515 a | |
| RBITS | 12.0 a | 9.1 a | 21.1 a | 0.509 a | |
| RBITR | 11.5 a | 7.2 b | 18.7 b | 0.534 a | |
| Mean | 12.0 | 8.4 | 20.4 | 0.517 | |
| Low-DTL | CF | 11.3 A | 7.3 B | 18.5 B | 0.489 A |
| RBIT | 10.4 A | 8.7 A | 19.1 B | 0.484 A | |
| RBITS | 11.2 A | 8.9 A | 20.4 A | 0.497 A | |
| RBITR | 10.6 A | 7.9 B | 18.1 B | 0.496 A | |
| Mean | 10.9 | 8.2 | 19.0 | 0.491 | |
| Type (TP) | 18.5 ** | 0.6 ns | 15.1 ** | 12.1 ** | |
| Treatment (T) | 1.5 ns | 8.8 ** | 7.7 ** | 1.1 ns | |
| TP × T | 0.8ns | 1.0 ns | 1.5 ns | 0.6 ns | |
| Type | Treatment | N Concentration (%) | K Concentration (%) | N/K | |||
|---|---|---|---|---|---|---|---|
| Stem + Leaf | Panicle | Stem + Leaf | Panicle | Stem + Leaf | Panicle | ||
| High-DTL | CF | 0.81 a | 1.18 c | 2.47a | 0.64 a | 0.33b | 2.18 a |
| RBIT | 1.17 a | 1.36 a | 2.70 a | 0.71 a | 0.45 a | 2.20 a | |
| RBITS | 1.19 a | 1.41 a | 2.75 a | 0.73 a | 0.45 a | 2.24 a | |
| RBITR | 0.78 a | 1.28 b | 2.49a | 0.69 a | 0.33 b | 2.06 a | |
| Mean | 0.97 | 1.31 | 2.60 | 0.69 | 0.39 | 2.17 | |
| Low-DTL | CF | 1.08 B | 1.34 A | 2.30 C | 0.56 AB | 0.49 A | 2.49 A |
| RBIT | 1.27 A | 1.43 A | 2.83 B | 0.61 AB | 0.47 A | 2.43 A | |
| RBITS | 1.18 A | 1.47 A | 3.13 A | 0.67 A | 0.40 A | 2.37 A | |
| RBITR | 1.12 B | 1.40 A | 2.43 BC | 0.55 B | 0.47 A | 2.74 A | |
| Mean | 1.16 | 1.41 | 2.65 | 0.60 | 0.46 | 2.50 | |
| Type (TP) | 50.7 ** | 27.1 ** | 0.9 ns | 7.4 ** | 9.2 ** | 9.4 ** | |
| Treatment (T) | 27.1 ** | 4.9 ** | 4.2 ** | 0.8 ns | 1.3 ns | 0.1 ns | |
| TP × T | 5.9 ** | 0.8 ns | 1.7 ns | 0.2 ns | 3.8 * | 0.7 ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zhou, Y.; Wu, L.; Xu, L.; Xu, C.; Liang, D.; Ding, Y.; Zhang, Y.; Wang, J.; Li, G. The Yield-Forming Role of Nitrogen in Rice in the Growing Seasons with Variable Thermal Conditions. Agronomy 2023, 13, 313. https://doi.org/10.3390/agronomy13020313
Zhang J, Zhou Y, Wu L, Xu L, Xu C, Liang D, Ding Y, Zhang Y, Wang J, Li G. The Yield-Forming Role of Nitrogen in Rice in the Growing Seasons with Variable Thermal Conditions. Agronomy. 2023; 13(2):313. https://doi.org/10.3390/agronomy13020313
Chicago/Turabian StyleZhang, Jianwei, Yan Zhou, Longmei Wu, Lei Xu, Cong Xu, Dong Liang, Yanfeng Ding, Yongchun Zhang, Jidong Wang, and Ganghua Li. 2023. "The Yield-Forming Role of Nitrogen in Rice in the Growing Seasons with Variable Thermal Conditions" Agronomy 13, no. 2: 313. https://doi.org/10.3390/agronomy13020313