

Insect Pollinator Monitoring in and around a Netted Plot of Apple Trees—Biosafety Implications for Genetically Engineered Fruit Trees
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Netted Plot
2.2. Pan Trap Sampling and Arthropod Identification
2.3. Flower Visitation Observations
2.4. Hand Collections
2.5. Data Analysis
3. Results
3.1. Arthropods Collected inside and outside the Netted Plot
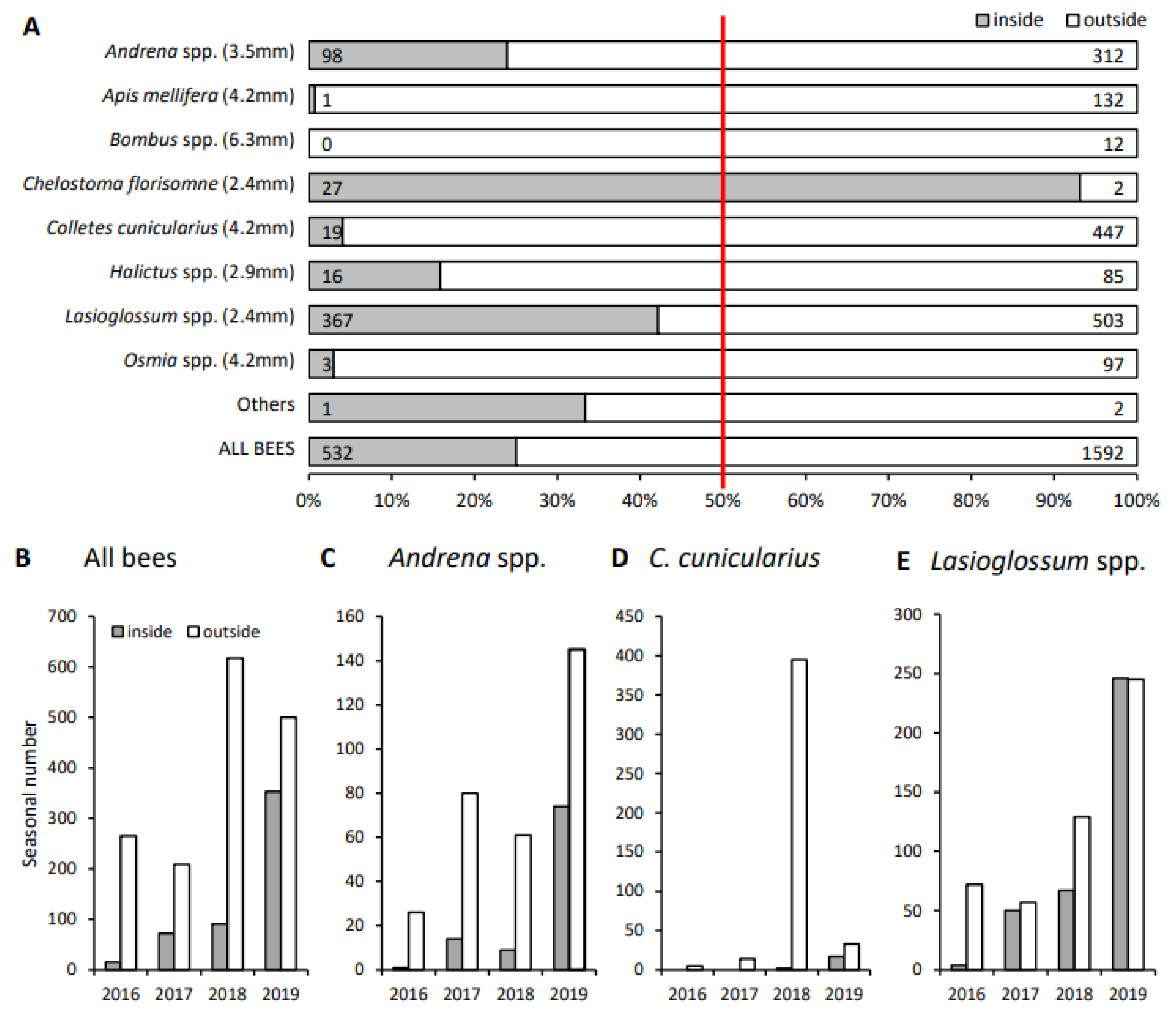
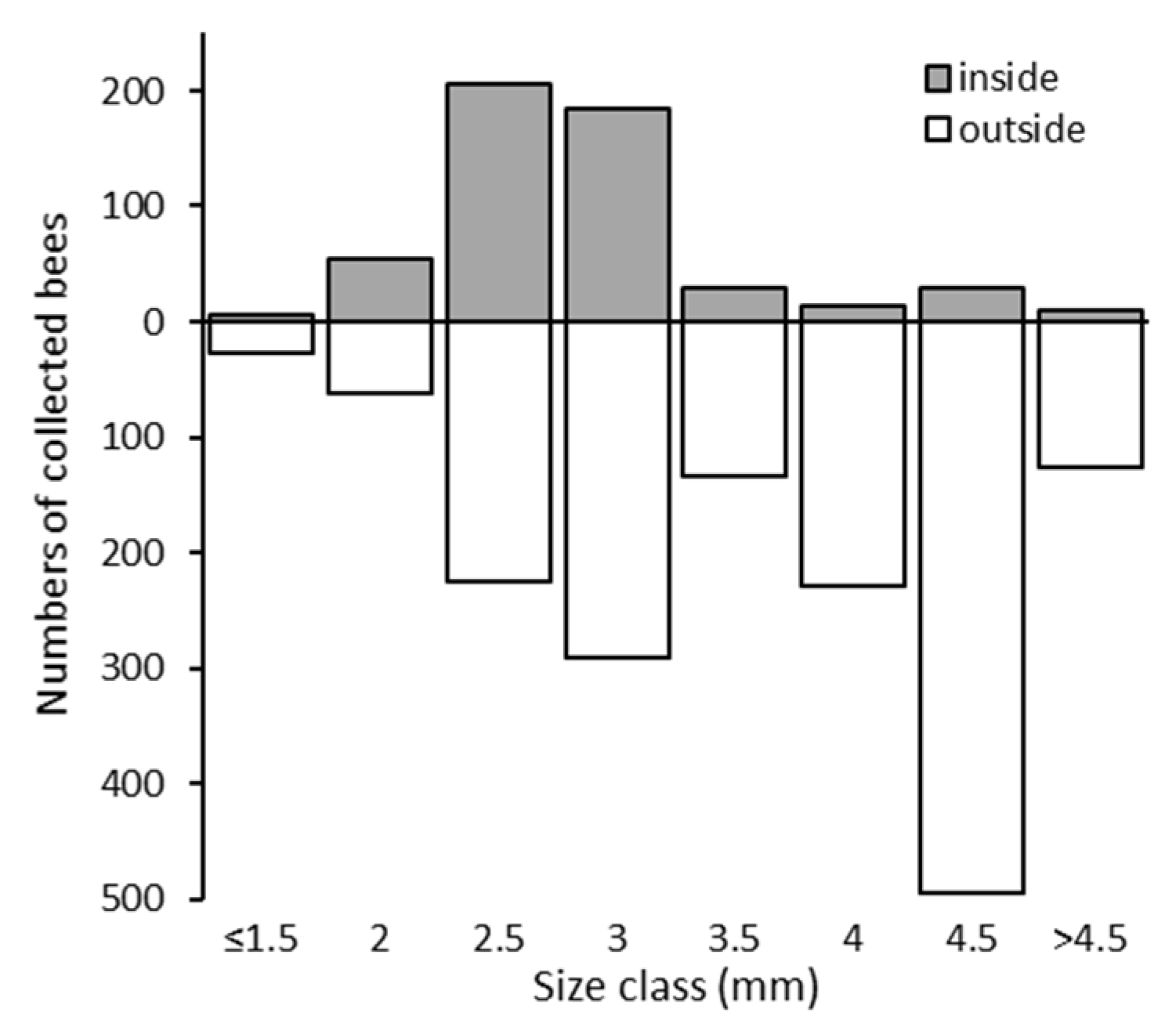
3.2. Bees Collected inside and outside the Netted Plot
3.3. Bees Collected before the Net Was Closed
3.4. Capability of Bees to Pass the Hail Net
3.5. Flower Visitation Observations
3.6. Hand Collections
4. Discussion
4.1. Fewer Arthropods in Netted Plot
4.2. Netted Plot Provides Habitat for Arthropods
4.3. Evidence for Pollen Flow through the Netting
4.4. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanblaere, T.; Szankowski, I.; Schaart, J.; Schouten, H.; Flachowsky, H.; Broggini, G.A.L.; Gessler, C. The development of a cisgenic apple plant. J. Biotechnol. 2011, 154, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Kost, T.D.; Gessler, C.; Jänsch, M.; Flachowsky, H.; Patocchi, A.; Broggini, G.A.L. Development of the first cisgenic apple with 441 increased resistance to fire blight. PLoS ONE 2015, 10, e0143980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. Royal Soc. B 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, F.; Davenport, T.L. Apple pollination: A review. Sci. Hortic. 2013, 162, 188–203. [Google Scholar] [CrossRef]
- Pardo, A.; Borges, P.A.V. Worldwide importance of insect pollination in apple orchards: A review. Agric. Ecosyst. Environ. 2020, 293, 106839. [Google Scholar] [CrossRef]
- Roquer-Beni, L.; Alins, G.; Arnan, X.; Boreux, V.; García, D.; Hambäck, P.A.; Happe, A.K.; Klein, A.M.; Miñarro, M.; Mody, K.; et al. Management-dependent effects of pollinator functional diversity on apple pollination services: A response–effect trait approach. J. Appl. Ecol. 2021, 58, 2843–2853. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Breeze, T.D.; Boreux, V.; Fountain, M.T.; McKerchar, M.; Webber, S.M.; Coston, D.J.; Jenner, N.; Dean, R.; Westbury, D.B.; et al. Apple pollination: Demand depends on variety and supply depends on pollinator identity. PloS ONE 2016, 11, e0153889. [Google Scholar] [CrossRef] [Green Version]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee foraging ranges and their relationship to body size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef]
- Zurbuchen, A.; Landert, L.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Maximum foraging ranges in solitary bees: Only few individuals have the capability to cover long foraging distances. Biol. Conserv. 2010, 143, 669–676. [Google Scholar] [CrossRef]
- Federal Assembly of the Swiss Confederation. Federal Act of 21 March 2003 on Non-Human Gene Technology (Gene Technology Act, GTA). SR 814.91. 2003. Available online: https://www.fedlex.admin.ch/eli/cc/2003/705/en (accessed on 26 December 2022).
- European Parliament. Directive 2001/18/EC of the European Parliament and of the Council of 12 March 2001 on the deliberate release into the environment of genetically modified organisms and repealing Council Directive 90/220/EEC. 2001. Available online: http://data.europa.eu/eli/dir/2001/18/2021-03-27 (accessed on 26 December 2022).
- Chouinard, G.; Firlej, A.; Cormier, D. Going beyond sprays and killing agents: Exclusion, sterilization and disruption for insect pest control in pome and stone fruit orchards. Sci. Hortic. 2016, 208, 13–27. [Google Scholar] [CrossRef]
- Zwahlen, D.; Kuster, T.; Kuske, S. Insektennetz im Apfelanbau: Kaum Auswirkungen auf Mikroklima und Fruchtqualität. Schweiz. Z. Für Obs.-Und Weinbau 2017, 5, 8–11. [Google Scholar]
- Schlathölter, I.; Dalbosco, A.; Meissle, M.; Knauf, A.; Dallemulle, A.; Keller, B.; Romeis, J.; Broggini, G.A.L.; Patocchi, A. Low outcrossing from an apple field trial protected with nets. Agronomy 2021, 11, 1754. [Google Scholar] [CrossRef]
- Bosch, J.; Blas, M. Foraging behaviour and pollinating efficiency of Osmia cornuta and Apis mellifera on almond (Hymenoptera, Megachilidae and Apidae). Appl. Entomol. Zool. 1994, 29, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Monzón, V.H.; Bosch, J.; Retana, J. Foraging behavior and pollinating effectiveness of Osmia cornuta (Hymenoptera: Megachilidae) and Apis mellifera (Hymenoptera: Apidae) on “Comice” pear. Apidologie 2004, 35, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.W.; Hanula, J.L. Efficiency of Malaise traps and colored pan traps for collecting flower-visiting insects from three forested ecosystems. J. Insect Conserv. 2007, 11, 399–408. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different European habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.-M.; Brittain, C.; Hendrix, S.D.; Thorp, R.; Williams, N.; Kremen, C. Wild pollination services to California almond rely on semi-natural habitat. J. Appl. Ecol. 2012, 49, 723–732. [Google Scholar] [CrossRef]
- Vrdoljak, S.M.; Samways, M.J. Optimising coloured pan traps to survey flower visiting insects. J Insect Conserv 2012, 16, 345–354. [Google Scholar] [CrossRef]
- Schlathölter, I.; Meissle, M.; Boeriis, T.; Heimo, D.; Studer, B.; Broggini, G.A.L.; Romeis, J.; Patocchi, A. No adverse dietary effect of a cisgenic fire blight resistant apple line on the non-target arthropods Drosophila melanogaster and Folsomia candida. Ecotoxicol. Env. Saf. 2022, 241, 113749. [Google Scholar] [CrossRef]
- Southwood, T.R.E. Ecological Methods–With Particular Reference to the Study of Insect Populations, ELBS ed.; University Printing House: Cambridge, UK, 1978. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 26 December 2022).
- Candian, V.; Pansa, M.G.; Santoro, K.; Spadaro, D.; Tavella, L.; Tedeschi, R. Photoselective exclusion netting in apple orchards: Effectiveness against pests and impact on beneficial arthropods, fungal diseases and fruit quality. Pest Manag. Sci. 2020, 76, 179–187. [Google Scholar] [CrossRef]
- Schluchter, M.; Späth, S.; Buchleither, S. Four-year experiences with exclusion netting row covers in an organic apple orchard: Handling, relevant pests and diseases. In Proceedings of the 20th International Conference on Organic Fruit-Growing, Online, 21–23 February 2022; pp. 94–100. [Google Scholar]
- Rigden, P. To Net or Not to Net, 3rd ed. The State of Queensland, Department of Primary Industries and Fisheries. 2008. Available online: https://www.daf.qld.gov.au/__data/assets/pdf_file/0009/72954/Orchard-Netting-Report.pdf (accessed on 26 December 2022).
- Mupambi, G.; Anthony, B.M.; Layne, D.R.; Musacchi, S.; Serra, S.; Schmidt, T.; Kalcsits, L.A. The influence of protective netting on tree physiology and fruit quality of apple: A review. Sci. Hortic. 2018, 236, 60–72. [Google Scholar] [CrossRef]
- Bogo, A.; Trezzi Casa, R.; Rufato, L.; Gonçalves, M.J. The effect of hail protection nets on Glomerella leaf spot in ‘royal Gala’ apple. Crop Prot. 2012, 31, 40–44. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Date Nets Closed | Start Trapping | Start Flowering | End Flowering | Last Collection | No. Sample Collections | Trap Days |
|---|---|---|---|---|---|---|---|
| 2016 | 22 April | 9 May | 1 June | 13 June | 24 June | 13 | 46 |
| 2017 | 4 April | 4 April | 7 April | 16 May | 15 May | 12 | 41 |
| 2018 | 23 March | 6 April | 21 April | 8 May | 7 May | 9 | 31 |
| 2019 1 | 5 April | 5 April | 20 April | 17 May | 22 May | 13 | 47 |
| Plot | 2016 | 2017 | 2018 | 2019 | All Years | |
|---|---|---|---|---|---|---|
| Species number | Outside | 27 | 30 | 24 | 27 | 46 |
| Inside | 8 | 16 | 15 | 22 | 30 | |
| Common species | 6 | 13 | 10 | 17 | 24 | |
| Sørensen index (SI) | 0.34 | 0.57 | 0.51 | 0.69 | 0.63 | |
| Shannon index (H’) | Outside | 2.27 | 2.67 | 1.43 | 2.13 | 2.44 |
| Inside | 1.65 | 2.22 | 1.61 | 1.64 | 1.94 | |
| Hmax | Outside | 3.30 | 3.40 | 3.18 | 3.30 | 3.83 |
| Inside | 2.08 | 2.77 | 2.71 | 3.09 | 3.40 | |
| Evenness (E) | Outside | 0.69 | 0.79 | 0.45 | 0.65 | 0.64 |
| Inside | 0.79 | 0.80 | 0.60 | 0.53 | 0.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meissle, M.; Waldburger, M.; Jeanneret, P.; Broggini, G.A.L.; Patocchi, A.; Romeis, J. Insect Pollinator Monitoring in and around a Netted Plot of Apple Trees—Biosafety Implications for Genetically Engineered Fruit Trees. Agronomy 2023, 13, 84. https://doi.org/10.3390/agronomy13010084
Meissle M, Waldburger M, Jeanneret P, Broggini GAL, Patocchi A, Romeis J. Insect Pollinator Monitoring in and around a Netted Plot of Apple Trees—Biosafety Implications for Genetically Engineered Fruit Trees. Agronomy. 2023; 13(1):84. https://doi.org/10.3390/agronomy13010084
Chicago/Turabian StyleMeissle, Michael, Mario Waldburger, Philippe Jeanneret, Giovanni A. L. Broggini, Andrea Patocchi, and Jörg Romeis. 2023. "Insect Pollinator Monitoring in and around a Netted Plot of Apple Trees—Biosafety Implications for Genetically Engineered Fruit Trees" Agronomy 13, no. 1: 84. https://doi.org/10.3390/agronomy13010084