
Exogenous Melatonin Alleviates the Inhibitory Effect of NaHCO3 on Tomato Growth by Regulating the Root pH Value and Promoting Plant Photosynthesis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions and Treatments
2.2. Assessment of Various Indexes
2.2.1. Measurement of Growth Indicators
2.2.2. Endogenous MT Levels
2.2.3. Determination of pH and Organic Acid Content in the Root Environment
2.2.4. Chlorophyll Content
2.2.5. Photosynthetic Gas-Exchange Parameters
2.2.6. Rapid Chlorophyll Fluorescence-Induced Kinetics (OJIP) Curve Measurement and JIP-Test Parameters
2.2.7. Determination of the Expression of Key Genes of Photosynthetic Carbon Assimilative Enzymes
2.3. Data Analysis
3. Results
3.1. Growth Traits, Biomass, and Endogenous MT Levels
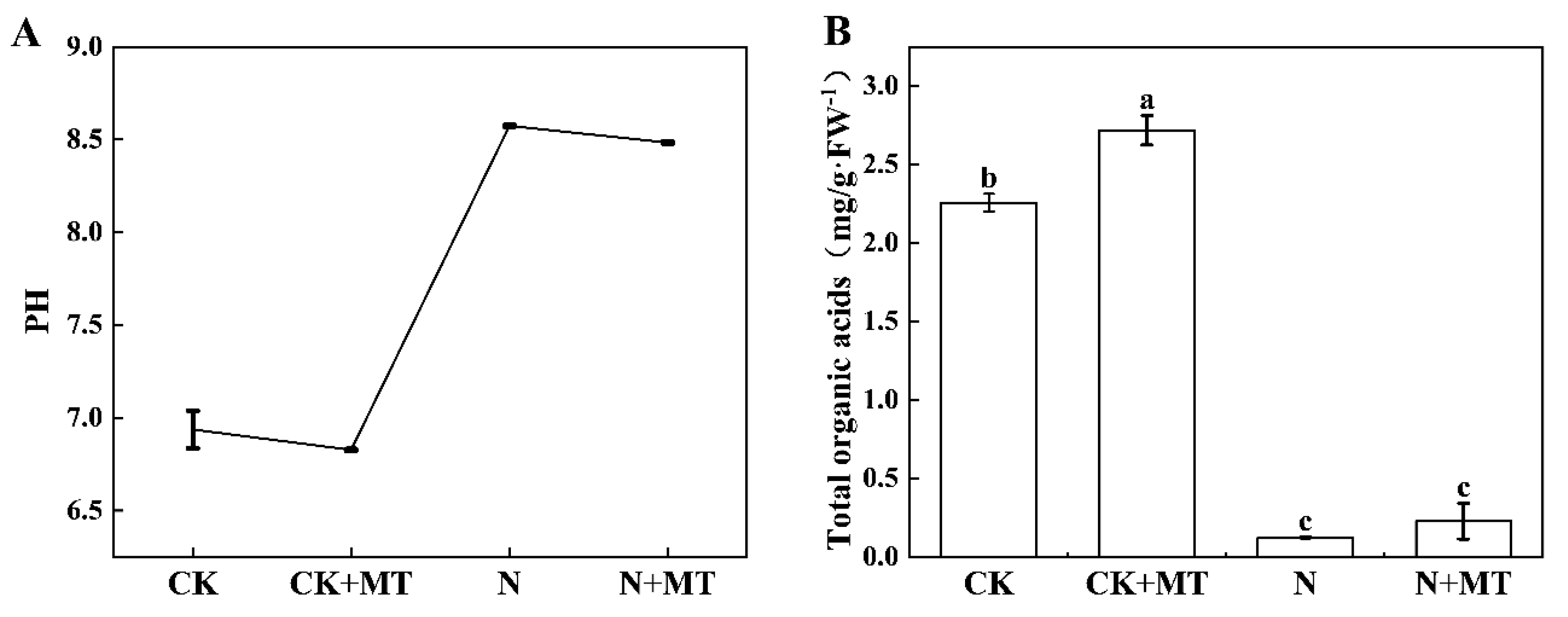
3.2. pH of the Root Environment
3.3. Chlorophyll Content
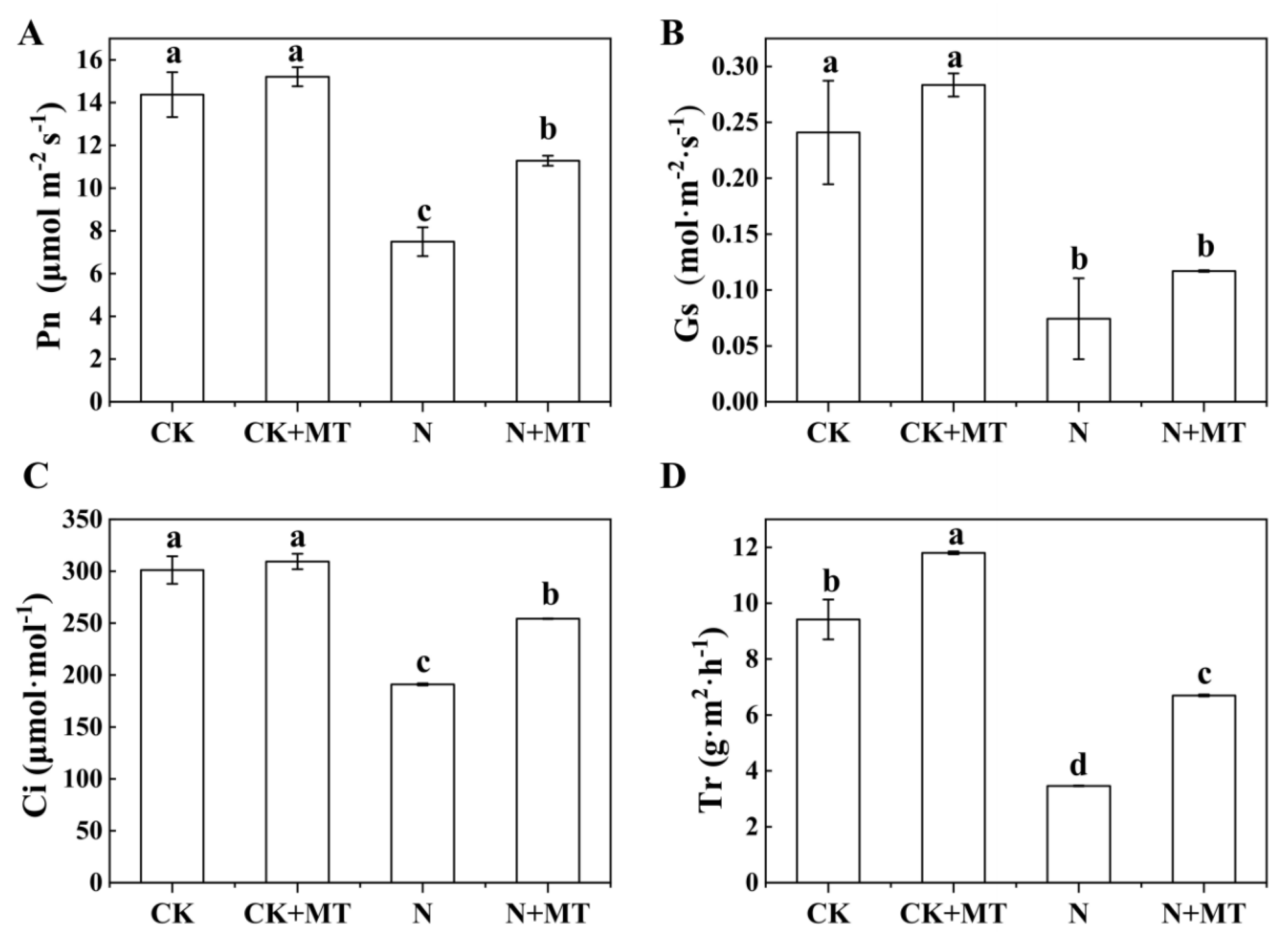
3.4. Photosynthetic Gas-Exchange Parameters
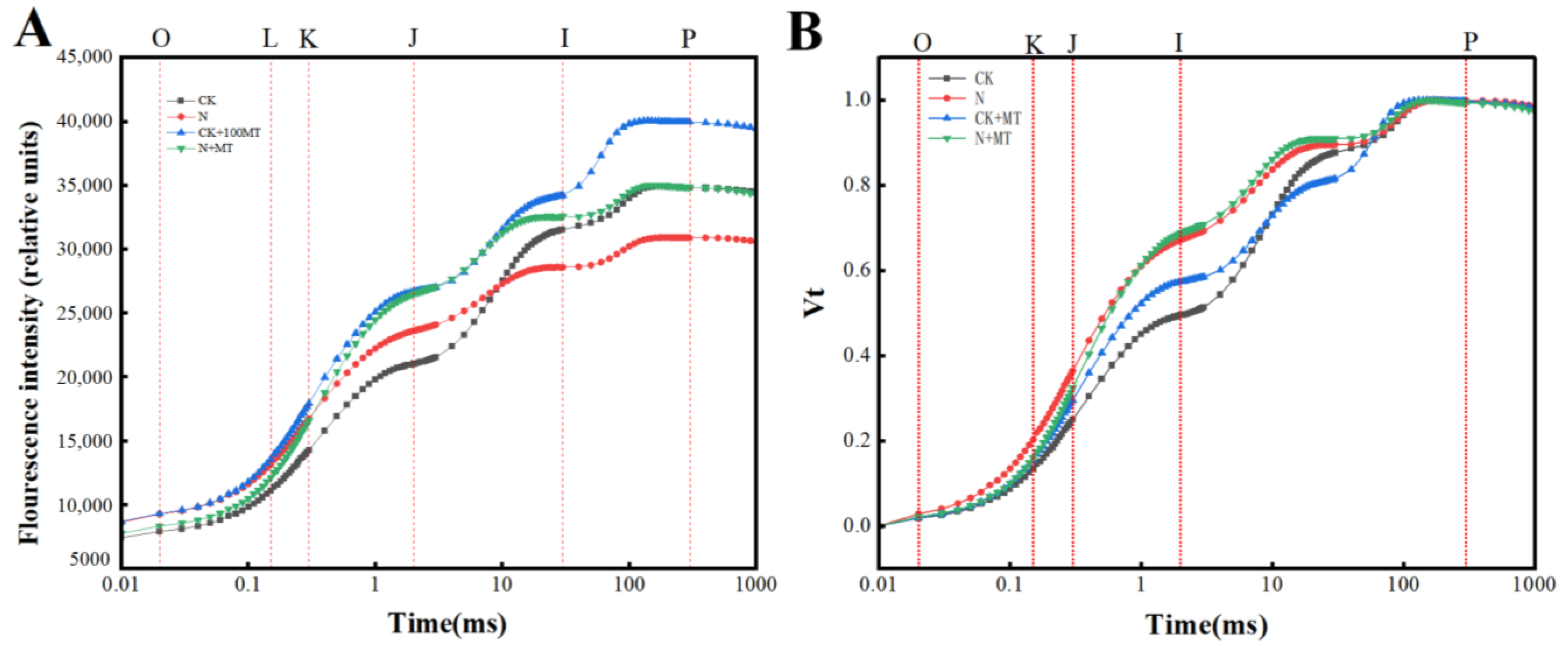
3.5. OJIP Curves
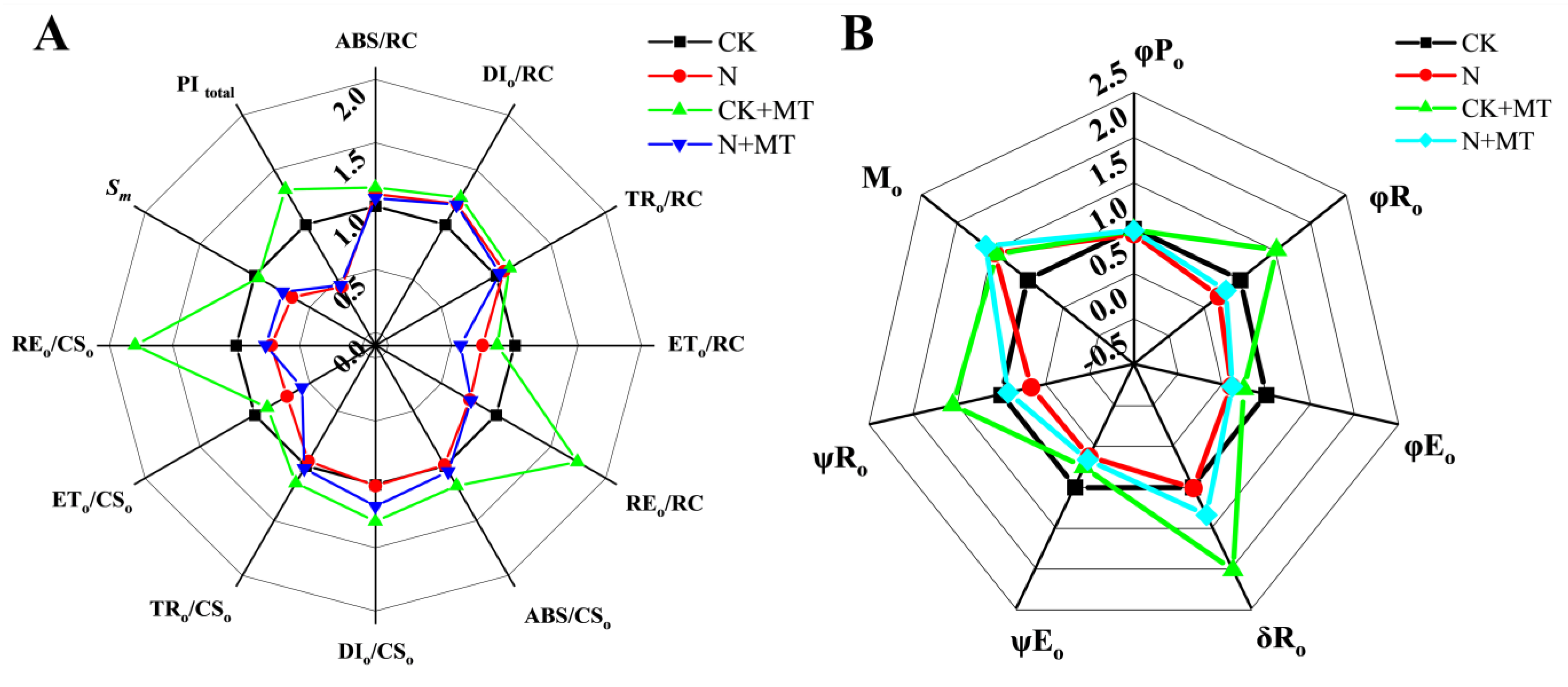
3.6. JIP-Test Parameters
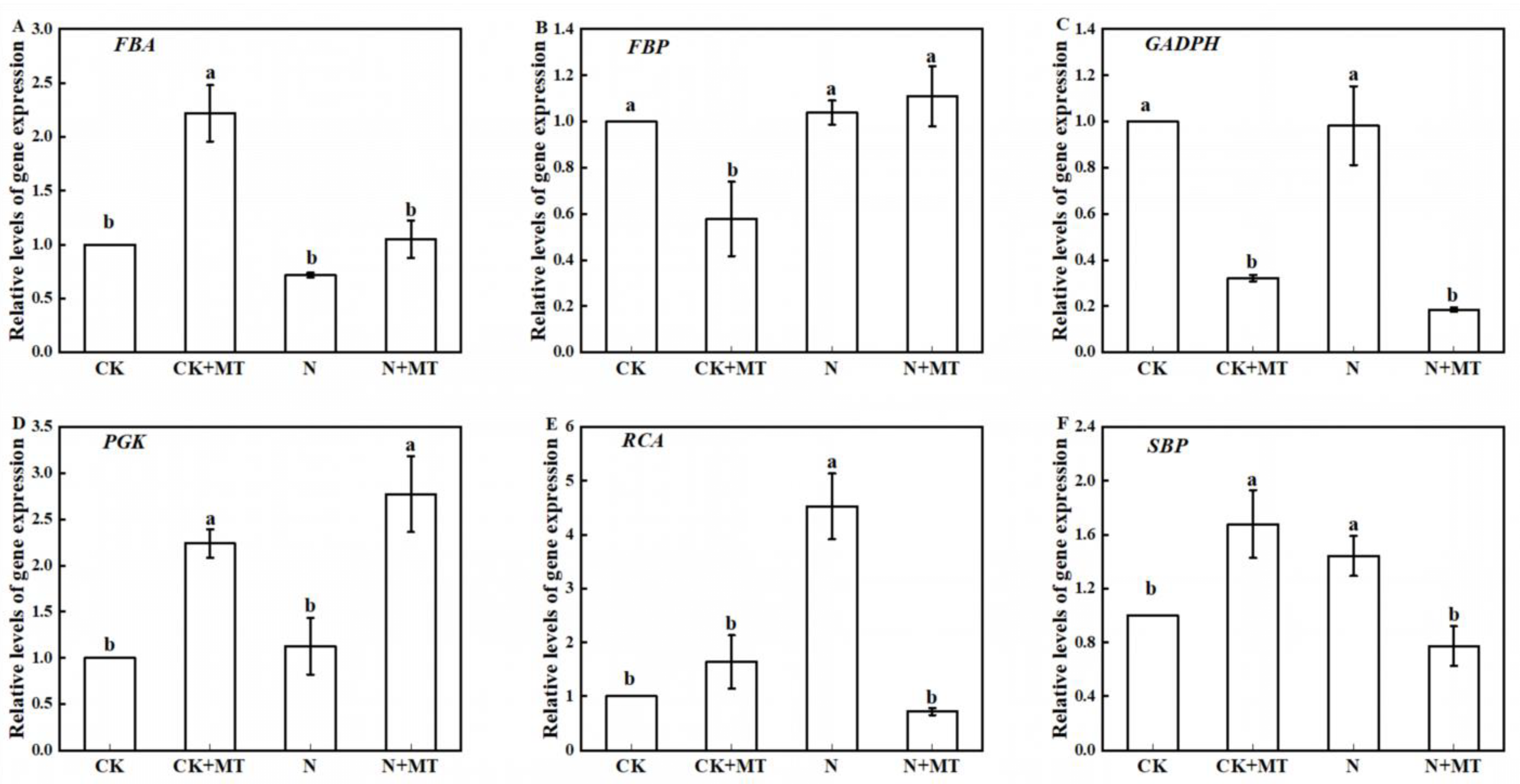
3.7. Expression of Key Genes for Photosynthetic Carbon Assimilation Enzymes
4. Discussion
4.1. Effect of Exogenous MT on Growth and Endogenous MT Levels in Tomato Seedlings under NaHCO3 Stress
4.2. Effect of Exogenous MT Application on the pH of the Root Environment of Tomato Seedlings under NaHCO3 Stress
4.3. Effects of Exogenous MT on the Content of Photosynthetic Pigments and Photosynthetic Gas-Exchange Parameters in Tomato Seedlings under NaHCO3 Stress
4.4. Effect of Exogenous MT on Chlorophyll Fluorescence Parameters of Tomato Seedlings under NaHCO3 Stress
4.5. Effect of Exogenous MT on the Expression of Key Enzyme Genes of the Calvin Cycle in Tomato Seedlings under NaHCO3 Stress Conditions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manasa, M.R.K.; Katukuri, N.R.; Darveekaran Nair, S.S.; Haojie, Y.; Yang, Z.; Guo, R. bo Role of Biochar and Organic Substrates in Enhancing the Functional Characteristics and Microbial Community in a Saline Soil. J. Environ. Manag. 2020, 269, 110737. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Chong, J.; Li, C.; Kim, C.; Shi, D.; Wang, D. Osmotic Adjustment and Ion Balance Traits of an Alkali Resistant Halophyte Kochia Sieversiana during Adaptation to Salt and Alkali Conditions. Plant Soil 2007, 294, 263–276. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Yan, X.; Guo, J. Physiological and Transcriptomic Analyses of Yellow Horn (Xanthoceras sorbifolia) Provide Important Insights into Salt and Saline-Alkali Stress Tolerance. PLoS ONE 2020, 15, e0244365. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, N.; Wang, H.; Ren, J.; Yao, Y. Hydraulic properties and carbon metabolism of Lycium barbarum and Tamarix tamarisk under salinity stress. J. Plant Sci. 2017, 35, 865–873. [Google Scholar]
- Li, N.; Zhang, Z.; Gao, S.; Lv, Y.; Chen, Z.; Cao, B.; Xu, K. Different Responses of Two Chinese Cabbage (Brassica rapa L. Ssp. Pekinensis) Cultivars in Photosynthetic Characteristics and Chloroplast Ultrastructure to Salt and Alkali Stress. Planta 2021, 254, 102. [Google Scholar] [CrossRef]
- Wang, X.; Ren, H.; Wei, Z.; Wang, Y.; Ren, W. Effects of Neutral Salt and Alkali on Ion Distributions in the Roots, Shoots, and Leaves of Two Alfalfa Cultivars with Differing Degrees of Salt Tolerance. J. Integr. Agric. 2017, 16, 1800–1807. [Google Scholar] [CrossRef]
- Wang, J.; Peng, X.; Gao, Y.; Wang, Y.; Lin, J.; Yan, X. Physiological and Proteomics Analyses Reveal the Resistance Response Mechanism to Alkali Stress in the Early Seedlings (Cotyledons vs. Roots) of Castor Plant (Ricinus communis L.). Environ. Exp. Bot. 2021, 185, 104414. [Google Scholar] [CrossRef]
- Guo, J.J.; Xu, X.X.; Zhang, R.D.; Chen, X.F.; Xing, Y.F.; Li, B.; Liu, C.; Zhou, Y.F. Effect of Short-Term Combined Alkaline Stress on Antioxidant Metabolism, Photosynthesis, and Leaf-Air Temperature Difference in Sorghum. Photosynthetica 2022, 60, 200–211. [Google Scholar] [CrossRef]
- Paredes, S.D.; Korkmaz, A.; Manchester, L.C.; Tan, D.-X.; Reiter, R.J. Phytomelatonin: A Review. J. Exp. Bot. 2009, 60, 57–69. [Google Scholar] [CrossRef]
- Sun, C.; Liu, L.; Wang, L.; Li, B.; Jin, C.; Lin, X. Melatonin: A Master Regulator of Plant Development and Stress Responses. J. Integr. Plant Biol. 2021, 63, 126–145. [Google Scholar] [CrossRef]
- Li, X.; Brestic, M.; Tan, D.-X.; Zivcak, M.; Zhu, X.; Liu, S.; Song, F.; Reiter, R.J.; Liu, F. Melatonin Alleviates Low PS I-Limited Carbon Assimilation under Elevated CO2 and Enhances the Cold Tolerance of Offspring in Chlorophyll b-Deficient Mutant Wheat. J. Pineal Res. 2018, 64, e12453. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, Y.; Sun, K.; Chen, Y.; Chen, X.; Li, X. Exogenous Melatonin Enhances Cold, Salt and Drought Stress Tolerance by Improving Antioxidant Defense in Tea Plant (Camellia sinensis (L.) O. Kuntze). Molecules 2019, 24, 1826. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, K.; Karuppasami, K.M.; Alagarswamy, S.; Shanmugam, K.P.; Rathinavelu, S.; Vellingiri, G.; Muniyappan, U.; Kanthan, T.; Kuppusamy, A.; Rajendran, M.; et al. Role of Melatonin in Directing Plant Physiology. Agronomy 2023, 13, 2405. [Google Scholar] [CrossRef]
- Debnath, B.; Islam, W.; Li, M.; Sun, Y.; Lu, X.; Mitra, S.; Hussain, M.; Liu, S.; Qiu, D. Melatonin Mediates Enhancement of Stress Tolerance in Plants. Int. J. Mol. Sci. 2019, 20, 1040. [Google Scholar] [CrossRef]
- Ma, S.; Gai, P.; Geng, B.; Wang, Y.; Ullah, N.; Zhang, W.; Zhang, H.; Fan, Y.; Huang, Z. Exogenous Melatonin Improves Waterlogging Tolerance in Wheat through Promoting Antioxidant Enzymatic Activity and Carbon Assimilation. Agronomy 2022, 12, 2876. [Google Scholar] [CrossRef]
- Alyammahi, O.; Gururani, M.A. Chlorophyll-a Fluorescence Analysis Reveals Differential Response of Photosynthetic Machinery in Melatonin-Treated Oat Plants Exposed to Osmotic Stress. Agronomy 2020, 10, 1520. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.; Yu, Y.; Zeng, H.; Deng, L.; Zhu, L.; Chen, G.; Wang, Y. Melatonin-Induced Resilience Strategies against the Damaging Impacts of Drought Stress in Rice. Agronomy 2022, 12, 813. [Google Scholar] [CrossRef]
- Zhou, X.; Zhao, H.; Cao, K.; Hu, L.; Du, T.; Baluška, F.; Zou, Z. Beneficial Roles of Melatonin on Redox Regulation of Photosynthetic Electron Transport and Synthesis of D1 Protein in Tomato Seedlings under Salt Stress. Front. Plant Sci. 2016, 7, 1823. [Google Scholar] [CrossRef]
- Sun, S.; Wen, D.; Yang, W.; Meng, Q.; Shi, Q.; Gong, B. Overexpression of Caffeic Acid O-Methyltransferase 1 (COMT1) Increases Melatonin Level and Salt Stress Tolerance in Tomato Plant. J. Plant Growth Regul. 2020, 39, 1221–1235. [Google Scholar] [CrossRef]
- Wu, P.; Lv, J.; Yu, J.; Liu, N.; Li, J.; Jing, L. Effects of melatonin on photosynthetic characteristics and osmoregulatory substance contents of cucumber seedlings under combined saline and alkaline stress. J. Appl. Ecol. 2022, 33, 1901–1910. [Google Scholar] [CrossRef]
- Liu, N.; Jin, Z.; Wang, S.; Gong, B.; Wen, D.; Wang, X.; Wei, M.; Shi, Q. Sodic Alkaline Stress Mitigation with Exogenous Melatonin Involves Reactive Oxygen Metabolism and Ion Homeostasis in Tomato. Sci. Hortic. 2015, 181, 18–25. [Google Scholar] [CrossRef]
- ElSayed, A.I.; Rafudeen, M.S.; Gomaa, A.M.; Hasanuzzaman, M. Exogenous Melatonin Enhances the Reactive Oxygen Species Metabolism, Antioxidant Defense-related Gene Expression, and Photosynthetic Capacity of Phaseolus vulgaris L. to Confer Salt Stress Tolerance. Physiol. Plant. 2021, 173, 1369–1381. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Wang, Z.; Qi, M.; Meng, S.; Li, T. Effect of externally applied melatonin on photosynthetic function of tomato seedlings under salt stress. J. Ecol. 2019, 38, 467–475. [Google Scholar] [CrossRef]
- Jiang, X.; Niu, N.; Cui, H.; Wei, S.; Xiong, H.; Liu, H.; Diao, M. Effect of foliar spraying of melatonin on photosynthetic characteristics of tomato seedlings under NaHCO3 stress. Agric. Res. Arid Areas 2022, 40, 87–95. [Google Scholar]
- Li, X.; Wang, S.; Chen, X.; Cong, Y.; Cui, J.; Shi, Q.; Liu, H.; Diao, M. The Positive Effects of Exogenous Sodium Nitroprusside on the Plant Growth, Photosystem II Efficiency and Calvin Cycle of Tomato Seedlings under Salt Stress. Sci. Hortic. 2022, 299, 111016. [Google Scholar] [CrossRef]
- Hao, H.; Chang, J.; Zhang, Z. Methods for studying plant root secretion. Plant Physiol. Newsl. 2003, 39, 56–60. [Google Scholar] [CrossRef]
- Zhang, X.; Tan, G. Experimental Techniques in Plant Physiology; Liaoning Science and Technology Press: Shengyang, China, 1989. [Google Scholar]
- Yan, Y.; Liu, X. A study of methods for the determination of chlorophyll. J. Beijing Agric. Univ. 1982, 8, 53–67. [Google Scholar]
- Schansker, G.; Srivastava, A.; Strasser, R.J. Characterization of the 820-Nm Transmission Signal Paralleling the Chlorophyll a Fluorescence Rise (OJIP) in Pea Leaves. Funct. Plant Biol. 2003, 30, 785. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ibrahim, M.F.M.; Elbar, O.H.A.; Farag, R.; Hikal, M.; El-Kelish, A.; El-Yazied, A.A.; Alkahtani, J.; El-Gawad, H.G.A. Melatonin Counteracts Drought Induced Oxidative Damage and Stimulates Growth, Productivity and Fruit Quality Properties of Tomato Plants. Plants 2020, 9, 1276. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.-X.; Altaf, M.M.; Jahan, M.S.; Khan, L.U. Melatonin Mitigates Nickel Toxicity by Improving Nutrient Uptake Fluxes, Root Architecture System, Photosynthesis, and Antioxidant Potential in Tomato Seedling. J. Soil Sci. Plant Nutr. 2021, 21, 1842–1855. [Google Scholar] [CrossRef]
- Hu, E.; Liu, M.; Zhou, R.; Jiang, F.; Sun, M.; Wen, J.; Zhu, Z.; Wu, Z. Relationship between Melatonin and Abscisic Acid in Response to Salt Stress of Tomato. Sci. Hortic. 2021, 285, 110176. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Khan, M.N.; Mukherjee, S.; Basahi, R.A.; Alamri, S.; Al-Amri, A.A.; Alsubaie, Q.D.; Ali, H.M.; Al-Munqedhi, B.M.A.; Almohisen, I.A.A. Exogenous Melatonin-mediated Regulation of K+/Na+ Transport, H+-ATPase Activity and Enzymatic Antioxidative Defence Operate through Endogenous Hydrogen Sulphide Signalling in NaCl-stressed Tomato Seedling Roots. Plant Biol. J. 2021, 23, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Yang, X.; Ma, C.; Wang, Y.; Zhao, J. Melatonin Enhances Drought Stress Tolerance in Maize through Coordinated Regulation of Carbon and Nitrogen Assimilation. Plant Physiol. Biochem. 2021, 167, 958–969. [Google Scholar] [CrossRef]
- Mansouri, S.; Sarikhani, H.; Sayyari, M.; Soleimani Aghdam, M. Melatonin Accelerates Strawberry Fruit Ripening by Triggering GAMYB Gene Expression and Promoting ABA Accumulation. Sci. Hortic. 2021, 281, 109919. [Google Scholar] [CrossRef]
- Sharafi, Y.; Aghdam, M.S.; Luo, Z.; Jannatizadeh, A.; Razavi, F.; Fard, J.R.; Farmani, B. Melatonin Treatment Promotes Endogenous Melatonin Accumulation and Triggers GABA Shunt Pathway Activity in Tomato Fruits during Cold Storage. Sci. Hortic. 2019, 254, 222–227. [Google Scholar] [CrossRef]
- Ni, J.; Wang, Q.; Shah, F.A.; Liu, W.; Wang, D.; Huang, S.; Fu, S.; Wu, L. Exogenous Melatonin Confers Cadmium Tolerance by Counterbalancing the Hydrogen Peroxide Homeostasis in Wheat Seedlings. Molecules 2018, 23, 799. [Google Scholar] [CrossRef]
- Robin, A.H.K.; Matthew, C.; Uddin, M.J.; Bayazid, K.N. Salinity-Induced Reduction in Root Surface Area and Changes in Major Root and Shoot Traits at the Phytomer Level in Wheat. J. Exp. Bot. 2016, 67, 3719–3729. [Google Scholar] [CrossRef]
- Xiang, G.; Ma, W.; Gao, S.; Jin, Z.; Yue, Q.; Yao, Y. Transcriptomic and Phosphoproteomic Profiling and Metabolite Analyses Reveal the Mechanism of NaHCO3-Induced Organic Acid Secretion in Grapevine Roots. BMC Plant Biol 2019, 19, 383. [Google Scholar] [CrossRef]
- Gong, B.; Li, X.; VandenLangenberg, K.M.; Wen, D.; Sun, S.; Wei, M.; Li, Y.; Yang, F.; Shi, Q.; Wang, X. Overexpression of S-adenosyl-l-methionine Synthetase Increased Tomato Tolerance to Alkali Stress through Polyamine Metabolism. Plant Biotechnol. J. 2014, 12, 694–708. [Google Scholar] [CrossRef]
- Wei, W.; Li, Q.-T.; Chu, Y.-N.; Reiter, R.J.; Yu, X.-M.; Zhu, D.-H.; Zhang, W.-K.; Ma, B.; Lin, Q.; Zhang, J.-S.; et al. Melatonin Enhances Plant Growth and Abiotic Stress Tolerance in Soybean Plants. J. Exp. Bot. 2015, 66, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Li, A.; Yang, M.; Song, B.; Gao, Y.; Li, H. Effects of externally applied melatonin on growth and physiological characteristics of safflower under salt stress. Chin. Herb. Med. 2019, 42, 1730–1737. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Kunderlikova, K.; Olsovska, K.; Allakhverdiev, S.I. Effect of Photosystem I Inactivation on Chlorophyll a Fluorescence Induction in Wheat Leaves: Does Activity of Photosystem I Play Any Role in OJIP Rise? J. Photochem. Photobiol. B Biol. 2015, 152, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Tikkanen, M.; Rantala, S.; Aro, E.-M. Electron Flow from PSII to PSI under High Light Is Controlled by PGR5 but Not by PSBS. Front. Plant Sci. 2015, 6, 521. [Google Scholar] [CrossRef]
- Ayyaz, A.; Amir, M.; Umer, S.; Iqbal, M.; Bano, H.; Gul, H.S.; Noor, Y.; Kanwal, A.; Khalid, A.; Javed, M.; et al. Melatonin Induced Changes in Photosynthetic Efficiency as Probed by OJIP Associated with Improved Chromium Stress Tolerance in Canola (Brassica napus L.). Heliyon 2020, 6, e04364. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Li, H.J.; Liu, J.; Bai, Y.W.; Xue, J.Q.; Zhang, R.H. Melatonin Alleviates Drought-Induced Damage of Photosynthetic Apparatus in Maize Seedlings. Russ. J. Plant Physiol. 2020, 67, 312–322. [Google Scholar] [CrossRef]
- Qi, X.-Y.; Wang, W.-L.; Hu, S.-Q.; Liu, M.-Y.; Zheng, C.-S.; Sun, X.-Z. Effects of Exogenous Melatonin on Photosynthesis and Physiological Characteristics of Chry-Santhemum Seedlings under High Temperature Stress. Ying Yong Sheng Tai Xue Bao 2021, 32, 2496–2504. [Google Scholar] [CrossRef]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Pérez-Pérez, M.E.; Francia, F.; Danon, A.; Marchand, C.H.; Fermani, S.; Trost, P.; et al. Redox Regulation of the Calvin–Benson Cycle: Something Old, Something New. Front. Plant Sci. 2013, 4, 470. [Google Scholar] [CrossRef]
- Lv, G.-Y.; Guo, X.-G.; Xie, L.-P.; Xie, C.-G.; Zhang, X.-H.; Yang, Y.; Xiao, L.; Tang, Y.-Y.; Pan, X.-L.; Guo, A.-G.; et al. Molecular Characterization, Gene Evolution, and Expression Analysis of the Fructose-1, 6-Bisphosphate Aldolase (FBA) Gene Family in Wheat (Triticum aestivum L.). Front. Plant Sci. 2017, 8, 1030. [Google Scholar] [CrossRef]
- Tristan, C.; Shahani, N.; Sedlak, T.W.; Sawa, A. The Diverse Functions of GAPDH: Views from Different Subcellular Compartments. Cell. Signal. 2011, 23, 317–323. [Google Scholar] [CrossRef]
- Liang, D.; Ni, Z.; Xia, H.; Xie, Y.; Lv, X.; Wang, J.; Lin, L.; Deng, Q.; Luo, X. Exogenous Melatonin Promotes Biomass Accumulation and Photosynthesis of Kiwifruit Seedlings under Drought Stress. Sci. Hortic. 2019, 246, 34–43. [Google Scholar] [CrossRef]
- Ding, F.; Liu, B.; Zhang, S. Exogenous Melatonin Ameliorates Cold-Induced Damage in Tomato Plants. Sci. Hortic. 2017, 219, 264–271. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, Z.; Zhang, Y.; Bai, L.; Hu, X.; Li, X.; Zhang, L.; Miao, Y.; Wang, Y. Melatonin Reduces Photoinhibition in Cucumber during Chilling by Regulating the Calvin-Benson Cycle. Sci. Hortic. 2022, 299, 111007. [Google Scholar] [CrossRef]
- Sharma, S.; Joshi, J.; Kataria, S.; Verma, S.K.; Chatterjee, S.; Jain, M.; Pathak, K.; Rastogi, A.; Brestic, M. Chapter 27—Regulation of the Calvin Cycle under Abiotic Stresses: An Overview. In Plant Life under Changing Environment; Tripathi, D.K., Pratap Singh, V., Chauhan, D.K., Sharma, S., Prasad, S.M., Dubey, N.K., Ramawat, N., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 681–717. ISBN 978-0-12-818204-8. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Acid | Regression Equation | Correlation Coefficient |
|---|---|---|
| oxalic acid | Y = 57,711.33x + 91.14 | R = 0.999 |
| malic acid | Y = 613.1733x + 0.3166 | R = 0.999 |
| citric acid | Y = 654.123x − 1.71992 | R = 0.999 |
| succinic acid | Y = 386.08x + 3.0225 | R = 0.999 |
| Parameters and Formulae | Definition |
|---|---|
| Minimal recorded fluorescence intensity | |
| Maximal recorded fluorescence intensity | |
| Variable fluorescence | |
| Performance index | |
| Reaction center Cross-section of the sample (t = tfm) | |
| Absorption flux per RC | |
| Trapped energy flux per RC | |
| Electron-transport flux per RC | |
| Dissipated energy flux per RC | |
| Trapped energy flux per CS at t = to | |
| Electron-transport flux per CS at t = to | |
| Dissipated energy flux per CS at t = to | |
| Absorption flux per CS at t = to | |
| Density of reaction centers per cross-section (at t = to) | |
| The normalized area between the J–P phase and the line F = Fm | |
| Maximum photochemical efficiency of PSII (at t = 0) | |
| Quantum ratios of PSII electron transfer (at t = 0) | |
| Quantum efficiency of PSI receptor-side terminal electron-acceptor reduction | |
| Reflects the efficiency of individual electrons traveling from the electron-transport chain between PSI and PSII of the photosystem to the terminal electron acceptor on the receptor side of PSI | |
| Maximum rate at which QA is reduced |
| Gene | Gene-ID | Sequence of Forward Primer | Sequence of Reverse Primer |
|---|---|---|---|
| RCA | LOC101250725 | TTGGACGGATTCTACATCGC | CTCCCCAAACACCCAAAATAAG |
| PGK | LOC101253805 | GAAGAGCGTTGGAGACCTTAG | AGTGTTTGATGGTAGGGATGG |
| GADPH | LOC100736499 | ACTCTGGTATATGTGTTACTC | AGGGAAGCAAGATTACTAAA |
| SBP | LOC100316873 | AGAAATACACCTTGAGATACACCG | TCAAGAATCCTAACGGTGCC |
| FBP | LOC101264273 | AATTTCCATCTCTTCCCCACC | TCGGTTTCTTGATCTGTGCTG |
| FBA | LOC1181999 | CTGTATGGACCGATGGACTTAC | AAGGTCTAAAGGGTAAGCTACATAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Guan, S.; Jiang, X.; Li, M.; Wei, S.; Diao, M. Exogenous Melatonin Alleviates the Inhibitory Effect of NaHCO3 on Tomato Growth by Regulating the Root pH Value and Promoting Plant Photosynthesis. Agronomy 2023, 13, 2777. https://doi.org/10.3390/agronomy13112777
Yang Y, Guan S, Jiang X, Li M, Wei S, Diao M. Exogenous Melatonin Alleviates the Inhibitory Effect of NaHCO3 on Tomato Growth by Regulating the Root pH Value and Promoting Plant Photosynthesis. Agronomy. 2023; 13(11):2777. https://doi.org/10.3390/agronomy13112777
Chicago/Turabian StyleYang, Yuanling, Sihui Guan, Xiyao Jiang, Ming Li, Shaowei Wei, and Ming Diao. 2023. "Exogenous Melatonin Alleviates the Inhibitory Effect of NaHCO3 on Tomato Growth by Regulating the Root pH Value and Promoting Plant Photosynthesis" Agronomy 13, no. 11: 2777. https://doi.org/10.3390/agronomy13112777