
Seed Priming Applied to Onion-Like Crops: State of the Art and Open Questions
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Seed Priming: Basic Principles and Technology
3. Seed Priming Applied to the Genus Allium
3.1. Onion
3.2. Leek
3.3. Welsh Onion
3.4. Seed Priming in Onion and Onion-Like Crops: Major Limitations and Lessons Learned
4. Molecular Hallmarks: From Model Plants to Allium Crops
5. Commercial Use of Seed Priming
Challenges, Open Questions, and Perspectives
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Sharifi-Rad, J.; Mnayer, D.; Tabanelli, G.; Stojanović-Radić, Z.; Sharifi-Rad, M.; Yousaf, Z.; Vallone, L.; Setzer, W.; Iriti, M. Plants of the genus Allium as antibacterial agents: From tradition to pharmacy. Cell. Mol. Biol. 2016, 62, 57–68. [Google Scholar]
- Food and Agriculture Organisation of the United Nations (FAO). Crops and Livestock products Countries—Select All; Regions—World + (Total); Elements—Production Quantity; Items—Onions and Shallots, Dry (Excluding Dehydrated)—Onions and Shallots, Green; Years. 2020. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 19 December 2022).
- Food and Agriculture Organization Corporate Statistical Database (FAOSTAT). Countries—Select All; Regions—World + (Total); Elements—Production Quantity; Items—Garlic; Years. 2020. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 19 December 2022).
- Padula, G.; Xia, X.Z.; Hołubowicz, R. Welsh onion (Allium fistulosum L.) seed physiology, breeding, production and trade. Plants 2022, 11, 343. [Google Scholar] [CrossRef]
- Hołubowicz, R. Seed Production and Technology; Wyd. UP w Poznaniu: Poznań, Poland, 2016; pp. 55–57. [Google Scholar]
- Morozowska, M.; Hołubowicz, R. Effect of bulb size on selected morphological characteristics of seed stalks, seed yield and quality of onion (Allium cepa L.) seeds. Folia Hortic. 2009, 21, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Bocian, S.; Hołubowicz, R. Effect of different ways of priming tomato (Lycopersicon esculentum Mill.) seeds on their quality. Pol. J. Nat. Sci. 2008, 23, 729–739. [Google Scholar] [CrossRef]
- Hołubowicz, R. Some Little Known Vegetable Crops from China for Growing in Poland; Wyd. UP w Poznaniu: Poznań, Poland, 2019; pp. 13–21. (In Polish) [Google Scholar]
- FAO. Seed and Seed Quality: Technical Information for FAO Emergency Staff; FAO Seed and Plant Genetic Resources Service: Rome, Italy, 2006. [Google Scholar]
- Khan, M.; Javed Iqbal, M.; Abbas, M.; Raza, H.; Waseem, R.; Arshad, A. Loss of vigour and viability in aged onion (Allium cepa L.) seeds. Int. J. Agric. Biol. 2004, 6, 708–771. [Google Scholar]
- Borowski, E.; Michałek, S. The effect of seed conditioning on the emergence and early growth of onion and carrot seedlings. Ann. Univ. Mariae Curie-Skłodowska Setio EEE 2006, 16, 119–129. [Google Scholar]
- Brar, N.S.; Kaushik, P.; Dudi, B.S. Assessment of natural ageing related physio-biochemical changes in onion seed. Agriculture 2019, 9, 163. [Google Scholar] [CrossRef] [Green Version]
- Hourston, J.E.; Pérez, M.; Gawthrop, F.; Richards, M.; Steinbrecher, T.; Leubner-Metzger, G. The effects of high oxygen partial pressure on vegetable Allium seeds with a short shelf-life. Planta 2020, 251, 105. [Google Scholar] [CrossRef]
- Heydecker, W.; Gibbins, B.M. The ‘priming’ of seeds. Acta Hortic. 1978, 83, 213–215. [Google Scholar] [CrossRef]
- Heydecker, W.; Higgins, J.; Gulliver, R.L. Accelerated germination by osmotic seed treatment. Nature 1973, 246, 42–44. [Google Scholar] [CrossRef]
- Paparella, S.; Araujo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef]
- Lutts, S.; Benincasa, P.; Wojtyla, L.; Kubala, S.; Pace, R.; Lechowska, K.; Quinet, M.; Garnczarska, M. Seed priming: New comprehensive approaches for an old empirical technique. In New Challenges in Seed Biology-Basic and Translational Research Driving Seed Technology; Intech Open: Rijeka, Croatia, 2016; pp. 1–46. [Google Scholar]
- Paul, S.; Dey, S.; Kundu, R. Seed priming: An emerging tool towards sustainable agriculture. Plant Growth Regul. 2022, 97, 215–234. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Bray, C.M.; West, C.E. Seeds and the art of genome maintenance. Front. Plant Sci. 2019, 10, 706. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.; Pathan, A.K.; Gothkar, P.; Joshi, A.; Chivasa, W.; Nyamudeza, P. On-farm seed priming: Using participatory methods to revive and refine a key technology. Agric. Syst. 2021, 69, 151–164. [Google Scholar] [CrossRef]
- Bray, C.M. Biochemical processes during the osmopriming of seeds. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Dekker, M. Inc.: New York, NY, USA, 1995; pp. 767–789. [Google Scholar]
- Farooq, M.; Usman, M.; Nadeem, F.; Rehman, H.; Wahid, A.; Basra, S.M.A.; Siddique, K.H.M. Seed priming in field crops: Potential benefits, adoption and challenges. Crop Pasture Sci. 2019, 70, 731–771. [Google Scholar] [CrossRef]
- Madsen, M.D.; Svejcar, L.; Radke, J.; Hulet, A. Inducing rapid seed germination of native cool season grasses with solid matrix priming and seed extrusion technology. PLoS ONE 2018, 13, e0204380. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, D.; Singh, S.; Parihar, M.; Rakshit, A. Seed bio-priming with microbial inoculants: A tailored approach towards improved crop performance, nutritional security, and agricultural sustainability for smallholder farmers. Curr. Res. Environ. Sustain. 2021, 3, 100093. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Imran, S.; Rauf, F.; Khatun, M.; Baskin, C.C.; Murata, Y.; Hasanuzzaman, M. Seed priming with phytohormones: An effective approach for the mitigation of abiotic stress. Plants 2021, 10, 37. [Google Scholar] [CrossRef]
- Mahakham, W.; Sarmah, A.K.; Maensiri, S.; Theerakulpisut, P. Nanopriming technology for enhancing germination and starch metabolism of aged rice seeds using phytosynthesized silver nanoparticles. Sci. Rep. 2017, 7, 8263. [Google Scholar] [CrossRef] [Green Version]
- El-Badri, A.M.A.; Batool, M.; Mohamed, I.A.A.; Khatab, A.; Sherif, A.; Wang, Z.K.; Salah, A.; Nishawy, E.; Ayaad, M.; Kuai, J.; et al. Modulation of salinity impact on early seedling stage via nano-priming application of zinc oxide on rape seed (Brassica napus L.). Plant Physiol. Biochem. 2021, 166, 376–392. [Google Scholar] [CrossRef]
- Shelar, A.; Singh, A.V.; Maharjan, R.S.; Laux, P.; Luch, A.; Gemmati, D.; Tisato, V.; Singh, S.P.; Santilli, M.F.; Shelar, A.; et al. Sustainable Agriculture through Multidisciplinary Seed Nanopriming: Prospects of Opportunities and Challenges. Cells 2021, 10, 2428. [Google Scholar] [CrossRef]
- Amritha, M.S.; Sridharan, K.; Puthur, J.T.; Dhankher, O.M. Priming with nanoscale materials for boosting abiotic stress tolerance in crop plants. J. Agric. Food Chem. 2021, 69, 10017–10035. [Google Scholar]
- Araújo, S.S.; Paparella, S.; Dondi, D.; Bentivoglio, A.; Carbonera, D.; Balestrazzi, A. Physical methods for seed invigoration: Advantages and challenges in seed technology. Front. Plant Sci. 2016, 7, 646. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.H.; Butcher, P.D. The effects of priming and ‘natural’ differences in quality amongst onion seed lots on the response of the rate of germination to temperature and the identification of the characteristics under genotypic control. J. Exp. Bot. 1988, 39, 935–950. [Google Scholar] [CrossRef]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Murray, G.A.; Swensen, J.B.; Beaver, G. Emergence of spring- and summer-planted onions following osmotic priming. Hortscience 1992, 27, 409–410. [Google Scholar] [CrossRef] [Green Version]
- Bujalski, W.; Nienow, A.W. Large-scale osmotic priming of onion seeds: A comparison of different strategies for oxygenation. Sci. Hortic. 1991, 46, 13–24. [Google Scholar] [CrossRef]
- Edelstein, M.; Welbaum, G.E. Seed O2 uptake and germination of cold-tolerant and cold-intolerant cultivars of muskmelon. Crop Sci. 2011, 51, 810–817. [Google Scholar] [CrossRef]
- Horita, M.; Saruyama, H. Acceleration of germination of onion seeds by priming treatment with trehalose and raffinose. Hortic. Res. 2006, 5, 75–78. [Google Scholar] [CrossRef]
- Fu, Y.; Zhang, Z.; Liu, J.; Chen, M.; Pan, R.; Hu, W.; Guan, Y.; Hu, J. Seed priming with spermidine and trehalose enhances chilling tolerance of rice via different mechanisms. J. Plant Regul. 2020, 39, 669–679. [Google Scholar] [CrossRef]
- Saranya, N.; Renugadevi, J.; Raja, K.; Rajashree, V.; Hemalatha, G. Seed priming studies for vigour enhancement in onion CO onion (5). J. Pharm. Phytochem. 2017, 6, 77–82. [Google Scholar]
- Veena, M.; Puthur, J.T. Seed nutripriming with zinc is an apt tool to alleviate malnutrition. Environ. Geochem. Health 2022, 44, 2355–2373. [Google Scholar] [CrossRef]
- Wang, Y.; Diao, P.; Kong, L.; Yu, R.; Zhang, M.; Zuo, T.; Fan, Y.; Niu, Y.; Yan, F.; Wuriyanghan, H. Ethylene enhances seed germination and seedling growth under salinity by reducing oxidative stress and promoting chlorophyll content via ETR2 pathway. Front. Plant Sci. 2020, 11, 1066. [Google Scholar] [CrossRef]
- Thejeshwini, B.; Manohar Rao, A.; Hanuman Nayak, M.; Sultana, R. Effect of seed priming on plant growth and bulb yield in onion (Allium cepa L.). Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 1242–1249. [Google Scholar] [CrossRef]
- Tsegay, B.A.; Andargie, M. Seed priming with gibberellic acid (GA3) alleviates salinity induced inhibition of germination and seedling growth of Zea mays L., Pisum sativum Var. abyssinicum A. Braun and Lathyrus sativus L. J. Crop Sci. Biotechnol. 2018, 21, 261–267. [Google Scholar] [CrossRef]
- Khan, M.N.; Khan, Z.; Luo, T.; Liu, J.; Rizwan, M.; Zhang, J.; Xu, Z.; Wu, H.; Liyong Hu, L. Seed priming with gibberellic acid and melatonin in rapeseed: Consequences for improving yield and seed quality under drought and non-stress conditions. Ind. Crops Prod. 2020, 156, 112850. [Google Scholar] [CrossRef]
- Caseiro, R.; Bennet, M.A.; Marcos-Filho, J. Comparison of three priming techniques for onion seed lots differing in initial seed quality. Seed Sci. Technol. 2004, 32, 365–375. [Google Scholar] [CrossRef]
- Da Silva, J.E.S.B.; de Paiva, E.P.; Leite, M.d.S.; Torres, S.B.; de Souza Neta, M.L.; Guirra, K.S. Salycilic acid in the physiological priming of onion seeds subjected to water and salt stress. Rev. Bras. Eng. Agric. Amb. 2019, 23, 219–224. [Google Scholar] [CrossRef]
- Dearman, J.; Brocklehurst, P.A.; Drew, R.L.K. Effects of osmotic priming and ageing on onion seed germination. Ann. Bot. 1986, 108, 639–648. [Google Scholar] [CrossRef]
- Basra, A.S.; Singh, B.; Malik, C.P. Amelioration of the effects of ageing in onion seeds by osmotic priming and associated changes in oxidative metabolism. Biol. Plant. 1994, 36, 365–371. [Google Scholar] [CrossRef]
- Brar, N.S.; Kaushik, P.; Dudi, B. Effect of seed priming treatment on the physiological quality of naturally aged onion (Allium cepa L.) seeds. Appl. Ecol. Environ. Res. 2019, 18, 849–862. [Google Scholar] [CrossRef]
- Muruli, C.N.; Bhanuprakash, K.; Channakeshava, B.C. Impact of seed priming on vigour in onion (Allium cepa L.) seeds. J. Appl. Hortic. 2016, 18, 68–70. [Google Scholar] [CrossRef]
- Acharya, P.; Jayaprakasha, G.K.; Crosby, K.M.; Jifon, J.L.; Patil, B.S. Green-synthesized nanoparticles enhanced seedling growth, yield, and quality of onion (Allium cepa L.). ACS Sustain. Chem. Eng. 2019, 7, 14580–14590. [Google Scholar] [CrossRef]
- Morillo-Coronado, A.C.; Martinez-Anzola, H.G.; Velandia-Diaz, J.D.; Morillo-Coronado, Y. Effects of static magnetic fields on onion (Allium cepa L.) seed germination and early seedling growth. Rev. Ciencia Agric. 2022, 39, 30–41. [Google Scholar] [CrossRef]
- Kubisz, L.; Holubowicz, R.; Gauza, M.; Li, H.; Hojan-Jezierska, D.; Jaroszyk, F. Effect of low frequency magnetic field on germination of onion (Allium cepa L.) seeds. Acta Phys. Pol. 2012, 121, A49–A53. [Google Scholar] [CrossRef]
- Hołubowicz, R.; Kubisz, L.; Gauza, M.; Tong, Y.; Hojan-Jezierska, D. Effecxt of low frequency magnetic field (LFMF) on the germination of seeds and selected useful characteristics of onion (Allium cepa L.). Not. Bot. Hortic. Agrobot. 2014, 42, 168–172. [Google Scholar]
- Ren, F.; Zhou, S. Phenolic components and health beneficial properties of onions. Agriculture 2021, 11, 872. [Google Scholar] [CrossRef]
- Davison, P.; Bray, C. Protein synthesis during osmopriming of leek (Allium porrum L.) seeds. Seed Sci. Res. 1991, 1, 29–35. [Google Scholar] [CrossRef]
- Clarke, N.A.; James, P.E. The effects of priming and accelerated ageing upon the nucleic acid content of leek seeds and their embryos. J. Exp. Bot. 1991, 42, 261–268. [Google Scholar] [CrossRef]
- Brocklehurst, P.A.; Dearman, J.; Drew, R.L.K. Effects of osmotic priming on seed germination and seedling growth in leek. Sci. Hortic. 1984, 24, 201–210. [Google Scholar] [CrossRef]
- Parera, C.A.; Cantliffe, D.J. Priming leek seed for improved germination and emergence at high temperature. Hortscience 1992, 27, 1077–1079. [Google Scholar] [CrossRef] [Green Version]
- Corbineau, F.; Picards, M.A.; Côme, D. Germinability of leek seeds and its improvement by osmopriming. Acta Hortic. 1994, 371, 45–52. [Google Scholar] [CrossRef]
- Jayawardhane, J.; Wijesinghe, M.K.P.S.; Bykova, N.V.; Igamberdiev, A.U. Metabolic changes in seed embryos of hypoxia-tolerant rice and hypoxia-sensitive barley at the onset of germination. Plants 2021, 10, 2456. [Google Scholar] [CrossRef] [PubMed]
- Gedam, P.A.; Shirsat, D.V.; Arunachalam, T.; Ghosh, S.; Gawande, S.J.; Mahajan, V.; Gupta, A.J.; Singh, M. Screening of onion (Allium cepa L.) genotypes for weaterlogging tolerance. Front. Plant Sci. 2022, 12, 727262. [Google Scholar] [CrossRef]
- Romano, A.; Bravi, R. Hydrotime model to evaluate the effects of a set of priming agents on seed germination of two leek cultivars under water stress. Seed Sci. Technol. 2021, 49, 159–174. [Google Scholar] [CrossRef]
- Hanci, F.; Cingi, M.; Akinci, H. Influence of L-tryptophan and melatonin on germination of onion and leek seed at different temperaures. Turk. J. Agric. Res. 2019, 6, 214–221. [Google Scholar]
- Dong, L.; Hao, Z.; Li, Z.; Zhu, J.; Wang, Q. Enhancement of Welsh onion (Allium fistolosum L.) seed vigor by KNO3 priming. J. Agric. Sci. Tech. 2014, 16, 1345–1353. [Google Scholar]
- Padula, G.; Xia, X.; Szopińska, D.; Hołubowicz, R. Effect of air temperature and relative humidity on the stored Welsh onion (Allium fistulosum L.) seeds. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12956. [Google Scholar] [CrossRef]
- Thakur, M.; Sharma, P.; Anand, A.; Pandita, V.K.; Bhatia, A.; Pushkar, S. Raffinose and hexose sugar content during germination are related to Infrared Thermal Fingerprints of primed onion (Allium cepa L.) seeds. Front. Plant Sci. 2020, 11, 579037. [Google Scholar] [CrossRef]
- Barga, S.; Dilts, T.E.; Leger, E.A. Climate variability affects the germination strategies exhibited by arid land plants. Oecologia 2017, 185, 437–452. [Google Scholar] [CrossRef]
- Mitchell, J.; Johnston, I.G.; Bassel, G.W. Variability in seeds: Biological, ecological, and agricultural implications. J. Exp. Bot. 2017, 68, 809–817. [Google Scholar] [CrossRef] [Green Version]
- Pagano, A.; Folini, G.; Pagano, P.; Sincinelli, F.; Rossetto, A.; Macovei, A.; Balestrazzi, A. ROS accumulation as a hallmark of dehydration stress in primed and overprimed Medicago truncatula seeds. Agronomy 2022, 12, 268. [Google Scholar] [CrossRef]
- Pagano, A.; Zannino, L.; Pagano, P.; Doria, E.; Dondi, D.; Macovei, A.; Biggiogera, M.; de Sousa Araujo, S.; Balestrazzi, A. Changes in genotoxic stress response, ribogenesis and PAP (3’-phosphoadenosine 5’-phosphate) levels are associated with loss of desiccation tolerance in overprimed Medicago truncatula seeds. Plant Cell Environ. 2022, 45, 1457–1473. [Google Scholar] [CrossRef]
- Dorone, Y.; Boeynaems, S.; Flores, E.; Jin, B.; Hateley, S.; Bossi, F.; Lazarus, E.; Pennington, J.G.; Michiels, E.; De Decker, M.; et al. A prion-like protein regulator of seed germination undergoes hydration-dependent phase separation. Cell 2021, 184, 4284–4298. [Google Scholar] [CrossRef]
- Boeynaems, S.; Alberti, S.; Fawzi, N.L.; Mittag, T.; Polymenidou, M.; Rousseau, F.; Schymkowitz, J.; Shorter, J.; Wolozin, B.; Van Den Bosch, L.; et al. Protein phase separation: A new phase in cell biology. Trends Cell Biol. 2018, 28, 420–435. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Jia, X.; Xiang, Y.; Jiang, W. Histone modification and chromatin remodeling during the seed life cycle. Front. Plant Sci. 2022, 13, 865361. [Google Scholar] [CrossRef]
- Yang, D.; Zhao, F.; Zhu, D.; Chen, X.; Kong, X.; Wu, Y.; Chen, M.; Du, J.; Qu, L.-J.; Wu, Z. Progressive chromatin silencing of ABA biosynthesis genes permits seed germination in Arabidopsis. Plant Cell 2022, 34, 2871–2891. [Google Scholar] [CrossRef]
- Forti, C.; Ottobrino, V.; Bassolino, L.; Toppino, L.; Rotino, G.L.; Pagano, A.; Macovei, A.; Balestrazzi, A. Molecular dynamics of pre-germinative metabolism in primed eggplant (Solanum melongena L.) seeds. Hortic. Res. 2020, 7, 87. [Google Scholar] [CrossRef]
- Forti, C.; Ottobrino, V.; Doria, E.; Bassolino, L.; Toppino, L.; Rotino, G.L.; Pagano, A.; Macovei, A.; Balestrazzi, A. Hydropriming applied on fast germinating Solanum villosum Miller seeds: Impact on pre-germinative metabolism. Front. Plant Sci. 2021, 12, 639336. [Google Scholar] [CrossRef]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Pagano, A.; Araújo, S.S.; Macovei, A.; Leonetti, P.; Balestrazzi, A. The seed repair response during germination: Disclosing correlations between DNA repair, antioxidant response, and chromatin remodeling in Medicago truncatula. Front. Plant Sci. 2017, 8, 1972. [Google Scholar] [CrossRef]
- Pagano, A.; De Sousa Araújo, S.; Macovei, A.; Dondi, D.; Lazzaroni, S.; Balestrazzi, A. Metabolic and gene expression hallmarks of seed germination uncovered by sodium butyrate in Medicago truncatula. Plant Cell Environ. 2019, 42, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Forti, C.; Shankar, A.; Singh, A.; Balestrazzi, A.; Prasad, V.; Macovei, A. Seed priming improves germination on heavy-metal contaminated soil by inducing upregulation of genes involved in DNA repair and antioxidant response. Genes 2020, 11, 242. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Bray, C. DNA synthesis in osmoprimed leek (Allium porrum L.) seeds and evidence for repair and replication. Seed Sci. Res. 1993, 3, 15–23. [Google Scholar] [CrossRef]
- Khandagale, K.; Krishna, R.; Roylawar, P.; Ade, A.B.; Benke, A.; Shinde, B.; Singh, M.; Suresh, J.; Gawande, S.J.; Ashutosh Rai, A. Omics approaches in Allium research: Progress and way ahead. PeerJ 2020, 8, e9824. [Google Scholar] [CrossRef]
- Finkers, R.; van Kaauwen, M.; Ament, K.; Burger-Meijer, K.; Egging, R.; Huits, H.; Kodde, L.; Kroon, L.; Shigyo, M.; Sato, S.; et al. Insights from the first genome assembly of onion (Allium cepa). G3 Genes Genomes Gen. 2021, 11, jkab243. [Google Scholar] [CrossRef]
- Ghodke, P.; Khandagale, K.; Thangasamy, A.; Kulkarni, A.; Narwade, N.; Shirsat, D.; Randive, P.; Roylawar, P.; Singh, I.; Gawande, S.J.; et al. Comparative transcriptome analyses in contrasting onion (Allium cepa L.) genotypes for drought stress. PLoS ONE 2020, 15, e0237457. [Google Scholar] [CrossRef]
- Alamar, M.C.; Anastasiadi, M.; Lopez-Cobollo, R.; Bennett, M.H.; Thompson, A.J.; Turnbull, C.G.N.; Mohareb, F.; Terry, L.A. Transcriptome and phytohormone changes associated with ethylene-induced onion bulb dormancy. Postharvest Biol. Technol. 2020, 168, 111267. [Google Scholar] [CrossRef]
- Wu, S.; Ning, F.; Wu, X.; Wang, W. Proteomic characterization of differential abundant proteins accumulated between lower and upper epidermises of fleshy scales in onion (Allium cepa L.) bulbs. PLoS ONE 2016, 11, e0168959. [Google Scholar] [CrossRef]



{kind=link}
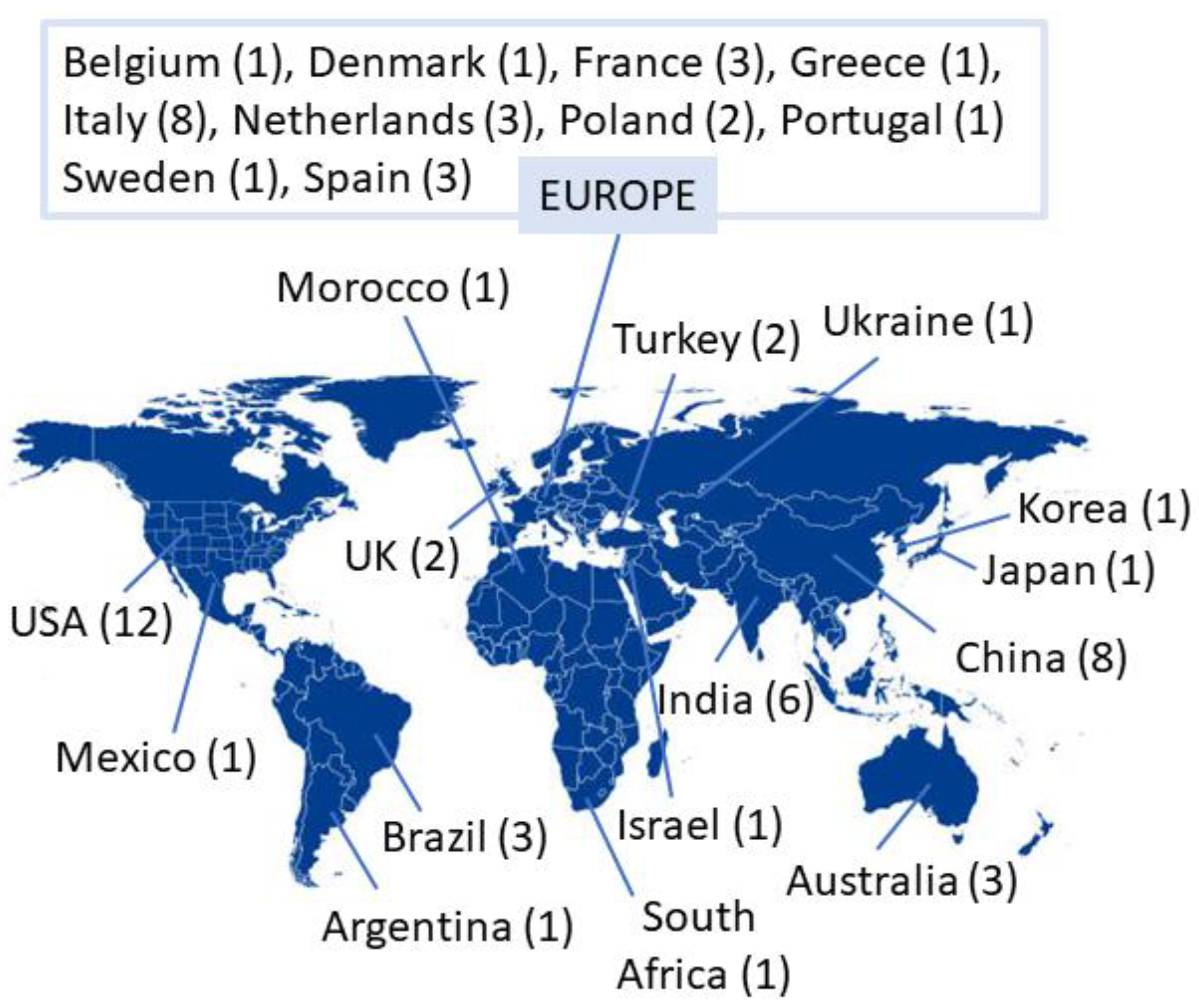{kind=link}
| Cultivar | Priming Method (*) | Reference |
|---|---|---|
| Granex 33 | hydropriming: water, 15 °C, 96 h | Caseiro et al. 2004 [44] |
| Granex 33 | drum priming: water, 15 °C/25 °C | Caseiro et al. 2004 [44] |
| White Lisbon; Rijnsburger Wijbo | osmopriming: 34.2% PEG 6000, 15 °C, 10 d | Dearman et al. 1986 [46] |
| White Lisbon; Senshy Semi Globe Yelllow | osmopriming: −1.4 MPa, 34.2% PEG 6000, 20 °C, 7 d | Ellis & Butcher 1988 [31] |
| Rijnsburger Robusta | osmopriming: −1.5 MPa, 34.2% PEG 6000, EA, 15 °C, 14 d | Bujalski & Nienow 1991 [34] |
| Bronze Wonder; Challenger; Big Mac; White Keeper | osmopriming: 30% PEG 8000, 10 °C, 7 d | Murray et al. 1992 [33] |
| Granex 33 | osmopriming: 30% PEG 8000, 15 °C, 24/48 h | Caseiro et al. 2004 [44] |
| Arka Kalyan | osmopriming: −1.5 MPa, PEG 6000, 25 °C, 24 h; osmopriming: 3% KNO3, 25 °C, 24 h | Muruli et al. 2016 [49] |
| Punjab Red-48 | osmopriming: 25% PEG 8000, 20 °C, 5 d | Basra et al. 1994 [47] |
| Hisar-2; Hisar Onion-3; Hisar Onion-4 | osmopriming: 0.5% KNO3, 16 h; osmopriming: 0.5% KH2PO4,16 h | Brar et al. 2019 [48] |
| Furanui; Tsukihikari; Early Grove | osmopriming: 0.4 M trehalose /raffinose, 15 °C, 4−6 d | Horita & Saruyama 2006 [36] |
| CO (On) 5 | nutripriming: 0.5% ZnSO4, r.t., 10 h | Saranya et al. 2017 [38] |
| Agrifound Dark Red | halopriming: 3% KNO3, r.t., 12 h | Thejeshwini et al. 2019 [41] |
| Arka Kalyan | hormonal priming: 50 ppm GA3, 25 °C, 24 h | Muruli et al. 2016 [49] |
| CO (On) 5 | hormonal priming: 0.2% methionine, r.t, 8 h | Saranya et al. 2017 [38] |
| Agrifound Dark Red | hormonal priming: 100 ppm GA3, r.t., 6 h | Thejeshwini et al. 2019 [41] |
| Hisar-2; Hisar Onion-3; Hisar Onion-4 | hormonal priming: 50 ppm GA3, 16 h | Brar et al. 2019 [48] |
| IPA 11 | hormonal priming: 50 μM SA, 25 °C, 24 h | Da Silva et al. 2019 [45] |
| Hisar-2; Hisar Onion-3; Hisar Onion-4 | biopriming: Azotobacter strain HT−57, 16 h | Brar et al. 2019 [48] |
| 1015Y Texas Super-sweet onion (variety Legend) | nanopriming: 31.3 ppm AgNPs, r.t., 12 h; nanopriming: 5.4 ppm AuNPs, r.t., 12 h | Acharya et al. 2019 [50] |
| Species/Cultivar | Priming Method (*) | Reference |
|---|---|---|
| Leek/Winterreuzen | osmopriming: −15 bars, 34.2% PEG, 15 °C, 7/14/21 d | Brocklehurst et al. 1984 [57] |
| Leek/Verina | osmopriming: PEG 8000, 15 °C, 14 d | Clarke & James 1991 [56] |
| Leek/Verina | osmopriming: −1.0 MPa, PEG 6000, 15 °C, 14 d | Davison & Bray 1991 [55] |
| Leek/Verina | osmopriming: −1.5 MPa, PEG 8000/D−mannitol/KNO3, 15 °C, 10 d | Parera & Cantliff 1992 [58] |
| Leek/Premier | osmopriming: −15 bars, PEG 6000, 15 °C, 1−14 d | Corbineau et al. 1994 [59] |
| Leek/Lungo della riviera; Monstrueux de Carentan | osmopriming: −1.8 MPa, PEG 8000/KCl/CaCl2/K2HPO4, 20 °C, 96 h | Romano & Bravi 2021 [62] |
| Welsh onion/Zhangqiu | osmopriming: 2% KNO3, 25 °C, 24 h | Dong et al. 2014 [64] |
| Leek/Inegol-92 | hormonal priming: 125 ppm L−tryptophan, 20 °C, 24 h, dark; hormonal priming: 5 μM melatonin, 20 °C, 24 h, dark | Hanci et al. 2019 [63] |
| Key Words | Timeline | Publications (N°) |
|---|---|---|
| seed priming | 1970–2023 | 1.151 |
| seed priming, rice | 1995–2023 | 148 |
| seed priming, wheat | 1980–2023 | 129 |
| seed priming, maize | 1970–2023 | 77 |
| seed priming, tomato | 1995–2023 | 52 |
| seed priming, soybean | 1985–2023 | 43 |
| seed priming, pepper | 2000–2023 | 7 |
| seed priming, onion | 2010–2023 | 7 |
| seed priming, leek | 2010–2023 | 4 |
| seed priming, eggplant | 2017–2023 | 4 |
| Product Commercial Name | Target Crop | Description | Company (City, Country) |
|---|---|---|---|
| Promotor onion | onion | tailored priming protocol to improve the speed of emergence and results in more uniform germination | Incotec (Enkhuizen, The Netherlands) |
| Incotec 118 Special | onion | a melting pellet allowing the seed quick access to oxygen, combined with a priming treatment for faster and more uniform germination | Incotec (Enkhuizen, The Netherlands) |
| Promoter leek | leek | priming formulation used to obtain faster, more uniform germination, and stronger crop stand; it broadens the temperature range at which seed will germinate; promotes photo-dormancy and thermo-dormancy release | Incotec (Enkhuizen, The Netherlands) |
| Promoter Plus Leek | leek | combination of priming and upgrading technology developed to enhance stand establishment through uniform germination, and to increase speed of emergence; increases the number of usable transplants | Incotec (Enkhuizen, The Netherlands) |
| B-Mox® | onion | seed enhancing formulation, promotes germination and seedling growth (5% improvement compared to stsandard priming treatments) | Bejo (Warmenhuizen, The Netherlands) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pagano, A.; Macovei, A.; Xia, X.; Padula, G.; Hołubowicz, R.; Balestrazzi, A. Seed Priming Applied to Onion-Like Crops: State of the Art and Open Questions. Agronomy 2023, 13, 288. https://doi.org/10.3390/agronomy13020288
Pagano A, Macovei A, Xia X, Padula G, Hołubowicz R, Balestrazzi A. Seed Priming Applied to Onion-Like Crops: State of the Art and Open Questions. Agronomy. 2023; 13(2):288. https://doi.org/10.3390/agronomy13020288
Chicago/Turabian StylePagano, Andrea, Anca Macovei, Xianzong Xia, Gregorio Padula, Roman Hołubowicz, and Alma Balestrazzi. 2023. "Seed Priming Applied to Onion-Like Crops: State of the Art and Open Questions" Agronomy 13, no. 2: 288. https://doi.org/10.3390/agronomy13020288