
Short-Term Dynamics of Bacterial Community Structure in Restored Abandoned Agricultural Soils under Semi-Arid Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Details and Field Experimental Design
2.2. Soil Physico-Chemical Properties Measurements
2.3. Bacterial Community Composition and Sequencing Data Processing
2.4. Statistical Analysis
3. Results
3.1. Changes in Soil Physico-Chemical Properties and Stoichiometry Nutrients over Time
3.2. Bacterial Community Composition
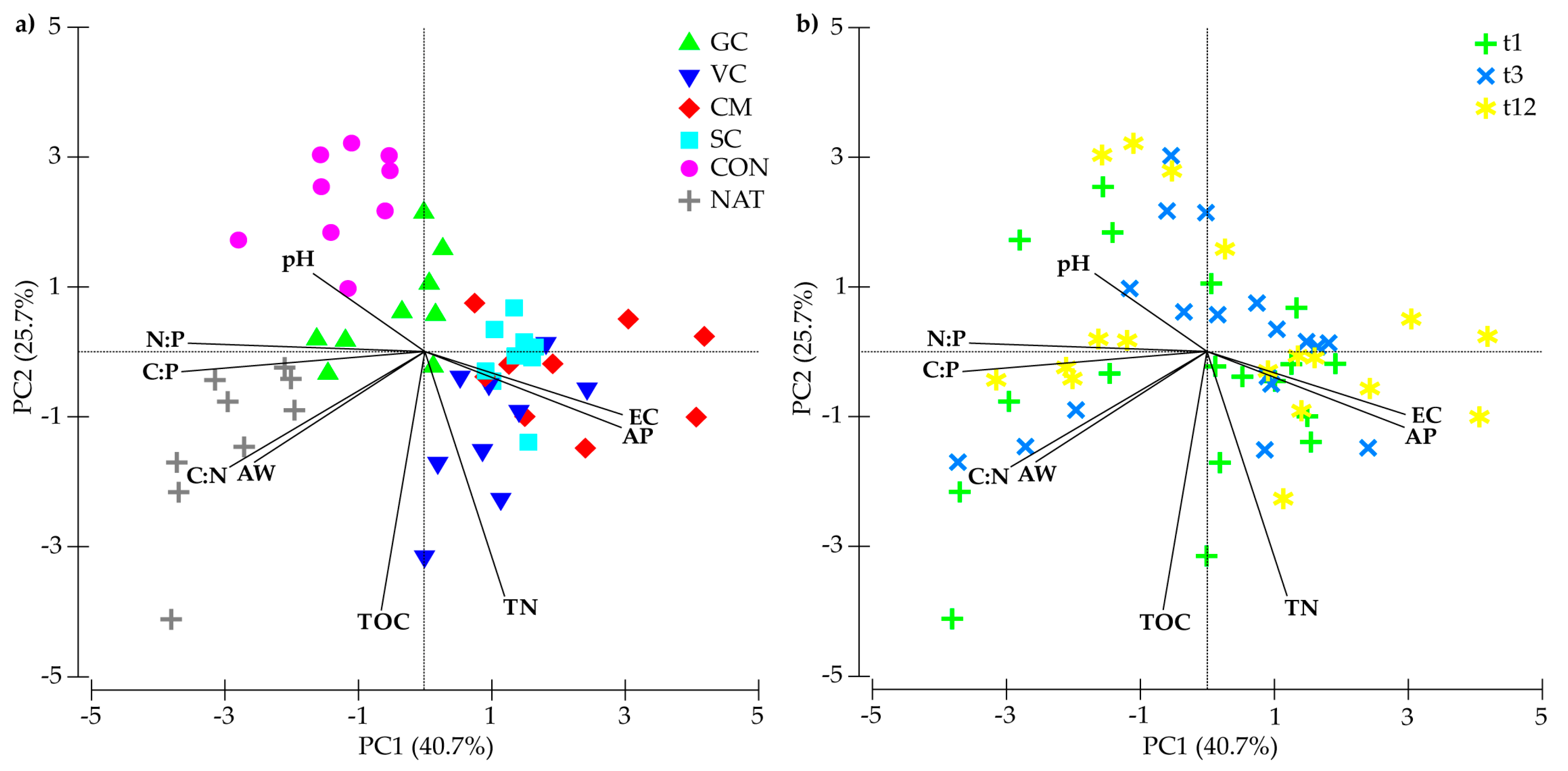
3.3. Relationships among Soil Chemical Properties, Nutrient Stoichiometry, and Bacterial Communities
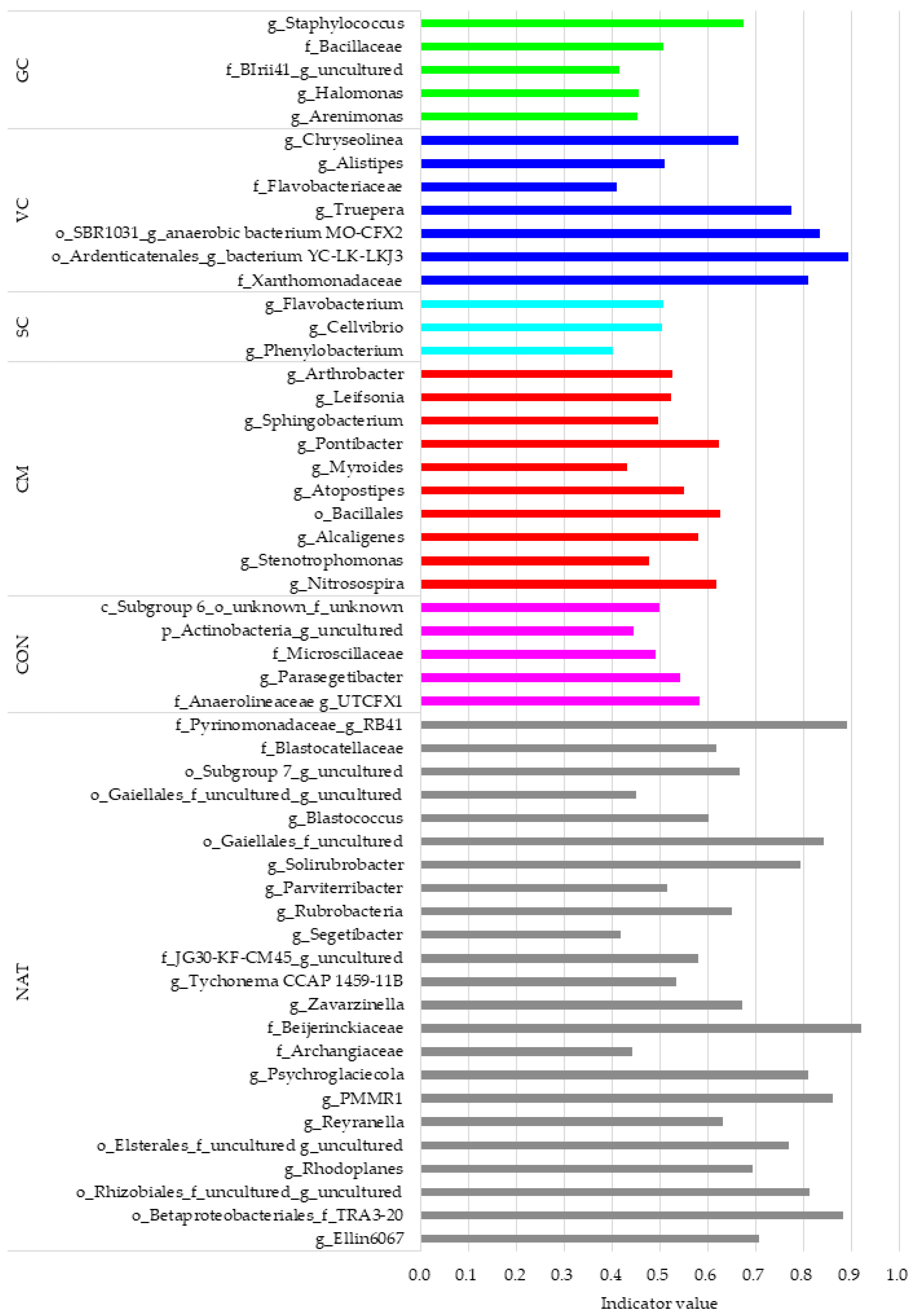
3.4. Indicator Bacterial Communities in Different Restored, Control, and Reference Soils
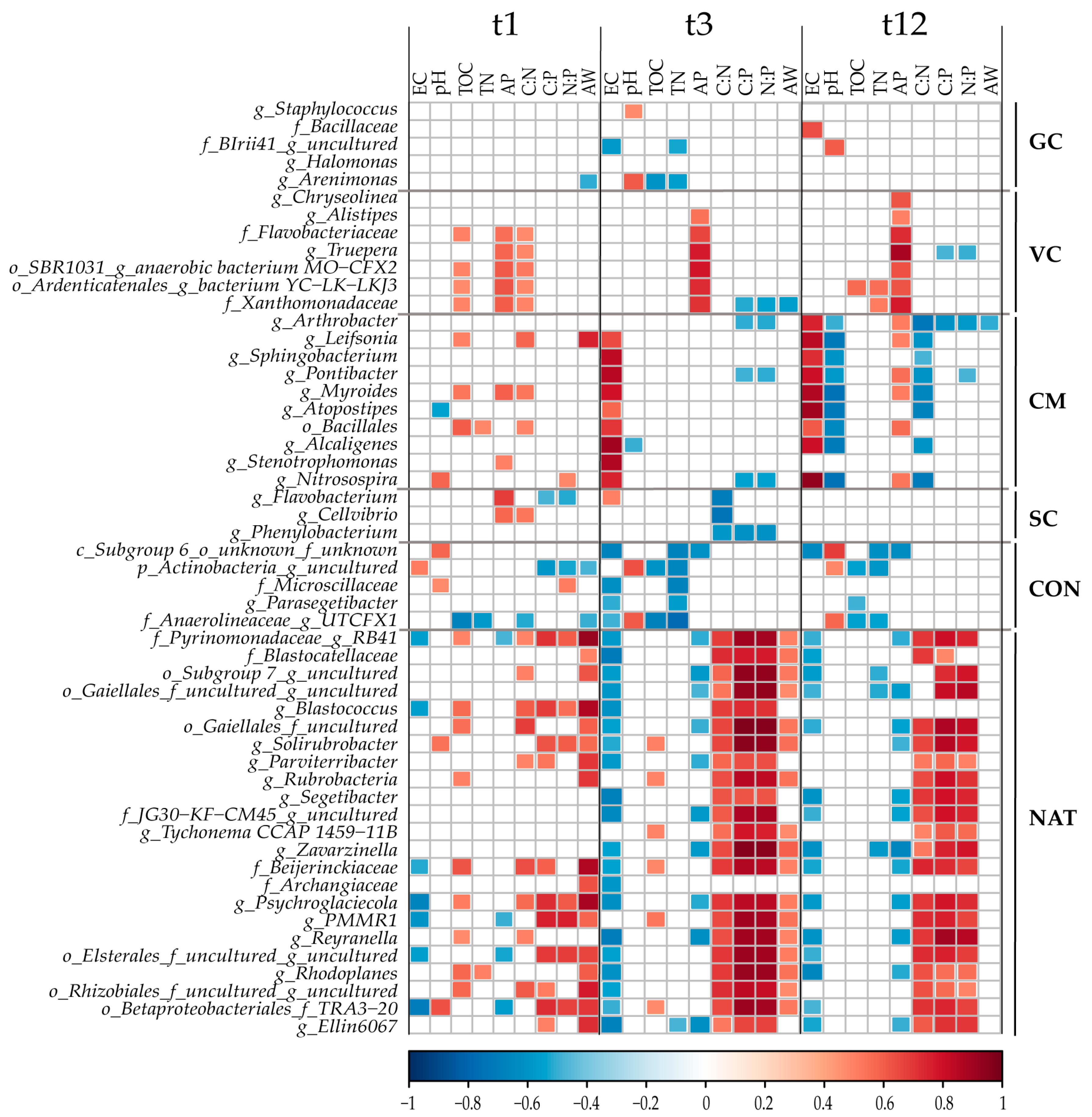
3.5. Correlation Analysis between Bacterial Taxa from Indicspecies Analysis and Soil Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stanturf, J.A. Landscape Degradation and Restoration. In Soils and Landscape Restoration; Elsevier: Amsterdam, The Netherlands, 2021; pp. 125–159. [Google Scholar]
- Navarro, J.A.C.; Estellés, M.G.; González, G.G.; Sánchez, V.M.C.; del Campo, M.V. Restauración Ecológica En Ambientes Semiáridos. Recuperar Las Interacciónes Biológicas y Las Funciones Ecosistémicas; CSIC: Madrid, Spain, 2017.
- Bastida, F.; Torres, I.F.; Hernández, T.; Bombach, P.; Richnow, H.H.; García, C. Can the Labile Carbon Contribute to Carbon Immobilization in Semiarid Soils? Priming Effects and Microbial Community Dynamics. Soil Biol. Biochem. 2013, 57, 892–902. [Google Scholar] [CrossRef]
- García, C.; Hernández, T.; Costa, F. Microbial Activity in Soils under Mediterranean Environmental Conditions. Soil Biol. Biochem. 1994, 26, 1185–1191. [Google Scholar] [CrossRef]
- Lasanta, T.; Arnáez, J.; Nadal-Romero, E. Soil Degradation, Restoration and Management in Abandoned and Afforested Lands; Elsevier: Amsterdam, The Netherlands, 2019; Volume 4, pp. 71–117. [Google Scholar]
- Li, Y.; Gong, J.; Liu, J.; Hou, W.; Moroenyane, I.; Liu, Y.; Jin, J.; Liu, J.; Xiong, H.; Cheng, C.; et al. Effects of Different Land Use Types and Soil Depth on Soil Nutrients and Soil Bacterial Communities in a Karst Area, Southwest China. Soil Syst. 2022, 6, 20. [Google Scholar] [CrossRef]
- Gao, W.; Xu, J.; Zhao, J.; Zhang, H.; Ni, Y.; Zhao, B.; Tebbe, C.C.; Zhang, J.; Jia, Z. Prokaryotic Community Assembly after 40 Years of Soda Solonetz Restoration by Natural Grassland and Reclaimed Farmland. Eur. J. Soil Biol. 2020, 100, 103213. [Google Scholar] [CrossRef]
- Calleja-Cervantes, M.E.; Menéndez, S.; Fernández-González, A.J.; Irigoyen, I.; Cibriáin-Sabalza, J.F.; Toro, N.; Aparicio-Tejo, P.M.; Fernández-López, M. Changes in Soil Nutrient Content and Bacterial Community after 12 Years of Organic Amendment Application to a Vineyard. Eur. J. Soil Sci. 2015, 66, 802–812. [Google Scholar] [CrossRef]
- Wijesekara, H.; Bolan, N.S.; Vithanage, M.; Xu, Y.; Mandal, S.; Brown, S.L.; Hettiarachchi, G.M.; Pierzynski, G.M.; Huang, L.; Ok, Y.S.; et al. Utilization of Biowaste for Mine Spoil Rehabilitation. Adv. Agron. 2016, 138, 97–173. [Google Scholar] [CrossRef]
- Rodríguez-Berbel, N.; Ortega, R.; Lucas-Borja, M.E.; Solé-Benet, A.; Miralles, I. Long-Term Effects of Two Organic Amendments on Bacterial Communities of Calcareous Mediterranean Soils Degraded by Mining. J. Environ. Manag. 2020, 271, 110920. [Google Scholar] [CrossRef]
- Rodríguez-Berbel, N.; Soria, R.; Ortega, R.; Bastida, F.; Miralles, I. Quarry Restoration Treatments from Recycled Waste Modify the Physicochemical Soil Properties, Composition and Activity of Bacterial Communities and Priming Effect in Semi-Arid Areas. Sci. Total Environ. 2021, 774, 145693. [Google Scholar] [CrossRef]
- Soria, R.; González-Pérez, J.A.; de la Rosa, J.M.; San Emeterio, L.M.; Domene, M.A.; Ortega, R.; Miralles, I. Effects of Technosols Based on Organic Amendments Addition for the Recovery of the Functionality of Degraded Quarry Soils under Semiarid Mediterranean Climate: A Field Study. Sci. Total Environ. 2022, 816, 151572. [Google Scholar] [CrossRef]
- Bastida, F.; Torres, I.F.; Hernández, T.; García, C. The Impacts of Organic Amendments: Do They Confer Stability against Drought on the Soil Microbial Community? Soil Biol. Biochem. 2017, 113, 173–183. [Google Scholar] [CrossRef]
- Tejada, M.; Hernandez, M.T.; Garcia, C. Application of Two Organic Amendments on Soil Restoration: Effects on the Soil Biological Properties. J. Environ. Qual. 2006, 35, 1010–1017. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhu, W.; Xu, F.; Du, J.; Tian, X.; Shi, J.; Wei, G. Organic Amendments Affect Soil Organic Carbon Sequestration and Fractions in Fields with Long-Term Contrasting Nitrogen Applications. Agric. Ecosyst. Environ. 2021, 322, 107643. [Google Scholar] [CrossRef]
- Wang, Q.; Gao, W.; Bol, R.; Xiao, Q.; Wu, L.; Zhang, W. Microbial Regulation of Net N Mineralisation Is Driven by C, N, P Content and Stoichiometry. Eur. J. Soil Sci. 2022, 73, e13257. [Google Scholar] [CrossRef]
- Xiao, C.; Janssens, I.A.; Zhou, Y.; Su, J.; Liang, Y.; Guenet, B. Strong Stoichiometric Resilience after Litter Manipulation Experiments; A Case Study in a Chinese Grassland. Biogeosciences 2015, 12, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Bastida, F.; Kandeler, E.; Moreno, J.L.; Ros, M.; García, C.; Hernández, T. Application of Fresh and Composted Organic Wastes Modifies Structure, Size and Activity of Soil Microbial Community under Semiarid Climate. Appl. Soil Ecol. 2008, 40, 318–329. [Google Scholar] [CrossRef]
- Xue, L.; Ren, H.; Brodribb, T.J.; Wang, J.; Yao, X.; Li, S. Long Term Effects of Management Practice Intensification on Soil Microbial Community Structure and Co-Occurrence Network in a Non-Timber Plantation. For. Ecol. Manag. 2020, 459, 117805. [Google Scholar] [CrossRef]
- Singh, D.P.; Prabha, R. Microbial Interventions in Agriculture and Environment; Springer: Singapore, 2019; Volume 3, ISBN 9789813290846. [Google Scholar]
- Li, F.; Chen, L.; Zhang, J.; Yin, J.; Huang, S. Bacterial Community Structure after Long-Term Organic and Inorganic Fertilization Reveals Important Associations between Soil Nutrients and Specific Taxa Involved in Nutrient Transformations. Front. Microbiol. 2017, 8, 187. [Google Scholar] [CrossRef] [Green Version]
- Neemisha; Sharma, S. Soil Enzymes and Their Role in Nutrient Cycling. Struct. Funct. Pedosph. 2022, 173–188. [Google Scholar] [CrossRef]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. Organic Amendments: Microbial Community Structure, Activity and Abundance of Agriculturally Relevant Microbes Are Driven by Long-Term Fertilization Strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [Green Version]
- Peinado, F.M.; Sierra, A.M. Proyecto LUCDEME. Mapa de Suelos a Escala 1:100.000 Provincia de Almería 2004. Available online: https://www.miteco.gob.es/es/biodiversidad/temas/desertificacion-restauracion/memoriamapasueloslucdemealmeria_tcm30-512287.pdf (accessed on 3 November 2022).
- SAIH Hidrosur Demarcación Hidrográfica de Las Cuencas Mediterráneas Andaluzas. Informe Hidrológico 2020–2021. Consejería de Agricultura, Ganadería, Pesca y Desarrollo Sostenible; Dirección General de Infraestructuras Del Agua: Sevilla, Spain, 2021. [Google Scholar]
- Wongkiew, S.; Koottatep, T.; Polprasert, C.; Prombutara, P.; Jinsart, W.; Khanal, S.K. Bioponic System for Nitrogen and Phosphorus Recovery from Chicken Manure: Evaluation of Manure Loading and Microbial Communities. Waste Manag. 2021, 125, 67–76. [Google Scholar] [CrossRef]
- Munoz-Ucros, J.; Panke-Buisse, K.; Robe, J. Bacterial Community Composition of Vermicompost-Treated Tomato Rhizospheres. PLoS ONE 2020, 15, e0230577. [Google Scholar] [CrossRef] [PubMed]
- Naseer, I.; Ahmad, M.; Nadeem, S.M.; Ahmad, I.; Najm-ul-Seher; Zahir, Z.A. Biofertilizers for Sustainable Agriculture and Environment; Springer: Cham, Switzerland, 2019; Volume 55, ISBN 978-3-030-18932-7. [Google Scholar]
- Gupta, A.K.; Tomar, J.M.S.; Kaushal, R.; Kadam, D.M.; Rathore, A.C.; Mehta, H.; Ojasvi, P.R. Aromatic Plants Based Environmental Sustainability with Special Reference to Degraded Land Management. J. Appl. Res. Med. Aromat. Plants 2021, 22, 100298. [Google Scholar] [CrossRef]
- Costa Pérez, J.C.; Valle Tendero, F. Modelos de Restauración Forestal: Datos Botánicos Aplicados a La Gestión Del Medio Ambiente Andaluz; Consejería de Medio Ambiente; Junta de Andalucía, Consejería de Medio Ambiente: Sevilla, Spain, 2004; ISBN 84-95785-97-8. [Google Scholar]
- Miralles, I.; Ortega, R.; Almendros, G.; Sánchez-Marañón, M.; Soriano, M. Soil Quality and Organic Carbon Ratios in Mountain Agroecosystems of South-East Spain. Geoderma 2009, 150, 120–128. [Google Scholar] [CrossRef]
- Mingorance, M.D.; Barahona, E.; Fernández-Gálvez, J. Guidelines for Improving Organic Carbon Recovery by the Wet Oxidation Method. Chemosphere 2007, 68, 409–413. [Google Scholar] [CrossRef]
- Olsen, S.R.; Watanabe, F.S. A Method to Determine a Phosphorus Adsorption Maximum of Soils as Measured by the Langmuir Isotherm. Soil Sci. Soc. Am. J. 1957, 21, 144–149. [Google Scholar] [CrossRef]
- Boudjabi, S.; Chenchouni, H. Soil Fertility Indicators and Soil Stoichiometry in Semi-Arid Steppe Rangelands. CATENA 2022, 210, 105910. [Google Scholar] [CrossRef]
- Richards, A. Pressure-Membrane Extraction Apparatus for Soil Solution. Soil Sci. 1941, 51, 377–386. [Google Scholar] [CrossRef]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium; US Department of Agriculture: Washington, DC, USA, 1954.
- Comeau, A.M.; Douglas, G.M.; Langille, M.G.I. Microbiome Helper: A Custom and Streamlined Workflow for Microbiome Research. mSystems 2017, 2, e00127-16. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef Statistics Reference Online; Wiley: New York, NY, USA, 2017; pp. 1–15. [Google Scholar] [CrossRef]
- Akaike, H. A New Look at the Statistical Model Identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- De Caceres, M.; Jansen, F. Package “indicspecies” Type Package Title Relationship Between Species and Groups of Sites. Version 1.7.6. Available online: https://cran.r-project.org/web/packages/indicspecies/index.html (accessed on 26 September 2022).
- Lv, X.-M.; Shao, M.-F.; Li, C.-L.; Li, J.; Gao, X.; Sun, F.-Y. A Comparative Study of the Bacterial Community in Denitrifying and Traditional Enhanced Biological Phosphorus Removal Processes. Microbes Environ. 2014, 29, 261–268. [Google Scholar] [CrossRef] [Green Version]
- González-Ubierna, S.; Jorge-Mardomingo, I.; Carrero-González, B.; de la Cruz, M.T.; Casermeiro, M.Á. Soil Organic Matter Evolution after the Application of High Doses of Organic Amendments in a Mediterranean Calcareous Soil. J. Soils Sediments 2012, 12, 1257–1268. [Google Scholar] [CrossRef]
- Li, J.; Cooper, J.M.; Lin, Z.; Li, Y.; Yang, X.; Zhao, B. Soil Microbial Community Structure and Function Are Significantly Affected by Long-Term Organic and Mineral Fertilization Regimes in the North China Plain. Appl. Soil Ecol. 2015, 96, 75–87. [Google Scholar] [CrossRef]
- Khan, K.S.; Mack, R.; Castillo, X.; Kaiser, M.; Joergensen, R.G. Microbial Biomass, Fungal and Bacterial Residues, and Their Relationships to the Soil Organic Matter C/N/P/S Ratios. Geoderma 2016, 271, 115–123. [Google Scholar] [CrossRef]
- Kadoglidou, K.; Chalkos, D.; Karamanoli, K.; Eleftherohorinos, I.G.; Constantinidou, H.I.A.; Vokou, D. Aromatic Plants as Soil Amendments: Effects of Spearmint and Sage on Soil Properties, Growth and Physiology of Tomato Seedlings. Sci. Hortic. 2014, 179, 25–35. [Google Scholar] [CrossRef]
- Zheng, X.; Aborisde, M.A.; Wang, H.; He, P.; Lu, S.; Cui, N.; Wang, S.; Zhang, H.; Ding, H.; Liu, K. Effect of Lignin and Plant Growth-Promoting Bacteria (Staphylococcus Pasteuri) on Microbe-Plant Co-Remediation: A PAHs-DDTs Co-Contaminated Agricultural Greenhouse Study. Chemosphere 2020, 256, 127079. [Google Scholar] [CrossRef]
- Pratap, D.; Harikesh, S.; Singh, B.; Prabha, R. Microbial Inoculants in Sustainable Agricultural Productivity; Springer: Berlin/Heidelberg, Germany, 2016; Volume 1, ISBN 9788132226451. [Google Scholar]
- Ortega, R.; Miralles, I.; Domene, M.A.; Soriano, M. Functions and Applications of Plant Growth Promoting Bacteria (PGPR) in Highly Technified Crops. Agric. Res. Technol. 2017, 12, 10–13. [Google Scholar]
- Fierer, N. Embracing the Unknown: Disentangling the Complexities of the Soil Microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Goldfarb, K.C.; Karaoz, U.; Hanson, C.A.; Santee, C.A.; Bradford, M.A.; Treseder, K.K.; Wallenstein, M.D.; Brodie, E.L. Differential Growth Responses of Soil Bacterial Taxa to Carbon Substrates of Varying Chemical Recalcitrance. Front. Microbiol. 2011, 2, 94. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Zhou, X.; Li, Y.; Li, K.; Wang, C.; Liu, J.; Yan, D.; Liu, Y.; Yang, D.; Xing, J. Heterotrophic Nitrification and Aerobic Denitrification by a Novel Halomonas Campisalis. Biotechnol. Lett. 2013, 35, 2045–2049. [Google Scholar] [CrossRef]
- Xu, J.; Xu, X.; Han, Y.; Sheng, S.; Meng, Q.; Wang, Z.; Liu, W.; Di, L. Evaluation of a Novel Thermophilic Nitrifying Bacillaceae Species Aliibacillus Thermotolerans BM62T Promoting Nitrogen Retention in Livestock Manure Compost. Environ. Prog. Sustain. Energy 2019, 39, e13357. [Google Scholar] [CrossRef]
- Neher, D.A.; Limoges, M.A.; Weicht, T.R.; Sharma, M.; Millner, P.D.; Donnelly, C. Bacterial Community Dynamics Distinguish Poultry Compost from Dairy Compost and Non-Amended Soils Planted with Spinach. Microorganism 2020, 8, 1601. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yu, C.; Wang, X.; Hai, L.; Hu, J. Increased Abundance of Nitrogen Fixing Bacteria by Higher C/N Ratio Reduces the Total Losses of N and C in Cattle Manure and Corn Stover Mix Composting. Waste Manag. 2020, 103, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Akyol, Ç.; Ince, O.; Ince, B. Crop-Based Composting of Lignocellulosic Digestates: Focus on Bacterial and Fungal Diversity. Bioresour. Technol. 2019, 288, 121549. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Carrazco, A.; Chávez-López, C.; Ramírez-Villanueva, D.A.; Dendooven, L. Bacteria in (Vermi)Composted Organic Wastes Mostly Survive When Applied to an Arable Soil Cultivated with Wheat (Triticum Sp. L.). Environ. Monit. Assess. 2022, 194, 363. [Google Scholar] [CrossRef] [PubMed]
- Miralles, I.; Soria, R.; Lucas-Borja, M.E.; Soriano, M.; Ortega, R. Effect of Biocrusts on Bacterial Community Composition at Different Soil Depths in Mediterranean Semi-Arid Ecosystems. Sci. Total Environ. 2020, 733, 138613. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Wang, X.; Zhang, N.; Li, B. Biogeographical Patterns and Assembly of Bacterial Communities in Saline Soils of Northeast China. Microorganisms 2022, 10, 1787. [Google Scholar] [CrossRef]
- Miralles, I.; Ortega, R.; Montero-Calasanz, M.C. Studying the Microbiome of Cyanobacterial Biocrusts from Drylands and Its Functional Inuence on Biogeochemical Cycles. 2021. Available online: https://www.researchgate.net/publication/349600353_Studying_the_Microbiome_of_Cyanobacterial_Biocrusts_From_Drylands_and_Its_Functional_Influence_on_Biogeochemical_Cycles (accessed on 3 November 2022).
- Ye, G.; Lin, Y.; Liu, D.; Chen, Z.; Luo, J.; Bolan, N.; Fan, J.; Ding, W. Long-Term Application of Manure over Plant Residues Mitigates Acidification, Builds Soil Organic Carbon and Shifts Prokaryotic Diversity in Acidic Ultisols. Appl. Soil Ecol. 2019, 133, 24–33. [Google Scholar] [CrossRef]
- Lin, Y.; Ye, G.; Kuzyakov, Y.; Liu, D.; Fan, J.; Ding, W. Long-Term Manure Application Increases Soil Organic Matter and Aggregation, and Alters Microbial Community Structure and Keystone Taxa. Soil Biol. Biochem. 2019, 134, 187–196. [Google Scholar] [CrossRef]
- McBride, M.J. The Family Flavobacteriaceae. In The Prokaryotes: Other Major Lineages of Bacteria and The Archaea; Springer: Berlin/Heidelberg, Germany, 2014; Volume 9783642389, pp. 643–676. ISBN 9783642301230. [Google Scholar]
- Elbl, J.; Maková, J.; Javoreková, S.; Medo, J.; Kintl, A.; Lošák, T.; Lukas, V. Response of Microbial Activities in Soil to Various Organic and Mineral Amendments as an Indicator of Soil Quality. Agronomy 2019, 9, 485. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an Ecological Classification of Soil Bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Zhang, C.; Li, J.; Wang, J.; Liu, G.; Wang, G.; Guo, L.; Peng, S. Decreased Temporary Turnover of Bacterial Communities along Soil Depth Gradient during a 35-Year Grazing Exclusion Period in a Semiarid Grassland. Geoderma 2019, 351, 49–58. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GC | VC | SC | CM | CON | NAT | ||
|---|---|---|---|---|---|---|---|
| t1 | EC (mS cm−1) | 0.98 ± 0.01 aA | 0.63 ± 0.8 bA | 0.93 ± 0.09 abA | 1.02 ± 0.11 aA | 0.10 ± 0.00 cA | 0.09 ± 0.00 cA |
| pH | 8.13 ± 0.04 abA | 8.25 ± 0.01 acA | 8.07 ± 0.02 bA | 8.01 ± 0.01 bA | 8.44 ± 0.08 cAB | 8.31 ± 0.07 acA | |
| TOC (%) | 2.55 ± 0.24 aA | 4.01 ± 0.55 aA | 2.81 ± 0.30 aAB | 2.80 ± 0.13 aA | 1.78 ± 0.12 bA | 4.25 ± 0.59 aA | |
| TN (%) | 0.29 ± 0.02 aA | 0.39 ± 0.02 bA | 0.367 ± 0.04 abA | 0.370 ± 0.01 bA | 0.210 ± 0.01 cA | 0.393 ± 0.04 abA | |
| AP (%) | 0.004 ± 0.00 aA | 0.028 ± 0.0 b0A | 0.019 ± 0.00 bA | 0.023 ± 0.00 bA | 0.002 ± 0.00 aA | 0.004 ± 0.00 aA | |
| C:N | 8.63 ± 0.45 aA | 10.25 ± 0.86 bA | 7.67 ± 0.25 abA | 7.56 ± 0.19 bA | 8.52 ± 0.20 cA | 10.78 ± 0.48 abA | |
| C:P | 581.75 ± 148.35 aA | 141.75 ± 5.77 bA | 141.84 ± 10.34 bA | 120.35 ± 8.87 bA | 730.80 ± 132.85 aA | 1022.07 ± 100.00 aA | |
| N:P | 66.15 ± 13.31 aA | 13.95 ± 0.82 bA | 18.54 ± 1.65 bA | 15.87 ± 0.79 bA | 86.57 ± 17.95 aA | 95.69 ± 12.89 aA | |
| AW (%) | 9.66 ± 0.26 aA | 9.19 ± 0.39 aA | 9.52 ± 0.24 aA | 9.47 ± 0.30 aA | 9.12 ± 0.37 aA | 13.02 ± 1.01 bA | |
| t3 | EC (mS cm−1) | 0.20 ± 0.01 aAB | 0.24± 0.01 abB | 0.43 ± 0.06 bB | 0.72 ± 0.16 bA | 0.09 ± 0.01 cA | 0.07 ± 0.01 cA |
| pH | 8.38 ± 0.14 abcA | 8.35 ± 0.05 aA | 8.04 ± 0.04 bA | 8.05 ± 0.08 bcB | 8.34 ± 0.07 acB | 8.03 ± 0.05 bB | |
| TOC (%) | 2.17 ± 0.23 abA | 3.20 ± 0.13 cA | 2.48 ± 0.04 aA | 2.74 ± 0.27 acAB | 1.64 ± 0.18 bAB | 3.44 ± 0.14 cA | |
| TN (%) | 0.27 ± 0.02 aA | 0.35 ± 0.00 bA | 0.34 ± 0.01 bA | 0.32 ± 0.04 abA | 0.14 ± 0.01 cA | 0.33 ± 0.01 abA | |
| AP (%) | 0.015 ± 0.00 aB | 0.12 ± 0.00 bB | 0.05 ± 0.00 cB | 0.07 ± 0.02 abcB | 0.007 ± 0.00 dA | 0.004 ± 0.00 dA | |
| C:N | 8.12 ± 0.43 aA | 9.15 ± 0.23 bA | 7.29 ± 0.38 bA | 8.68 ± 0.54 abA | 8.84 ± 0.43 cA | 10.36 ± 0.03 abA | |
| C:P | 139.91 ± 3.24 aB | 26.58 ± 0.96 bB | 54.05 ± 7.14 cB | 42.63 ± 9.13 bc B | 246.07 ± 40.96 aB | 840.35 ± 184.24 dA | |
| N:P | 17.3 ± 1.04 aB | 2.90 ± 0.16 bB | 7.35 ± 0.58 cB | 4.90 ± 1.03 bcB | 28.43 ± 6.31 aB | 81.18 ± 18.03 dA | |
| AW (%) | 10.70 ± 0.36 aA | 8.42 ± 2.13 abA | 9.65 ± 0.41 aA | 10.08 ± 0.27 aA | 10.43 ± 1.68 abA | 13.12 ± 0.24 bA | |
| t12 | EC (mS cm−1) | 0.32 ± 0.06 aB | 0.45 ± 0.08 abA | 0.47 ± 0.04 aB | 1.30 ± 0.30 bA | 0.10 ± 0.01 cA | 0.08 ± 0.00 cA |
| pH | 8.76 ± 0.16 aB | 8.55 ± 0.14 abB | 8.11 ± 0.00 cA | 7.67 ± 0.11 dC | 8.54 ± 0.01 aA | 8.19 ± 0.02 bAB | |
| TOC (%) | 2.61 ± 0.22 abA | 3.47 ± 0.27 aA | 2.86 ± 0.05 aB | 2.23 ± 0.12 bB | 1.31 ± 0.05 cB | 3.03 ± 0.03 aA | |
| TN (%) | 0.27 ± 0.01 aA | 0.39 ± 0.01 bA | 0.34 ± 0.01 bA | 0.33 ± 0.04 abA | 0.16 ± 0.00 cA | 0.28 ± 0.00 aA | |
| AP (%) | 0.01 ± 0.00 aB | 0.158 ± 0.02 bB | 0.055 ± 0.00 cB | 0.141 ± 0.02 bB | 0.003 ± 0.00 dA | 0.004 ± 0.00 dA | |
| C:N | 9.51 ± 0.29 aA | 8.70 ± 0.36 bA | 8.42 ± 0.27 bA | 6.86 ± 0.66 abA | 8.14 ± 0.26 cA | 10.7 ± 0.37 aA | |
| C:P | 214.96 ± 41.2 aAB | 23.22 ± 4.40 bB | 51.78 ± 4.08 cB | 16.42 ± 2.43 bC | 457.89 ± 141.61 adAB | 728.85 ± 133.25 dA | |
| N:P | 22.36 ± 3.75 aB | 2.63 ± 0.42 bB | 6.13 ± 0.29 cB | 2.48 ± 0.52 bB | 55.19 ± 15.86 adAB | 67.34 ± 11.16 dA | |
| AW (%) | 10.63 ± 1.59 abA | 8.91 ± 1.33 abA | 8.42 ± 1.23 abAB | 7.37 ± 0.67 aB | 8.49 ± 1.17 abA | 11.02 ± 0.07 bA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Berbel, N.; Soria, R.; Villafuerte, A.B.; Ortega, R.; Miralles, I. Short-Term Dynamics of Bacterial Community Structure in Restored Abandoned Agricultural Soils under Semi-Arid Conditions. Agronomy 2023, 13, 86. https://doi.org/10.3390/agronomy13010086
Rodríguez-Berbel N, Soria R, Villafuerte AB, Ortega R, Miralles I. Short-Term Dynamics of Bacterial Community Structure in Restored Abandoned Agricultural Soils under Semi-Arid Conditions. Agronomy. 2023; 13(1):86. https://doi.org/10.3390/agronomy13010086
Chicago/Turabian StyleRodríguez-Berbel, Natalia, Rocío Soria, Ana B. Villafuerte, Raúl Ortega, and Isabel Miralles. 2023. "Short-Term Dynamics of Bacterial Community Structure in Restored Abandoned Agricultural Soils under Semi-Arid Conditions" Agronomy 13, no. 1: 86. https://doi.org/10.3390/agronomy13010086