



Salicylic Acid Improves Agro-Morphology, Yield and Ion Accumulation of Two Wheat (Triticum aestivum L.) Genotypes by Ameliorating the Impact of Salt Stress
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Planting Materials and Treatment
2.3. Treatment Imposition
2.4. Estimation of Relative Water Content
2.5. Gas Exchange Characteristics
2.6. Determination of Photosynthetic Pigment
2.7. Collection of Growth and Yield Data
2.8. Determination of Na and K Ion Concentration
2.9. Statistical Analysis
3. Results
3.1. Relative Water Content
3.2. Photosynthetic Rate
3.3. Stomatal Conductance
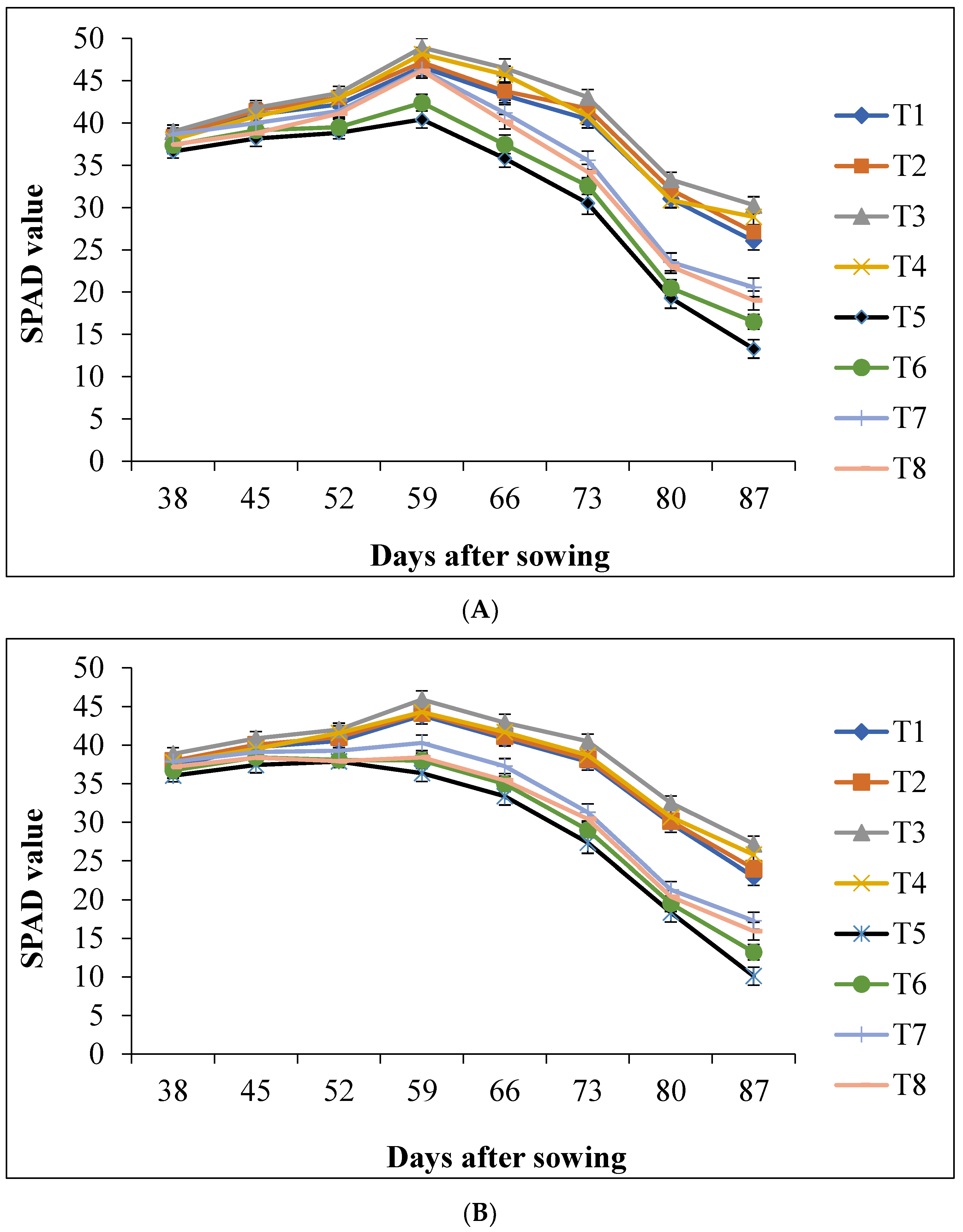
3.4. Chlorophyll Content (SPAD)
3.5. Growth and Biomass of Wheat
3.6. Days to Maturity
3.7. Yield-Contributing Parameters and Yield of Wheat
3.8. Ion Accumulation
4. Discussions
4.1. Relative Water Content and Gas Exchange Activities
4.2. Chlorophyll Content (SPAD)
4.3. Growth and Biomass of Wheat
4.4. Yield-Contributing Parameters and Yield of Wheat
4.5. Ion Accumulation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BSMRAU | Bangabandhu Sheikh Mujibur Rahman Agricultural University |
| SA | Salicylic acid |
| mM | Millimolar |
| dSm−1 | Decisiemens per meter |
| cm | Centimeter |
| G 200-4 | Genotype 200-4 |
| BARI | Bangladesh Agricultural Research Institute |
| Na+ | Sodium ion |
| K+ | Potassium ion |
| Cl− | Chloride ion |
| ROS | Reactive oxygen species |
| NaCl | Sodium chloride |
| cm | Centimeter |
| LSD | Least significant difference |
| SPAD | Soil plant analysis development |
| DAS | Days after sowing |
| Ca2+ | Calcium ion |
| Mg2+ | Magnesium ion |
| CRD | Completely randomized design |
| DS | Days of sowing |
| EC | Electrical conductivity |
| pH | Potential of Hydrogen |
| DW | Dry weight |
| HNO3 | Nitric acid |
| HCLO4 | Perchloric acid |
| v/v | Volume/volume |
| ANOVA | Analysis of variance |
| RWC | Relative water content |
| Pn | Photosynthetic rate |
| Gs | Stomatal conductance |
| AEZ | Agro-ecological zone |
References
- Tubiello, F.N.; Salvatore, M.; Rossi, S.; Ferrara, A.; Fitton, N.; Smith, P. The FAOSTAT database of greenhouse gas emissions from agriculture. Environ. Res. Lett. 2013, 8, 015009. [Google Scholar] [CrossRef]
- Dos Santos, T.B.; Ribas, A.F.; De Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological Responses to Drought, Salinity, and Heat Stress in Plants: A Review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Morad, M.; Sara, S.; Mohammad, D.; Javad, R.M.; Majid, R. Effect of salicylic acid on alleviation of salt stress on growth and some physiological traits of wheat. Int. J. Biosci. 2013, 3, 20–27. [Google Scholar] [CrossRef]
- EL Sabagh, A.; Islam, M.S.; Skalicky, M.; Ali Raza, M.; Singh, K.; Anwar Hossain, M.; Hossain, A.; Mahboob, W.; Iqbal, M.A.; Ratnasekera, D.; et al. Salinity stress in wheat (Triticum aestivum L.) in the changing climate: Adaptation and management strategies. Front. Agron. 2021, 3, 661932. [Google Scholar] [CrossRef]
- Alsahli, A.; Mohamed, A.K.; Alaraidh, I.; Al-Ghamdi, A.; Al-Watban, A.; El-Zaidy, M.; Alzahrani, S.M. Salicylic acid alleviates salinity stress through the modulation of biochemical attributes and some key antioxidants in wheat seedlings. Pak. J. Bot. 2019, 51, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Mguis, K. Morphological, eco-physiological and nutritional behavior of three accessions of Aegilops geniculata (Roth) and a durum wheat cultivar subjected to saline stress. Ph.D. Thesis, University Tunisia, Tunis, Tunisia, 2010. [Google Scholar]
- Abhinandan, K.; Skori, L.; Stanic, M.; Hickerson, N.; Jamshed, M.; Samuel, M.A. Abiotic stress signaling in wheat–an inclusive overview of hormonal interactions during abiotic stress responses in wheat. Front. Plant Sci. 2018, 9, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad, Z.; Hussain, F. Effect of NaCl salinity on the germination and seedling growth of seven wheat genotypes. Pak. J. Bot. 2012, 44, 1845–1850. [Google Scholar]
- Ashraf, M. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef]
- Rahnama, A.; James, R.A.; Poustini, K.; Munns, R. Stomatal conductance as a screen for osmotic stress tolerance in durum wheat growing in saline soil. Funct. Plant Biol. 2010, 37, 255–263. [Google Scholar] [CrossRef]
- Muchate, N.S.; Nikalje, G.C.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. Plant salt stress: Adaptive responses, tolerance mechanism and bioengineering for salt tolerance. Bot. Rev. 2016, 82, 371–406. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Ranjbar, G.; Hasanuzzaman, M.; Khanna, K.; Bhardwaj, R.; Ahmad, P. Salicylic acid-mediated regulation of morpho-physiological and yield attributes of wheat and barley plants in deferring salinity stress. J. Plant Growth Regul. 2022, 41, 1291–1303. [Google Scholar] [CrossRef]
- Abdi, N.; Van Biljon, A.; Steyn, C.; Labuschagne, M.T. Salicylic acid improves growth and physiological attributes and salt tolerance differentially in two bread wheat cultivars. Plants 2022, 11, 1853. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zimei, L.; Cui, J.; Jiangang, D.; Xia, X.; Liu, D.; Yu, J. Alleviation of chilling induced oxidative damage by salicylic acid pretreatment and related gene expression in eggplant seedlings. Plant Growth Regul. 2011, 65, 101–108. [Google Scholar] [CrossRef]
- Kang, G.; Li, G.; Zheng, B.; Han, Q.; Wang, C.; Zhu, Y.; Guo, T. Proteomic analysis on salicylic acid-induced salt tolerance in common wheat seedlings (Triticum aestivum L.). Biochim. Biophys. Acta—Proteins Proteom. 2012, 1824, 1324–1333. [Google Scholar] [CrossRef]
- Li, G.; Peng, X.; Wei, L.; Kang, G. Salicylic acid increases the contents of glutathione and ascorbate and temporally regulates the related gene expression in salt-stressed wheat seedlings. Gene 2013, 529, 321–325. [Google Scholar] [CrossRef]
- Farzana, S.; Rasel, M.; Tahjib-Ul-Arif, M.; Hossain, M.A.; Azam, M.G.; Al Galib, M.A.; Mahamud, A.G.M.S.U.; Hossain, M.A. Salicylic acid and thiourea ameliorate the negative impact of salt stress in wheat (Triticum aestivum L.) seedlings by up-regulating photosynthetic pigments, leaf water status, and antioxidant defense system. J. Phytol. 2021, 13, 130–145. [Google Scholar] [CrossRef]
- Abdel-Lattif, H.M.; Abbas, M.S.; Taha, M.H. Effect of salicylic acid on productivity and chemical constituents of some wheat (Triticum aestivum L.) varieties grown under saline conditions. J. Anim. Plant Sci. 2019, 29, 1054–1064. [Google Scholar]
- Morton, M.J.; Awlia, M.; Al-Tamimi, N.; Saade, S.; Pailles, Y.; Negrao, S.; Tester, M. Salt stress under the scalpel–dissecting the genetics of salt tolerance. Plant J. 2019, 97, 148–163. [Google Scholar] [CrossRef] [Green Version]
- Arora, N.K. Impact of climate change on agriculture production and its sustainable solutions. Environ. Sustain. 2019, 2, 95–96. [Google Scholar] [CrossRef] [Green Version]
- FAO. High Level Expert Forum-How to Feed the World in 2050. In Economic and Social Development; Food and Agricultural Organization of the United Nations: Rome, Italy, 2009. [Google Scholar]
- Ashik, T.; Islam, M.M.; Rana, M.S.; Jahan, K. Evaluation of salinity tolerant wheat (Triticum aestivum L.) Genotypes through Multivariate Analysis of Agronomic Traits. Agric. Sci. Digest. 2021, 1–7. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Schonfeld, M.A.; Johnson, R.C.; Carver, B.F.; Mornhinweg, D.W. Water relations in winter wheat as drought resistance indicator. Crop. Sci. 1988, 28, 526–531. [Google Scholar] [CrossRef]
- Rahman, A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium supplementation improves Na+/K+ ratio, antioxidant defense and glyoxalase systems in salt-stressed rice seedlings. Front. Plant Sci. 2016, 7, 609. [Google Scholar] [CrossRef] [PubMed]
- Fardaus, J.; Matin, M.A.; Hasanuzzaman, M.; Hossain, M.S.; Nath, S.D.; Hossain, M.A.; Rohman, M.M.; Hasanuzzaman, M. Exogenous salicylic acid-mediated physiological responses and improvement in yield by modulating antioxidant defense system of wheat under salinity. Not. Sci. Biol. 2017, 9, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Waqas, M.; Yaning, C.; Iqbal, H.; Shareef, M.; Rehman, H.; Yang, Y. Paclobutrazol improves salt tolerance in quinoa: Beyond the stomatal and biochemical interventions. J. Agron. Crop. Sci. 2017, 203, 315–322. [Google Scholar] [CrossRef]
- El-Bially, M.; Saudy, H.; El-Metwally, I.; Shahin, M. Efficacy of ascorbic acid as a cofactor for alleviating water deficit impacts and enhancing sunflower yield and irrigation water–use efficiency. Agric. Water Manag. 2018, 208, 132–139. [Google Scholar] [CrossRef]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Biswas, D.; Mannan, M.A.; Karim, M.A.; Miah, M.Y. Amelioration of salinity tolerance in foxtail millet by applying plant growth regulators. Bangladesh Agron. J. 2019, 22, 25–39. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Ashraf, M.H.P.J.C.; Harris, P.J. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Kordrostami, M.; Rabiei, B.; Kumleh, H.H. Biochemical, physiological and molecular evaluation of rice cultivars differing in salt tolerance at the seedling stage. Physiol. Mol. Biol. Plants 2017, 23, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, A.A.; Abd El-Kader, A.A.; Abouziena, H.F.; El-Awadi, M.; Gharib, F.A. Effects of benzoic acid and thiourea on growth and productivity of wheat (Triticum aestivum L.) plants. Int. Sci. Res. J. 2016, 72, 132–149. [Google Scholar]
- Suhaib, M.; Ahmad, I.; Munir, M.; Iqbal, M.B.; Abuzar, M.K.; Ali, S. Salicylic acid induced and ionic efficiency in wheat under salt stress. Pak. J. Agric. Sci. 2018, 31, 79–85. [Google Scholar] [CrossRef]
- Khan, M.I.; Shoukat, M.A.; Cheema, S.A.; Ali, S.; Azam, M.; Rizwan, M.; Al-Wabel, M.I. Foliar-and soil-applied salicylic acid and bagasse compost addition to soil reduced deleterious effects of salinity on wheat. Arab. J. Geosci. 2019, 12, 78. [Google Scholar] [CrossRef]
- Loutfy, N.; Sakuma, Y.; Gupta, D.K.; Inouhe, M. Modifications of water status, growth rate and antioxidant system in two wheat cultivars as affected by salinity stress and salicylic acid. J. Plant Res. 2020, 133, 549–570. [Google Scholar] [CrossRef]
- Mazher, A.A.; El-Quesni, E.F.; Farahat, M.M. Responses of ornamental and woody trees to salinity. World J. Agric. Sci. 2007, 3, 386–395. [Google Scholar]
- Ahmad, M.; Shahzad, A.; Iqbal, M.; Asif, M.; Hirani, A.H. Morphological and molecular genetic variation in wheat for salinity tolerance at germination and early seedling stage. Austral. J. Crop. Sci. 2013, 7, 66–74. [Google Scholar]
- Sanaullah, T.; Wahid, A.; Sadia, B.; Hanif, A.; Maqbool, N.; Arshad, T.; Kabir, Z. Exogenous application of thiourea ameliorates salt stress effects by alleviation of oxidative damage in hybrid maize. J. Agric. Sci. Technol. 2016, 6, 220–231. [Google Scholar] [CrossRef]
- Khanam, T.; Akhtar, N.; Halim, M.A.; Hossain, F. Effect of irrigation salinity on the growth and yield of two Aus rice cultivars of Bangladesh. Jahangirnagar Univ. J. Biol. Sci. 2018, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Enteshari, S.; Sharifian, S. Influence of salicylic acid on growth and some biochemical parameters in a C4 plant (Panicum miliaceum L.) under saline conditions. Afr. J. Biotechnol. 2012, 11, 621–627. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef] [PubMed]
- Khataar, M.; Mohammadi, M.H.; Shabani, F. Soil salinity and matric potential interaction on water use, water use efficiency and yield response factor of bean and wheat. Sci. Rep. 2018, 8, 2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharbatkhari, M.; Shobbar, Z.S.; Galeshi, S.; Nakhoda, B. Wheat stem reserves and salinity tolerance: Molecular dissection of fructan biosynthesis and remobilization to grains. Planta 2016, 244, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.; Gong, B.; Chen, Y.; Wang, J.; Wei, M.; Shi, Q. Photosynthetic capacity, ion homeostasis and reactive oxygen metabolism were involved in exogenous salicylic acid increasing cucumber seedlings tolerance to alkaline stress. Sci. Hortic. 2018, 235, 413–423. [Google Scholar] [CrossRef]
- Silva, A.A.R.D.; Lima, G.S.D.; Azevedo, C.A.V.D.; Veloso, L.L.D.S.A.; Gheyi, H.R. Salicylic acid as an attenuator of salt stress in soursop. Rev. Caatinga. 2020, 33, 1092–1101. [Google Scholar] [CrossRef]
- Desoky, E.S.M.; Merwad, A.R.M. Improving the salinity tolerance in wheat plants using salicylic and ascorbic acids. J. Agric. Sci. 2015, 7, 203. [Google Scholar] [CrossRef] [Green Version]
- Jini, D.; Joseph, B. Physiological mechanism of salicylic acid for alleviation of salt stress in rice. Rice Sci. 2017, 24, 97–108. [Google Scholar] [CrossRef]
- Fardus, J.; Matin, M.A.; Hasanuzzaman, M.; Hossain, M.A.; Hasanuzzaman, M. Salicylic acid-induced improvement in germination and growth parameters of wheat under salinity stress. J. Anim. Plant Sci. 2018, 28, 197–207. [Google Scholar]
- Iqbal, M.; Ashraf, M. Wheat seed priming in relation to salt tolerance: Growth, yield and levels of free salicylic acid and polyamines. Ann. Bot. Fenn 2006, 43, 250–259. [Google Scholar]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.; Meng, L.; Mao, P.C.; Jia, Y.Q.; Shi, Y.J. Role of exogenous salicylic acid in alleviating cadmium-induced toxicity in Kentucky bluegrass. Biochem. Syst. Ecol. 2013, 50, 269–276. [Google Scholar] [CrossRef]
- Koc, E.; Ustun, A.S.; Celik, N. Effect of exogenously applied salicylic acid on cadmium chloride-induced oxidative stress and nitrogen metabolism in tomato (Lycopersicon esculentum L.). Turk. J. Biol. 2013, 37, 361–369. [Google Scholar]
- Yan, S.; Dong, X. Perception of the plant immune signal salicylic acid. Curr. Opin. Plant Biol. 2014, 20, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Assaha, D.V.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef] [Green Version]
- Sa, G.; Yao, J.; Deng, C.; Liu, J.; Zhang, Y.; Zhu, Z.; Zhang, Y.; Ma, X.; Zhao, R.; Lin, S.; et al. Amelioration of nitrate uptake under salt stress by ectomycorrhiza with and without a Hartig net. New Phytol. 2019, 222, 1951–1964. [Google Scholar] [CrossRef]
- Hafez, E.M. Influence of salicylic acid on ion distribution, enzymatic activity and some agromorphological characteristics of wheat under salt-affected soil. Egypt. J. Agron. 2016, 38, 455–469. [Google Scholar] [CrossRef] [Green Version]
- Abbas, M.; Nawaz, S.; Ahmed, N.; Abbas, G.; Usman, M.; Bashir, H.M.N.; Ali, A. Salicylic acid in amelioration of salt tolerance in wheat (Triticum aestivum L.) under in vitro conditions. Int. J. Earth Sci. 2016, 3, 44–49. [Google Scholar]
- Noreen, S.; Fatima, K.; Athar, H.U.R.; Ahmad, S.; Hussain, K. Enhancement of physio-biochemical parameters of wheat through exogenous application of salicylic acid under drought stress. J. Anim. Plant Sci. 2017, 27, 153–163. [Google Scholar]
- Liu, S.; Dong, Y.; Xu, L.; Kong, J. Effects of foliar applications of nitric oxide and salicylic acid on salt-induced changes in photosynthesis and antioxidative metabolism of cotton seedlings. Plant Growth Regul. 2014, 73, 67–78. [Google Scholar] [CrossRef]
- Nazar, R.; Iqbal, N.; Syeed, S.; Khan, N.A. Salicylic acid alleviates decreases in photosynthesis under salt stress by enhancing nitrogen and sulfur assimilation and antioxidant metabolism differentially in two mungbean cultivars. J. Plant Physiol. 2011, 168, 807–815. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Sources of Variation | DF | Mean Square Values | ||
|---|---|---|---|---|
| Relative Water Content (%) | Photosynthetic Rate (µmol m−2s−1) | Stomatal Conductance (mmol m−2s−1) | ||
| Variety (V) | 1 | 586.094 * | 3.20192 * | 0.0096 * |
| Salinity (S) | 1 | 2738.63 * | 166.201 * | 0.093 * |
| Variety × salinity | 1 | 322.988 * | 0.389 * | 0.0008 * |
| Salicylic acid (SA) | 3 | 60.2644 * | 1.677 * | 0.0005 * |
| Variety × SA | 3 | 0.420683 * | 0.051 * | 0.000008 * |
| Salinity × SA | 3 | 17.1832 * | 0.025 * | 0.00009 * |
| V × S × SA | 3 | 0.0311 ** | 0.0018 * | 0.00004 ** |
| Error | 48 | 1.64564 | 0.576 | 0.00005 |
| Variety | Treatment | Relative Water Content (%) | Photosynthetic Rate (µmol m−2s−1) | Stomatal Conductance (mmol m−2s−1) | |
|---|---|---|---|---|---|
| BU 200-4 | Control | T1 | 84.025 cd | 15.654 ab | 0.258 c |
| 0.5 mM SA | T2 | 85.368 ab | 16.005 ab | 0.261 b | |
| 1 mM SA | T3 | 86.573 a | 16.440 a | 0.267a | |
| 1.5 mM SA | T4 | 84.853 bc | 15.740 ab | 0.263 b | |
| Salinity (12 dS m−1) | T5 | 73.640 h | 12.268 cd | 0.185 j | |
| Salinity + 0.5 mM SA | T6 | 77.883 g | 12.520 cd | 0.192 h | |
| Salinity + 1 mM SA | T7 | 80.450 f | 13.175 c | 0.205 g | |
| Salinity + 1.5 mM SA | T8 | 74.485 h | 12.360 cd | 0.189 i | |
| BARI gom-25 | Control | T1 | 82.753 e | 15.110 b | 0.241 f |
| 0.5 mM SA | T2 | 83.798 cde | 15.380 b | 0.244 e | |
| 1 mM SA | T3 | 84.698 bc | 15.698 ab | 0.248 d | |
| 1.5 mM SA | T4 | 83.333 de | 15.238 b | 0.245 e | |
| Salinity (12 dS m−1) | T5 | 63.458 k | 12.085 d | 0.155 m | |
| Salinity + 0.5 mM SA | T6 | 67.433 j | 12.230 cd | 0.160 l | |
| Salinity + 1 mM SA | T7 | 69.323 i | 12.710 cd | 0.171 k | |
| Salinity + 1.5 mM SA | T8 | 64.064 k | 12.133 d | 0.159 l | |
| Sources of Variation | DF | Mean Square Values | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 38 DAS | 45 DAS | 52 DAS | 59 DAS | 66 DAS | 73 DAS | 80 DAS | 87 DAS | ||
| Variety (V) | 1 | 4.463 | 14.784 | 50.8690 * | 305.988 * | 173.712 * | 165.573 * | 30.581 * | 160.643 |
| Salinity (S) | 1 | 14.307 | 61.898 | 129.795 * | 414.631 * | 630.010 * | 1236.40 * | 1789.29 * | 1866.28 * |
| Variety × salinity | 1 | 0.054 * | 1.102 * | 0.4196 * | 22.920 ** | 0.203 * | 3.950 * | 1.82 ** | 0.0627 * |
| Salicylic acid (SA) | 3 | 6.625 * | 5.478 * | 8.312 * | 38.171 * | 40.645 * | 35.274 * | 26.103 * | 101.445 * |
| Variety × SA | 3 | 0.143 ** | 0.041 ** | 0.650 * | 4.568 * | 3.531 * | 0.448 * | 0.1704 * | 0.0092 * |
| Salinity × SA | 3 | 0.560 * | 0.735 * | 1.431 * | 7.321 * | 2.965 * | 5.576 * | 4.870 * | 7.755 * |
| V × S × SA | 3 | 0.201 ** | 0.121 * | 1.392 * | 1.625 ** | 0.1363 ** | 0.739 ** | 2.373 * | 0.0092 ** |
| Error | 48 | 2.731 | 1.518 | 1.633 | 2.111 | 1.720 | 1.630 | 1.126 | 0.514 |
| Sources of Variation | DF | Mean Square Values | |||||
|---|---|---|---|---|---|---|---|
| Plant Height (cm) | No of Tillers/Plant | Leaf Number/Plant | Leaf Areas (cm2) | Dry Weight (g)/Plant | Days to Maturity | ||
| Variety (V) | 1 | 151.936 * | 7.459 * | 22.314 * | 3139.64 * | 13.829 * | 0.0156 * |
| Salinity (S) | 1 | 2464.75 * | 36.466 * | 777.503 * | 20908.4 * | 71.170 * | 141.016 * |
| Variety × salinity | 1 | 193.106 ** | 1.880 * | 58.122 * | 261.711 ** | 1.425 * | 19.141 * |
| Salicylic acid (SA) | 3 | 116.341 * | 3.307 * | 69.951 * | 2307.60 * | 3.228 * | 12.516 * |
| Variety × SA | 3 | 0.296 * | 0.027 * | 0.596 * | 18.822 * | 0.173 * | 0.057 * |
| Salinity × SA | 3 | 3.554 * | 0.174 * | 13.708 * | 512.809 * | 0.152 * | 16.474 * |
| V × S × SA | 3 | 0.334 ** | 0.014 ** | 0.758 ** | 0.659 * | 0.046 * | 0.599 * |
| Error | 48 | 1.535 | 0.037 | 0.377 | 2.7406 | 0.051 | 1.036 |
| Variety | Treatment | Plant Height (cm) | No of Tillers/Plant | Leaf Number/Plant | Leaf Area (cm2) | Dry Weight (g)/Plant | Days to Maturity | |
|---|---|---|---|---|---|---|---|---|
| BU 200-4 | Control | T1 | 65.90 d | 6.00 de | 33.00 cd | 106.26 g | 6.13 c | 103.50 b |
| 0.5 mM SA | T2 | 67.95 bc | 6.50 c | 33.25 c | 139.00 c | 6.74 b | 102.50 c | |
| 1 mM SA | T3 | 71.80 a | 7.25 a | 35.50 a | 146.05 a | 7.64 a | 102.00 c | |
| 1.5 mM SA | T4 | 68.93 b | 7.25 a | 34.50 b | 141.66 b | 7.36 a | 104.00 a | |
| Salinity (12 dS m−1) | T5 | 55.60 gh | 5.15 fg | 25.50 i | 90.42 i | 4.68 g | 99.50 e | |
| Salinity + 0.5 mM SA | T6 | 59.85 f | 5.40 f | 26.75 h | 98.65 h | 5.00 f | 100.50 d | |
| Salinity + 1 mM SA | T7 | 63.33 e | 6.01 de | 32.50 d | 108.90 f | 5.67 e | 102.00 c | |
| Salinity + 1.5 mM SA | T8 | 60.05 f | 5.78 e | 31.25 e | 106.57 g | 5.26 f | 102.50 c | |
| BARI gom-25 | Control | T1 | 66.58 cd | 5.81 e | 33.42 c | 98.69 h | 5.91 de | 105.00 a |
| 0.5 mM SA | T2 | 68.12 bc | 6.22 d | 34.16 b | 129.31 e | 6.12 c | 103.50 b | |
| 1 mM SA | T3 | 72.33 a | 6.81 b | 36.02 a | 134.52 d | 6.89 b | 102.75 c | |
| 1.5 mM SA | T4 | 69.12 b | 6.80 b | 35.55 a | 130.59 e | 6.42 c | 105.00 a | |
| Salinity (12 dS m−1) | T5 | 49.36 j | 4.18 i | 23.16 k | 75.66 l | 3.61 j | 97.75 f | |
| Salinity + 0.5 mM SA | T6 | 53.02 i | 4.35 i | 24.22 j | 81.25 k | 3.81 ij | 99.25 e | |
| Salinity + 1 mM SA | T7 | 56.22 g | 4.89 gh | 28.61 f | 88.32 j | 4.31 h | 101.25 c | |
| Salinity + 1.5 mM SA | T8 | 54.01 hi | 4.81 h | 27.66 g | 87.10 j | 4.01 i | 101.75 cd | |
| Sources of Variation | DF | Mean Square Values | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant Height (cm) | Total Tillers Per Plant | Effective Tillers Per Plant | Spike Length (cm) | No. of Spikelets/Spike | Filled Grains/Spike | Unfilled Grains/Spike | Thousand-Grain Weight (g) | Grain Yield (g)/Plant | Straw Yield (g)/Plant | ||
| Variety (V) | 1 | 30.802 * | 6.663 * | 3.432 * | 6.3397 * | 41.433 * | 222.700 * | 0.167 | 500.948 * | 16.221 * | 5.475 * |
| Salinity (S) | 1 | 5322.80 * | 41.845 * | 46.581 * | 22.369 * | 70.241 * | 434.592 * | 0.880 * | 1655.63 * | 242.888 * | 543.123 * |
| Variety × salinity | 1 | 257.282 ** | 1.095 * | 0.084 * | 1.804 * | 0.925 * | 0.000004 ** | 0.042 * | 0.323 * | 0.570 * | 12.531 * |
| Salicylic acid (SA) | 3 | 103.695 * | 0.893 * | 1.215 * | 3.229 * | 4.226 * | 121.780 * | 0.110 * | 35.379 * | 5.759 * | 4.051 * |
| Variety × SA | 3 | 7.881 * | 0.066 * | 0.004 * | 0.086 * | 0.020 * | 0.00005 * | 0.002 * | 1.086 * | 0.069 * | 0.025 * |
| Salinity × SA | 3 | 32.565 * | 0.024 * | 0.303 * | 0.686 * | 1.462 * | 15.506 * | 0.032 * | 4.729 * | 0.146 * | 0.028 * |
| V × S × SA | 3 | 4.156 ** | 0.004 * | 0.023 * | 0.162 * | 0.003 ** | 0.00002 ** | 0.002 * | 0.064 * | 0.0101 ** | 0.063 ** |
| Error | 48 | 1.756 | 0.0541 | 0.022 | 0.0511 | 0.341 | 1.016 | 0.003 | 1.167 | 0.176 | 0.135 |
| Variety | Treatment | Plant Height (cm) | Total Tillers/Plant | Effective Tillers/Plant | Spike Length (cm) | No. of Spikelets/Spike | Filled Grains/Spike | Unfilled Grains/Spike | Thousand-Grain Weight (g) | Grain Yield (g)/Plant | Straw Yield (g)/Plant | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BU 200-4 | Control | T1 | 79.21 d | 7.14 bc | 6.29 c | 10.64 cd | 19.02 bc | 41.99 c | 0.27 ef | 50.37 c | 12.01 cd | 12.08 f |
| 0.5 mM SA | T2 | 79.59 d | 7.33 b | 6.88 a | 10.92 bc | 19.56 ab | 43.11 b | 0.19 gh | 52.29 b | 12.67 b | 12.33 ef | |
| 1 mM SA | T3 | 82.93 b | 7.63 a | 6.93 a | 11.23 a | 19.87 a | 46.73 a | 0.12 h | 54.30 a | 13.41 a | 13.16 cd | |
| 1.5 mM SA | T4 | 80.49 cd | 7.88 a | 6.28 c | 10.74 bc | 19.06 bc | 42.93 bc | 0.25 efg | 50.91 c | 12.96 ab | 12.79 de | |
| Salinity (12 dS m−1) | T5 | 60.83 g | 5.82 e | 4.55 g | 8.88 h | 16.49 f | 34.55 g | 0.53 bc | 39.61 h | 8.24 hi | 7.09 jk | |
| Salinity + 0.5 mM SA | T6 | 67.81 e | 5.93 e | 4.98 f | 10.28 e | 17.48 e | 36.85 f | 0.46 cd | 41.81 g | 8.70 h | 7.29 ijk | |
| Salinity + 1 mM SA | T7 | 68.96 e | 6.38 d | 5.22 e | 10.67 cd | 18.11 de | 42.82 bc | 0.25 efg | 43.79 f | 9.94 f | 8.44 g | |
| Salinity + 1.5 mM SA | T8 | 67.69 e | 6.38 d | 5.08 ef | 10.31 e | 18.01 de | 39.69 d | 0.32 e | 42.53 g | 9.34 g | 7.77 hi | |
| BARI gom-25 | Control | T1 | 81.85 bc | 6.88 c | 6.00 d | 10.24 e | 17.60 e | 38.26 e | 0.31 e | 45.20 e | 11.42 e | 13.55 bc |
| 0.5 mM SA | T2 | 84.95 a | 6.97 c | 6.35 bc | 10.72 bc | 18.22 de | 39.38 d | 0.25 efg | 46.30 e | 11.80 de | 13.91 b | |
| 1 mM SA | T3 | 85.90 a | 7.26 b | 6.55 b | 10.98 ab | 18.40 cd | 43.00 b | 0.21 fg | 48.50 d | 12.50 bc | 14.72 a | |
| 1.5 mM SA | T4 | 80.00 cd | 7.29 b | 5.90 d | 10.41 de | 17.81 de | 39.20 de | 0.25 efg | 46.05 e | 12.06 cd | 14.06 b | |
| Salinity (12 dS m−1) | T5 | 55.70 h | 5.06 g | 4.00 h | 8.41 i | 14.60 h | 30.82 i | 0.67 a | 34.25 k | 7.18 k | 6.88 k | |
| Salinity + 0.5 mM SA | T6 | 62.55 fg | 5.11 g | 4.53 g | 9.22 g | 15.68 g | 33.12 h | 0.60 ab | 35.83 j | 7.59 jk | 7.06 jk | |
| Salinity + 1 mM SA | T7 | 63.90 f | 5.41 f | 4.66 g | 9.59 f | 16.21 fg | 39.09 de | 0.42 d | 37.40 i | 8.66 h | 7.91 h | |
| Salinity + 1.5 mM SA | T8 | 61.55 g | 5.29 fg | 4.50 g | 9.06 gh | 16.20 fg | 35.96 f | 0.48 cd | 37.30 i | 8.01 ij | 7.55 hij | |
| Sources of Variation | DF | Mean Square Values | |||||
|---|---|---|---|---|---|---|---|
| %Na+ | %K+ | Na+/K+ | |||||
| Leaves | Roots | Leaves | Roots | Leaves | Roots | ||
| Variety (V) | 1 | 280.206 * | 2421.61 * | 718.935 * | 2843.74 * | 0.00395 * | 0.413 * |
| Salinity (S) | 1 | 75,872.3 * | 35,653.5 * | 336,840.0 * | 529,799.0 * | 1.209 * | 9.351 * |
| Variety × salinity | 1 | 346.322 * | 387.114 * | 7703.73 * | 6158.93 * | 0.0122 * | 0.339 * |
| Salicylic acid (SA) | 3 | 7779.93 * | 16,360.4 * | 4174.71 * | 8035.60 * | 0.0978 * | 0.692 * |
| Variety × SA | 3 | 23.4053 * | 371.237 * | 61.1594 * | 74.001 * | 0.0001 * | 0.0168 * |
| Salinity × SA | 3 | 2686.11 * | 955.824 * | 2345.17 * | 2379.39 * | 0.0541 * | 0.129 * |
| V × S × SA | 3 | 76.9926 * | 204.659 * | 66.5672 ** | 187.345 * | 0.00032 ** | 0.0091 * |
| Error | 48 | 60.4481 | 144.617 | 387.276 | 25.313 | 0.00087 | 0.0040 |
| Variety | Treatments | mM (Na+) | mM (K+) | Na+/K+ | ||||
|---|---|---|---|---|---|---|---|---|
| Leaves | Roots | Leaves | Roots | Leaves | Roots | |||
| BU 200-4 | Control | T1 | 106.7 hi | 214.7 cde | 483.4 c | 320.6 e | 0.221 h | 0.670 g |
| 0.5 mM SA | T2 | 90.9 jk | 174.4 ijk | 493.5 bc | 359.7 d | 0.184 hij | 0.485 h | |
| 1 mM SA | T3 | 89.3 jk | 159.3 k | 494.4 bc | 388.0 b | 0.181 ij | 0.411 h | |
| 1.5 mM SA | T4 | 92.2 j | 160.0 k | 491.8 bc | 377.5 c | 0.188 hij | 0.424 h | |
| Salinity (12 dS m−1) | T5 | 204.9 b | 259.5 b | 330.7 gh | 183.7 i | 0.619 b | 1.413 c | |
| Salinity + 0.5 mM SA | T6 | 160.1 cd | 228.9 c | 355.4 fg | 193.8 h | 0.450 cd | 1.181 d | |
| Salinity + 1 mM SA | T7 | 123.2 g | 180.5 hij | 403.2 d | 219.2 g | 0.306 g | 0.823 f | |
| Salinity + 1.5 mM SA | T8 | 148.0 e | 208.8 def | 381.2 de | 199.8 h | 0.388 ef | 1.045 e | |
| BARI gom-25 | Control | T1 | 109.2 h | 222.7 cd | 512.3 ab | 312.0 f | 0.213 hi | 0.714 g |
| 0.5 mM SA | T2 | 90.3 jk | 186.7 ghi | 521.7 a | 348.9 c | 0.173 j | 0.535 h | |
| 1 mM SA | T3 | 81.4 k | 161.8 k | 522.5 a | 370.9 a | 0.156 j | 0.436 h | |
| 1.5 mM SA | T4 | 96.4 ij | 165.3 jk | 521.2 a | 362.3 b | 0.185 hij | 0.456 h | |
| Salinity (12 dS m−1) | T5 | 217.9 a | 286.2 a | 323.0 h | 156.4 k | 0.675 a | 1.829 a | |
| Salinity + 0.5 mM SA | T6 | 167.0 c | 259.8 b | 346.1 fgh | 166.3 j | 0.483 c | 1.562 b | |
| Salinity + 1 mM SA | T7 | 135.4 f | 196.2 fgh | 383.3 d | 183.7 i | 0.353 f | 1.068 e | |
| Salinity + 1.5 mM SA | T8 | 151.1 de | 202.8 efg | 357.1 ef | 165.8 j | 0.423 de | 1.223 d | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fairoj, S.A.; Islam, M.M.; Islam, M.A.; Zaman, E.; Momtaz, M.B.; Hossain, M.S.; Jahan, N.A.; Shams, S.-N.-U.; Urmi, T.A.; Rasel, M.A.; et al. Salicylic Acid Improves Agro-Morphology, Yield and Ion Accumulation of Two Wheat (Triticum aestivum L.) Genotypes by Ameliorating the Impact of Salt Stress. Agronomy 2023, 13, 25. https://doi.org/10.3390/agronomy13010025
Fairoj SA, Islam MM, Islam MA, Zaman E, Momtaz MB, Hossain MS, Jahan NA, Shams S-N-U, Urmi TA, Rasel MA, et al. Salicylic Acid Improves Agro-Morphology, Yield and Ion Accumulation of Two Wheat (Triticum aestivum L.) Genotypes by Ameliorating the Impact of Salt Stress. Agronomy. 2023; 13(1):25. https://doi.org/10.3390/agronomy13010025
Chicago/Turabian StyleFairoj, Syeda Afia, Md. Moshiul Islam, Md. Ariful Islam, Erin Zaman, Milia Bente Momtaz, Md. Saddam Hossain, Nilufar Akhtar Jahan, Shahjadi-Nur-Us Shams, Tahmina Akter Urmi, Md Asadujjaman Rasel, and et al. 2023. "Salicylic Acid Improves Agro-Morphology, Yield and Ion Accumulation of Two Wheat (Triticum aestivum L.) Genotypes by Ameliorating the Impact of Salt Stress" Agronomy 13, no. 1: 25. https://doi.org/10.3390/agronomy13010025