


Exogenous Easily Extractable Glomalin-Related Soil Protein Induces Differential Response in Plant Growth of Tea Plants via Regulating Water Channel Protein Expression

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
2.3. Variable Determinations
2.4. Statistical Analysis
3. Results and Analysis
3.1. Plant Growth Performance and Biomass
3.2. Root Morphology
3.3. Tea Quality
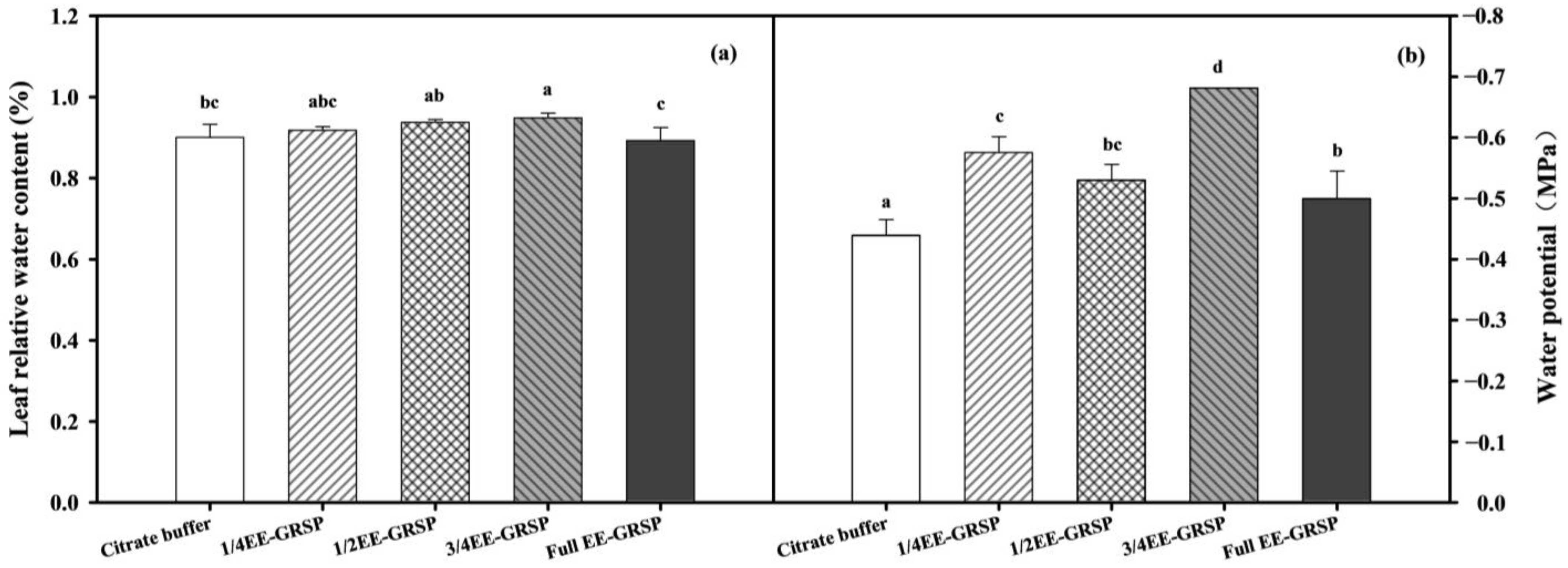
3.4. Leaf Water Status
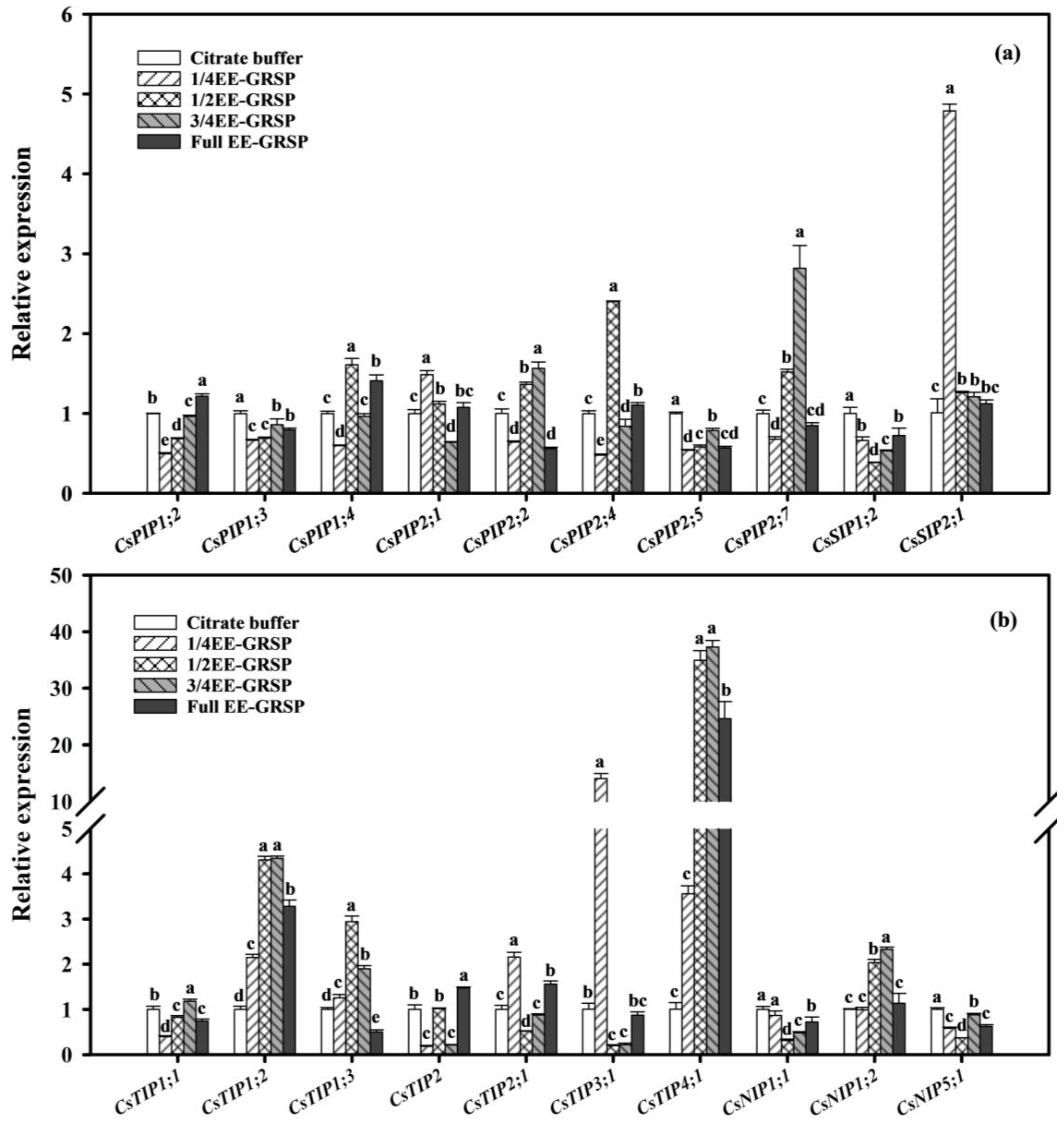
3.5. Relative Expression of AQP Genes in Roots
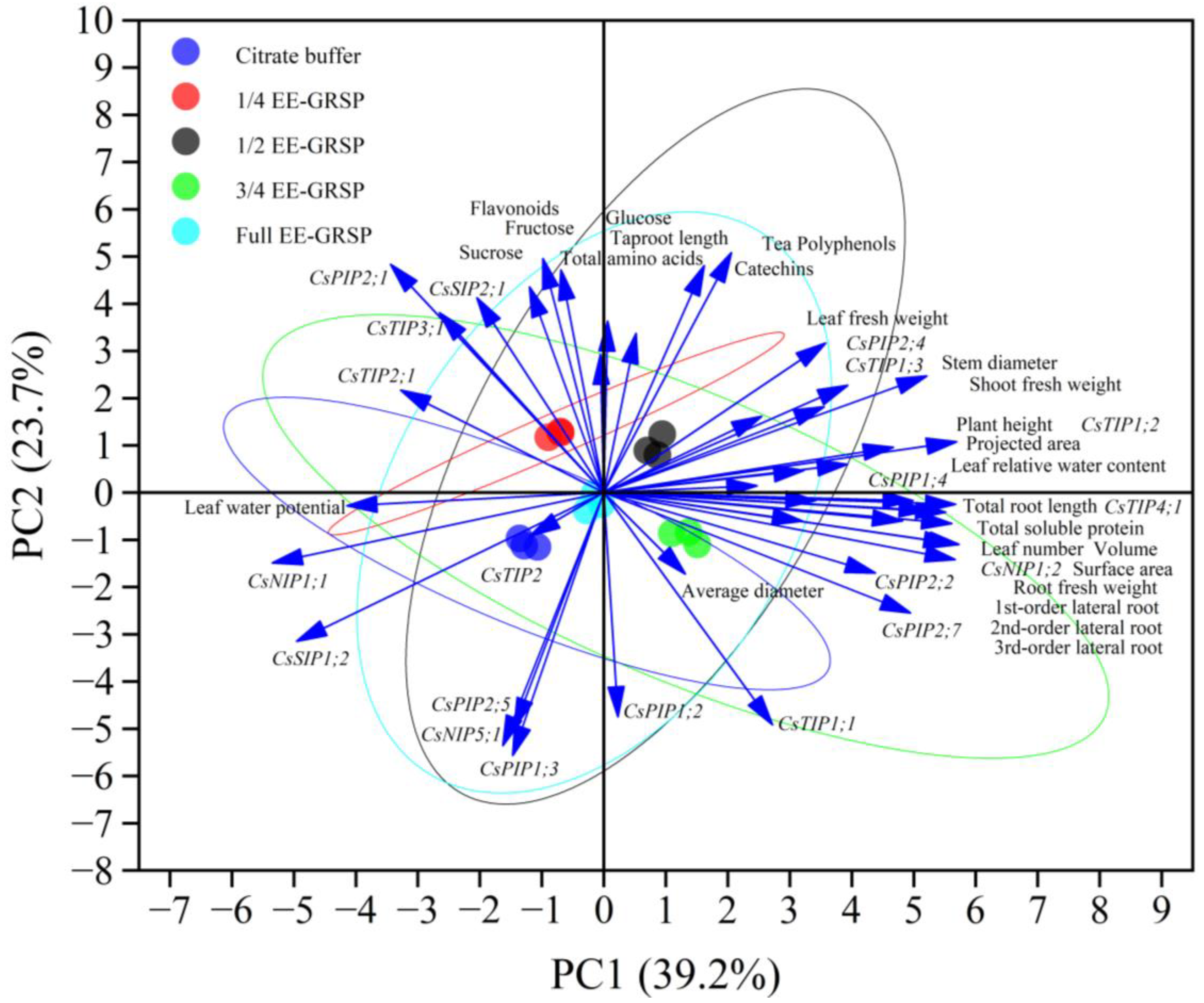
3.6. Correlation Analysis and Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Diagne, N.; Ngom, M.; Djighaly, P.-I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of arbuscular mycorrhizal fungi on plant growth and performance: Importance in biotic and abiotic stressed regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Schindler, F.-V.; Mercer, E.-J.; Rice, J.-A. Chemical characteristics of glomalin-related soil protein (GRSP) extracted from soils of varying organic matter content. Soil Biol. Biochem. 2007, 39, 320–329. [Google Scholar] [CrossRef]
- Driver, J.-D.; Holben, W.-E.; Rillig, M.-C. Characterization of glomalin as a hyphal wall component of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2005, 37, 101–106. [Google Scholar] [CrossRef]
- Rillig, M.-C. Arbuscular mycorrhizae, glomalin, and soil aggregation. Can. J. Soil Sci. 2004, 84, 355–363. [Google Scholar] [CrossRef]
- Bradford, M.-M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Koide, R.-T.; Peoples, M.-S. Behavior of Bradford-reactive substances is consistent with predictions for glomalin. Appl. Soil Ecol. 2013, 63, 8–14. [Google Scholar] [CrossRef]
- Wang, S.; Wu, Q.-S.; He, X.-H. Exogenous easily extractable glomalin-related soil protein promotes soil aggregation, relevant soil enzyme activities and plant growth in trifoliate orange. Plant Soil Environ. 2015, 61, 66–71. [Google Scholar] [CrossRef]
- Chi, G.-G.; Srivastava, A.-K.; Wu, Q.-S. Exogenous easily extractable glomalin-related soil protein improves drought tolerance of trifoliate orange. Arch. Agron. Soil Sci. 2018, 64, 1341–1350. [Google Scholar] [CrossRef]
- Xu, F.-J. Study on The Mechanism of Endophytic Fungus Phomopsis Liquidambar Formosan a Promoting the Symbiosis of Arbuscular Mycorrhizal Fungi and Peanut and the Drought Resistance of Symbionts; Nanjing Normal University: Nanjing, China, 2021. [Google Scholar] [CrossRef]
- Liu, R.-C.; Gao, W.-Q.; Srivastava, A.-K.; Zou, Y.-N.; Kuča, K.; Hashem, A.; Abd_Allah, E.-F.; Wu, Q.-S. Differential effects of exogenous glomalin-related soil proteins on plant growth of trifoliate orange through regulating auxin changes. Front. Plant Sci. 2021, 12, 745402. [Google Scholar] [CrossRef]
- Zou, Y.-N.; Srivastava, A.-K.; Wu, Q.-S.; Huang, Y.-M. Glomalin-related soil protein and water relations in mycorrhizal citrus (Citrus tangerina) during soil water deficit. Arch. Agron. Soil Sci. 2014, 60, 1103–1114. [Google Scholar] [CrossRef]
- Li, G.; Santoni, V.; Maurel, C. Plant aquaporins: Roles in plant physiology. BBA-Gen. Subj. 2014, 1840, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Verdoucq, L.; Luu, D.-T.; Santoni, V. Plant aquaporins: Membrane channels with multiple integrated functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef]
- Danielson, J.-Å.; Johanson, U. Unexpected complexity of the aquaporin gene family in the moss Physcomitrella patens. Bmc. Plant Biol. 2008, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.-J.; Rong, Z.-Y.; Wu, Q.-S.; Abd-Allah, E.-F.; Liu, C.-Y.; Liu, S.-R. Mycorrhiza improves plant growth and photosynthetic characteristics of tea plants in response to drought stress. Biocell 2022, 46, 1339–1346. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Guo, X.-N.; Wu, X.-L.; Dai, F.-J.; Wu, Q.-S. The comprehensive effects of Rhizophagus intraradices and P on root system architecture and P transportation in Citrus limon L. Agriculture 2022, 12, 317. [Google Scholar] [CrossRef]
- Guo, X.-N.; Hao, Y.; Wu, X.-L.; Chen, X.; Liu, C.-Y. Exogenous easily extractable glomalin-related soil protein stimulates plant growth by regulating tonoplast intrinsic protein expression in lemon. Plants 2023, 12, 2955. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Zou, Y.-N.; Zhan, T.-T.; Liu, C.-Y. Polyamines participate in mycorrhizal and root development of citrus (Citrus tangerine) seedlings. Not. Bot. Horti Agrobot. 2010, 38, 25–31. [Google Scholar] [CrossRef]
- Cao, J.-L.; Shao, Y.-D.; Zou, Y.-N.; Wu, Q.-S.; Yang, T.-Y.; Kuča, K. Inoculation with Clariodeoglomus etunicatum improves leaf food quality of tea exposed to P stress. Not. Bot. Horti Agrobot. 2021, 49, 12166. [Google Scholar] [CrossRef]
- Chu, Y.-H.; Chang, C.-L.; Hsu, H.-F. Flavonoid content of several vegetables and their antioxidant activity. J. Sci. Food Agr. 2000, 80, 561–566. [Google Scholar] [CrossRef]
- Livak, K.-J.; Schmittgen, T.-D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zou, Y.-N.; Srivastava, A.-K.; Wu, Q.-S. Glomalin: A potential soil conditioner for perennial fruits. Int. J. Agric. Biol. 2016, 18, 293–297. [Google Scholar] [CrossRef]
- Rillig, M.-C.; Wright, S.-F.; Eviner, V.-T. The role of arbuscular mycorrhizal fungi and glomalin in soil aggregation: Comparing effects of five plant species. Plant Soil 2002, 238, 325–333. [Google Scholar] [CrossRef]
- Gao, W.-Q.; Chi, G.-G.; Liu, C.-Y.; Wu, Q.-S. Evaluation of exogenous application of EE-GRSP on soil aggregation and root hormone levels in trifoliate orange (Poncirus trifoliata) under drought stress. Curr. Hortic. 2018, 6, 7–11. Available online: https://www.indianjournals.com/ijor.aspx?target=ijor:chr&volume=6&issue=2&article=002 (accessed on 4 June 2019).
- Najarian, A.; Souri, M.-K.; Nabigol, A. Influence of humic substance on vegetative growth, flowering and leaf mineral elements of Pelargonium x hortorum. J. Plant Nutr. 2022, 45, 107–112. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Q.; Wang, H.; Nie, S.; Liang, Z. Effects of soil salinity on the content, composition, and ion binding capacity of glomalin-related soil protein (GRSP). Sci. Total Environ. 2017, 581, 657–665. [Google Scholar] [CrossRef]
- He, J.-D.; Chi, G.-G.; Zou, Y.-N.; Shu, B.; Wu, Q.-S.; Srivastava, A.-K.; Kuča, K. Contribution of glomalin-related soil proteins to soil organic carbon in trifoliate orange. Appl. Soil Ecol. 2020, 154, 103592. [Google Scholar] [CrossRef]
- Adamczyk, B.; Smolander, A.; Kitunen, V.; Godlewski, M. Proteins as nitrogen source for plants: A short story about exudation of proteases by plant roots. Plant Signal. Behav. 2010, 5, 817–819. [Google Scholar] [CrossRef]
- Chen, W.; Mou, X.; Meng, P.; Chen, J.; Tang, X.; Meng, G.; Xin, K.; Zhang, Y.; Wang, C. Effects of arbuscular mycorrhizal fungus inoculation on the growth and nitrogen metabolism of Catalpa bungei CA Mey. under different nitrogen levels. Front. Plant Sci. 2023, 14, 1138184. [Google Scholar] [CrossRef]
- Li, D.; Li, C.; Sun, H.; Wang, W.; Liu, L.; Zhang, Y. Effects of drought on soluble protein content and protective enzyme system in cotton leaves. Front. Agr. China 2010, 4, 56–62. [Google Scholar] [CrossRef]
- Boyer, J.-S.; James, R.-A.; Munns, R.; Condon, T.-A.; Passioura, J.-B. Osmotic adjustment leads to anomalously low estimates of relative water content in wheat and barley. Funct. Plant Biol. 2008, 35, 1172–1182. [Google Scholar] [CrossRef]
- Aharon, R.; Shahak, Y.; Wininger, S.; Bendov, R.; Kapulnik, Y.; Galili, G. Overexpression of a plasma membrane aquaporin in transgenic tobacco improves plant vigor under favorable growth conditions but not under drought or salt stress. Plant Cell 2003, 15, 439–447. [Google Scholar] [CrossRef]
- Heinen, R.-B.; Ye, Q.; Chaumont, F. Role of aquaporins in leaf physiology. J. Exp. Bot. 2009, 60, 2971–2985. [Google Scholar] [CrossRef]
- Bari, A.; Farooq, M.; Hussain, A.; Qamar, M.-T.; Abbas, M.-W.; Mustafa, G.; Karim, A.; Ahmed, I.; Hussain, T. Genome-wide bioinformatics analysis of aquaporin gene family in maize (Zea mays L.). J. Phylogenet. Evol. Biol. 2018, 6, 1000197. [Google Scholar] [CrossRef]
- Zargar, S.-M.; Nagar, P.; Deshmukh, R.; Nazir, M.; Wani, A.-A.; Masoodi, K.-Z.; Agrawal, G.-K.; Rakwal, R. Aquaporins as potential drought tolerance inducing proteins: Towards instigating stress tolerance. J. Proteomics 2017, 169, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C. Aquaporins and water permeability of plant membranes. Annu. Rev. Plant Biol. 1997, 48, 399–429. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; von Wirén, N. Ammonium as a signal for physiological and morphological responses in plants. J. Exp. Bot. 2017, 68, 2581–2592. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, H.; Hachez, C.; Bienert, M.-D.; Beebo, A.; Swarup, K.; Voß, U.; Bouhidel, K.; Frigerio, L.; Schjoerring, J.-K.; Bennett, M.-J.; et al. Tonoplast aquaporins facilitate lateral root emergence. Plant Physiol. 2016, 170, 1640–1654. [Google Scholar] [CrossRef]
- Singh, R.-K.; Deshmukh, R.; Muthamilarasan, M.; Rani, R.; Prasad, M. Versatile roles of aquaporin in physiological processes and stress tolerance in plants. Plant Physiol. Biochem. 2020, 149, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Zangi, R.; Filella, M. Transport routes of metalloids into and out of the cell: A review of the current knowledge. Chem. Biol. Interact. 2012, 197, 47–57. [Google Scholar] [CrossRef]
- Noronha, H.; Agasse, A.; Martins, A.-P.; Berny, M.-C.; Gomes, D.; Zarrouk, O.; Thiebaud, P.; Delrot, S.; Soveral, G.; Chaumont, F.; et al. The grape aquaporin VvSIP1 transports water across the ER membrane. J. Exp. Bot. 2014, 65, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.-A.; Haque, A.-M.; Akther, M.-S.; Islam, M.; Lee, K.-W.; Kabir, A.-H. The NIP Genes in Sugar Beet: Underlying Roles in Silicon Uptake and Growth Improvement. Silicon-Neth 2021, 14, 3551–3562. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID | Primer Sequence | Gene | Gene ID | Primer Sequence |
|---|---|---|---|---|---|
| CsPIP1;2 | XM_028242073.1 | F-AGATCGTCGGTACCTTTGTC | CsTIP1;1 | XM_028239672.1 | F-GTTTGTCACCAATGGCATGA |
| R-GACTCCTTGCTGGGTTGATA | R-ATAATTCCCAACTCGCCCTT | ||||
| CsPIP1;3 | XM_028197133.1 | F-TGGTATCTCAGGAGGACACA | CsTIP1;2 | XM_028241002.1 | F-TCGGTAGGGGCTAACATTTC |
| R-AATCTCAGCACCAAGACCAT | R-CTCGAAAACTAGTGCGTTCC | ||||
| CsPIP1;4 | XM_028209956.1 | F-TTGGTGCACTTAGCCACTAT | CsTIP1;3 | XM_028271063.1 | F-ATAAGCTGACGGATAACGGG |
| R-TCACGACTTGGTGGTACAAA | R-ATTGAACCCAAACACTGTGC | ||||
| CsPIP2;1 | XM_028250124.1 | F-TCCAGAGCTCCTACTACGAC | CsTIP2 | XM_028212337.1 | F-TCATTCAGTGTTGGGTCACT |
| R-GGCTAGGTGAACCATGAACA | R-AGGTGACAGCTGGATTCAAA | ||||
| CsPIP2;2 | XM_028201387.1 | F-ATTCTCAGCCAAGGACTACG | CsTIP2;1 | XM_028220182.1 | F-TTGGTTTCATTGTTGGTGCC |
| R-TGCAGTATACGAGGACGAAG | R-TTTGGATAGATGAGTCCGGC | ||||
| CsPIP2;4 | XM_028247087.1 | F-GGTGGAATGATCTTCGTCCT | CsTIP3;1 | XM_028223148.1 | F-CGCGCTTTTATTGGAGATCG |
| R-GACGTAGTAGGCCTTTTGGA | R-GACCGAGCCAATAGATCCAG | ||||
| CsPIP2;5 | XM_028241134.1 | F-TCTCAGCCAAGGATTACCAC | CsTIP4;1 | XM_028269043.1 | F-GTCATCTTAATCCGGCGGTA |
| R-GGGTCAATCTGGCTCTTGTA | R-GGGAGTAGTCCATTCCACTG | ||||
| CsPIP2;7 | XM_028229390.1 | F-ACATTTGGGTTGTTCTTGGC | CsNIP1;1 | XM_028206307.1 | F-TTAACCCTGCTGTCACCATT |
| R-AACAGTGTAAACGAGGACGA | R-ACACGAAAGATTGCATGTCG | ||||
| CsSIP1;2 | XM_028198192.1 | F-TAACCACCGTCCTTGTCTTC | CsNIP1;2 | XM_028232417.1 | F-TTAACCCTGCAGTCACCATT |
| R-TGTTTTTGTACTGCATGGGC | R-GAAAGACTGTGCGTCAGAAC | ||||
| CsSIP2;1 | XM_028214111.1 | F-GGGGATTTCAGTCGATTCCT | CsNIP5;1 | XM_028230765.1 | F-GCCTCTTAGATTTCCCTGCT |
| R-ATGTCAGAAATCCCTCCGTC | R-GAGGGAGGGATTGAGATGTG | ||||
| Cs-GADPH | XM_002263109 | F-TTGGCATCGTTGAGGGTCT | |||
| R-CAGTGGGAACACGGAAAGC |
| Treatments | Plant Height (cm) | Stem Diameter (mm) | Leaf Number (#/Plant) | Biomass (g FW/Plant) | ||
|---|---|---|---|---|---|---|
| Leaf | Shoot | Root | ||||
| Citrate buffer | 15.18 ± 1.48 b | 2.44 ± 0.30 b | 7.83 ± 0.75 d | 1.78 ± 0.13 b | 0.58 ± 0.05 c | 3.22 ± 0.11 c |
| 1/4 EE-GRSP | 16.52 ± 1.18 b | 2.58 ± 0.25 b | 9.67 ± 0.82 c | 2.66 ± 0.18 a | 0.74 ± 0.03 b | 3.89 ± 0.22 b |
| 1/2 EE-GRSP | 19.58 ± 1.90 a | 3.04 ± 0.29 a | 12.67 ± 1.51 b | 2.50 ± 0.22 a | 0.89 ± 0.07 a | 3.91 ± 0.15 b |
| 3/4 EE-GRSP | 19.25 ± 2.76 a | 2.75 ± 0.20 ab | 15.33 ± 1.86 a | 2.66 ± 0.18 a | 0.86 ± 0.03 a | 4.59 ± 0.44 a |
| Full EE-GRSP | 16.35 ± 1.47 b | 2.63 ± 0.22 b | 12.00 ± 1.41 b | 2.55 ± 0.17 a | 0.78 ± 0.06 b | 4.55 ± 0.45 a |
| Treatments | Total Length (cm) | Projected Area (cm²) | Surface Area (cm²) | Average Diameter (mm) | Volume (cm³) | Taproot Length (cm) | Lateral Root Numbers (#/Plant) | ||
|---|---|---|---|---|---|---|---|---|---|
| First-Order | Second-Order | Third-Order | |||||||
| Citrate Buffer | 102.97 ± 8.49 c | 10.13 ± 0.94 b | 9.78 ± 0.23 c | 0.75 ± 0.09 a | 1.14 ± 0.11 d | 10.70 ± 0.89 b | 77.50 ± 4.65 c | 389.50 ± 38.54 d | 39.00 ± 2.00 d |
| 1/4 EE-GRSP | 116.38 ± 9.84 bc | 10.91 ± 1.02 ab | 10.39 ± 0.36 bc | 0.73 ± 0.09 a | 1.44 ± 0.07 c | 10.68 ± 0.91 b | 91.00 ± 7.30 c | 411.50 ± 37.97 d | 44.00 ± 4.08 d |
| 1/2 EE-GRSP | 126.99 ± 1.55 ab | 11.61 ± 0.40 ab | 10.90 ± 0.49 b | 0.73 ± 0.05 a | 1.63 ± 0.11 b | 11.95 ± 0.67 a | 128.00 ± 10.23 b | 599.00 ± 55.78 b | 93.00 ± 7.39 b |
| 3/4 EE-GRSP | 141.37 ± 5.74 a | 11.71 ± 0.59 ab | 12.04 ± 0.42 a | 0.81 ± 0.07 a | 2.02 ± 0.13 a | 10.05 ± 0.42 b | 171.25 ± 13.77 a | 748.75 ± 24.92 a | 143.50 ± 9.28 a |
| Full EE-GRSP | 123.08 ± 10.40 b | 12.12 ± 0.94 a | 10.09 ± 0.70 bc | 0.68 ± 0.04 a | 1.67 ± 0.07 b | 10.70 ± 0.73 b | 119.25 ± 7.50 b | 495.00 ± 11.34 c | 67.25 ± 6.24 c |
| Treatments | Fructose (mg/g) | Sucrose (mg/g) | Glucose (mg/g) | Tea Polyphenols (mg/g) | Total Amino Acids (mg/g) | Catechins (mg/g) | Flavonoids (mg/g) | Total Soluble Protein (mg/g) |
|---|---|---|---|---|---|---|---|---|
| Citrate buffer | 29.25 ± 1.22 c | 49.49 ± 2.13 bc | 14.65 ± 0.82 b | 80.03 ± 1.10 c | 30.26 ± 3.66 b | 24.40 ± 2.34 c | 83.50 ± 3.16 b | 5.88 ± 0.88 b |
| 1/4 EE-GRSP | 32.56 ± 0.97 a | 55.11 ± 2.98 a | 14.76 ± 0.21 b | 92.83 ± 3.57 ab | 30.07 ± 1.59 b | 40.47 ± 1.72 a | 92.11 ± 0.87 a | 6.52 ± 0.42 ab |
| 1/2 EE-GRSP | 31.49 ± 1.31 ab | 53.01 ± 1.32 ab | 16.99 ± 0.83 a | 97.41 ± 2.98 a | 39.37 ± 2.34 a | 37.45 ± 4.34 ab | 94.89 ± 2.45 a | 7.15 ± 1.03 ab |
| 3/4 EE-GRSP | 29.56 ± 1.11 bc | 47.84 ± 3.48 bc | 13.23 ± 0.37 c | 85.88 ± 4.89 bc | 22.22 ± 5.91 c | 33.09 ± 1.72 b | 75.78 ± 0.27 c | 8.04 ± 1.07 a |
| Full EE-GRSP | 31.56 ± 1.24 ab | 47.06 ± 3.58 c | 13.61 ± 0.68 bc | 86.92 ± 5.09 bc | 17.10 ± 0.33 c | 36.38 ± 4.14 ab | 77.33 ± 2.22 c | 7.65 ± 1.32 ab |
| CsNIP1;1 | CsNIP1;2 | CsNIP5;1 | CsPIP1;2 | CsPIP1;3 | CsPIP2;5 | CsPIP2;7 | CsTIP1;1 | CsTIP1;2 | CsTIP4;1 | CsSIP1;2 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant height | −0.964 ** | 0.943 * | −0.443 | −0.249 | −0.426 | −0.317 | 0.743 | 0.321 | 0.917 * | 0.877 | −0.933 * |
| Stem diameter | −0.962 ** | 0.768 | −0.708 | −0.280 | −0.561 | −0.475 | 0.436 | 0.116 | 0.852 | 0.808 | −0.926 * |
| Leaf number | −0.835 | 0.874 | −0.207 | 0.165 | −0.249 | −0.297 | 0.811 | 0.429 | 0.947 * | 0.942 * | −0.768 |
| Shoot fresh weight | −0.934 * | 0.783 | −0.64 | −0.179 | −0.643 | −0.635 | 0.541 | 0.057 | 0.971 ** | 0.889 * | −0.968 ** |
| Number of first-order lateral roots | −0.792 | 0.898 * | −0.070 | 0.208 | −0.122 | −0.157 | 0.884 * | 0.541 | 0.900 * | 0.918 * | −0.697 |
| Number of second-order lateral roots | −0.830 | 0.962 ** | −0.054 | 0.133 | −0.082 | −0.065 | 0.934 * | 0.610 | 0.886 * | 0.914 * | −0.715 |
| Number of third-order lateral roots | −0.792 | 0.949 * | 0.006 | 0.154 | −0.040 | −0.026 | 0.949 * | 0.632 | 0.859 | 0.890 * | −0.676 |
| Total length of root system | −0.811 | 0.858 | −0.226 | 0.059 | −0.321 | −0.346 | 0.801 | 0.351 | 0.931 * | 0.899 * | −0.792 |
| Root surface area | −0.726 | 0.915 * | −0.015 | −0.123 | −0.169 | −0.076 | 0.927 * | 0.477 | 0.783 | 0.757 | −0.711 |
| Root volume | −0.745 | 0.793 | −0.176 | 0.159 | −0.28 | −0.348 | 0.767 | 0.343 | 0.901 * | 0.876 | −0.722 |
| Leaf relative water content | −0.751 | 0.893 * | −0.171 | −0.435 | −0.289 | −0.082 | 0.813 | 0.354 | 0.691 | 0.637 | −0.774 |
| Leaf soluble protein | −0.706 | 0.687 | −0.214 | 0.336 | −0.252 | −0.397 | 0.644 | 0.314 | 0.886 * | 0.882 * | −0.648 |
| Sucrose | 0.000 | −0.166 | −0.550 | −0.966 ** | −0.663 | −0.415 | −0.376 | −0.692 | −0.160 | −0.344 | −0.284 |
| Fructose | −0.089 | −0.294 | −0.830 | −0.526 | −0.909 * | −0.928 * | −0.556 | −0.917 * | 0.125 | −0.076 | −0.385 |
| Tea polyphenols | −0.630 | 0.305 | −0.945 * | −0.681 | −0.936 * | −0.820 | −0.064 | −0.526 | 0.542 | 0.371 | −0.825 |
| Catechins | −0.415 | 0.111 | −0.827 | −0.512 | −0.969 ** | −0.973 ** | −0.139 | −0.688 | 0.495 | 0.289 | −0.692 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.-L.; Hao, Y.; Dai, F.-J.; Chen, X.; Liu, C.-Y. Exogenous Easily Extractable Glomalin-Related Soil Protein Induces Differential Response in Plant Growth of Tea Plants via Regulating Water Channel Protein Expression. Agronomy 2023, 13, 2797. https://doi.org/10.3390/agronomy13112797
Wu X-L, Hao Y, Dai F-J, Chen X, Liu C-Y. Exogenous Easily Extractable Glomalin-Related Soil Protein Induces Differential Response in Plant Growth of Tea Plants via Regulating Water Channel Protein Expression. Agronomy. 2023; 13(11):2797. https://doi.org/10.3390/agronomy13112797
Chicago/Turabian StyleWu, Xiao-Long, Yong Hao, Feng-Jun Dai, Xin Chen, and Chun-Yan Liu. 2023. "Exogenous Easily Extractable Glomalin-Related Soil Protein Induces Differential Response in Plant Growth of Tea Plants via Regulating Water Channel Protein Expression" Agronomy 13, no. 11: 2797. https://doi.org/10.3390/agronomy13112797