
Light Intensity and Growth Media Influence Growth, Nutrition, and Phytochemical Content in Trachyandra divaricata Kunth

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Location
2.2. Collection of Plant Materials and Experimental Design
2.3. Treatments
2.4. Determination of Plant Growth
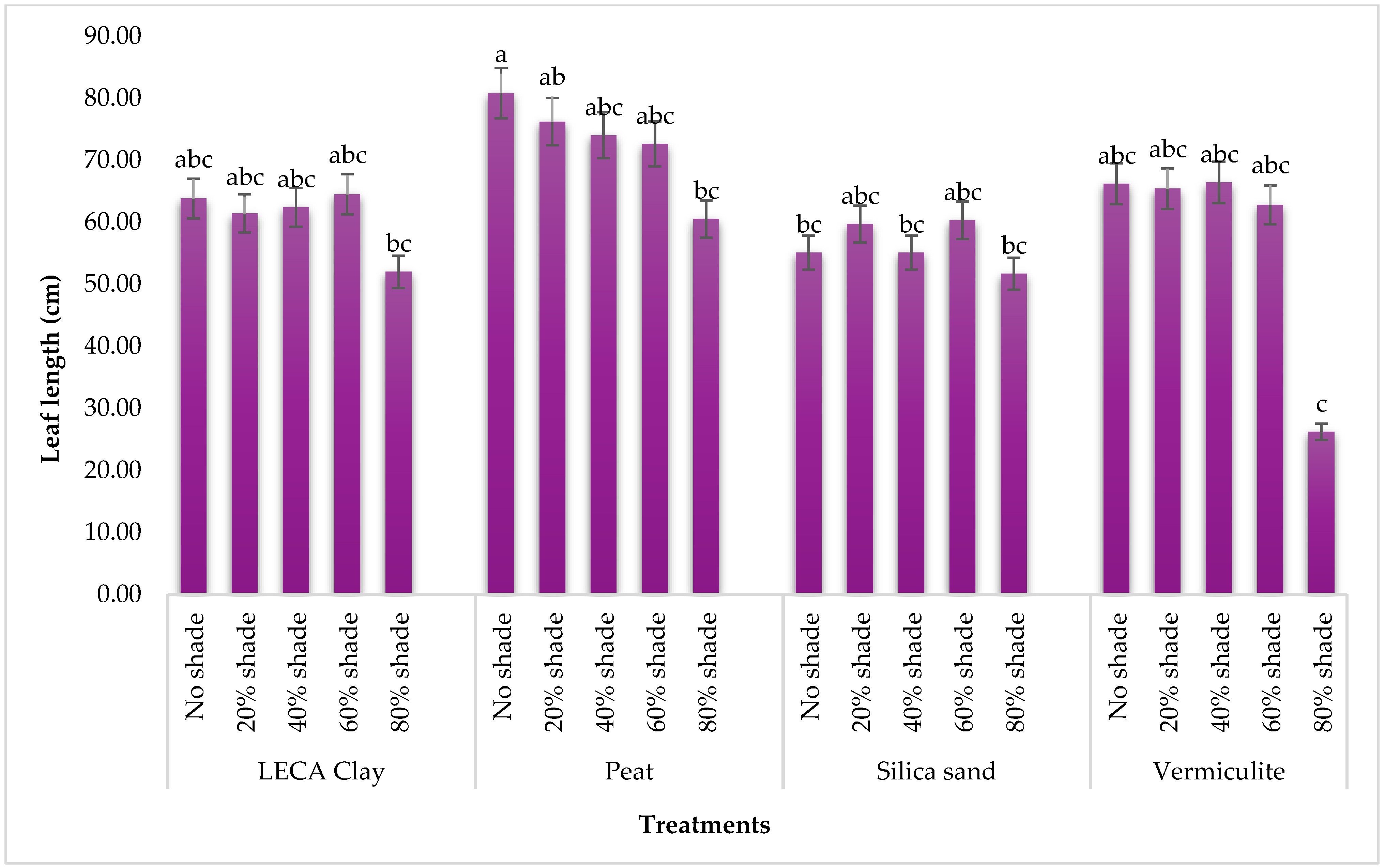
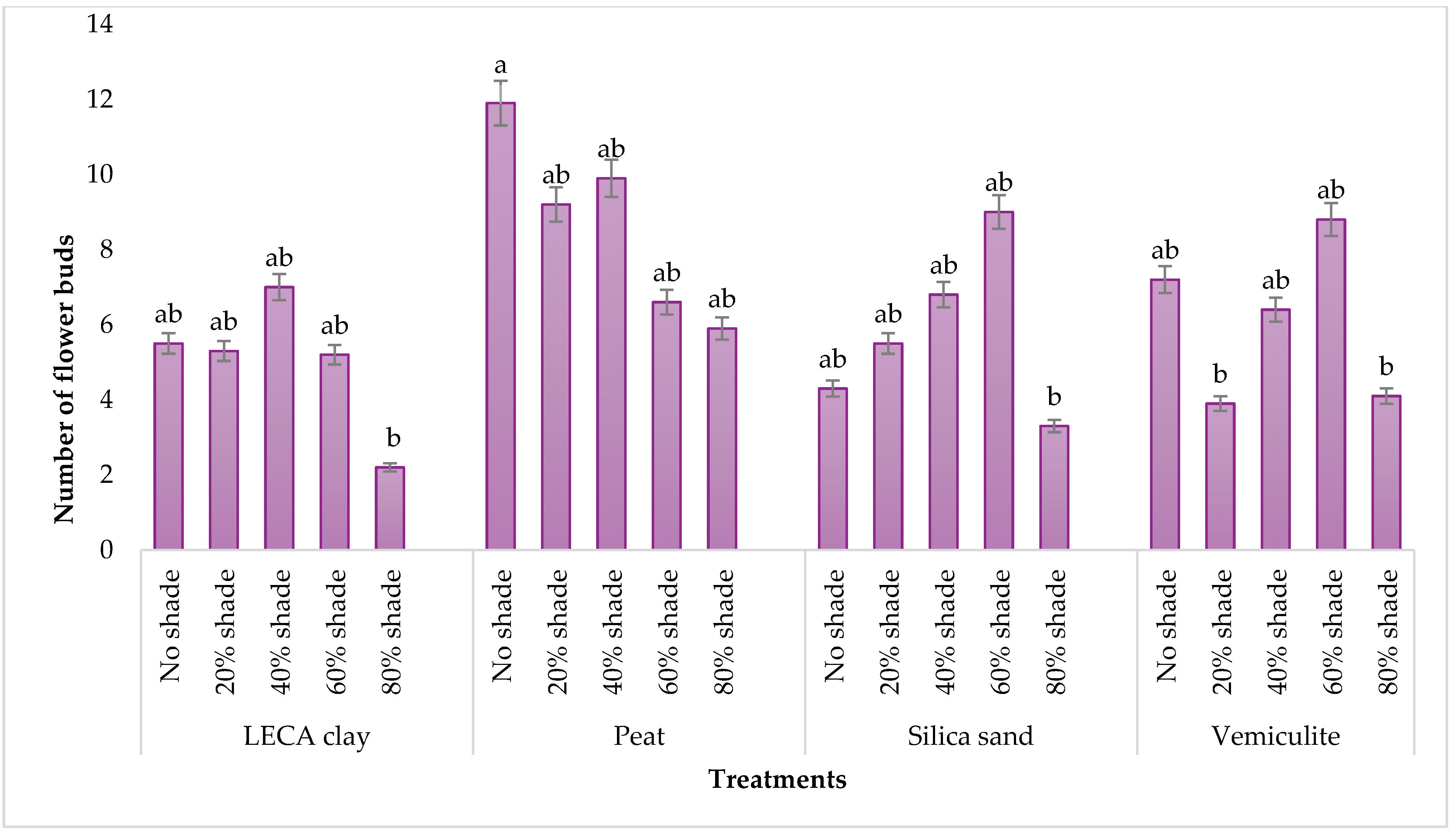
2.4.1. Leaf Length and the Number of Flower Buds
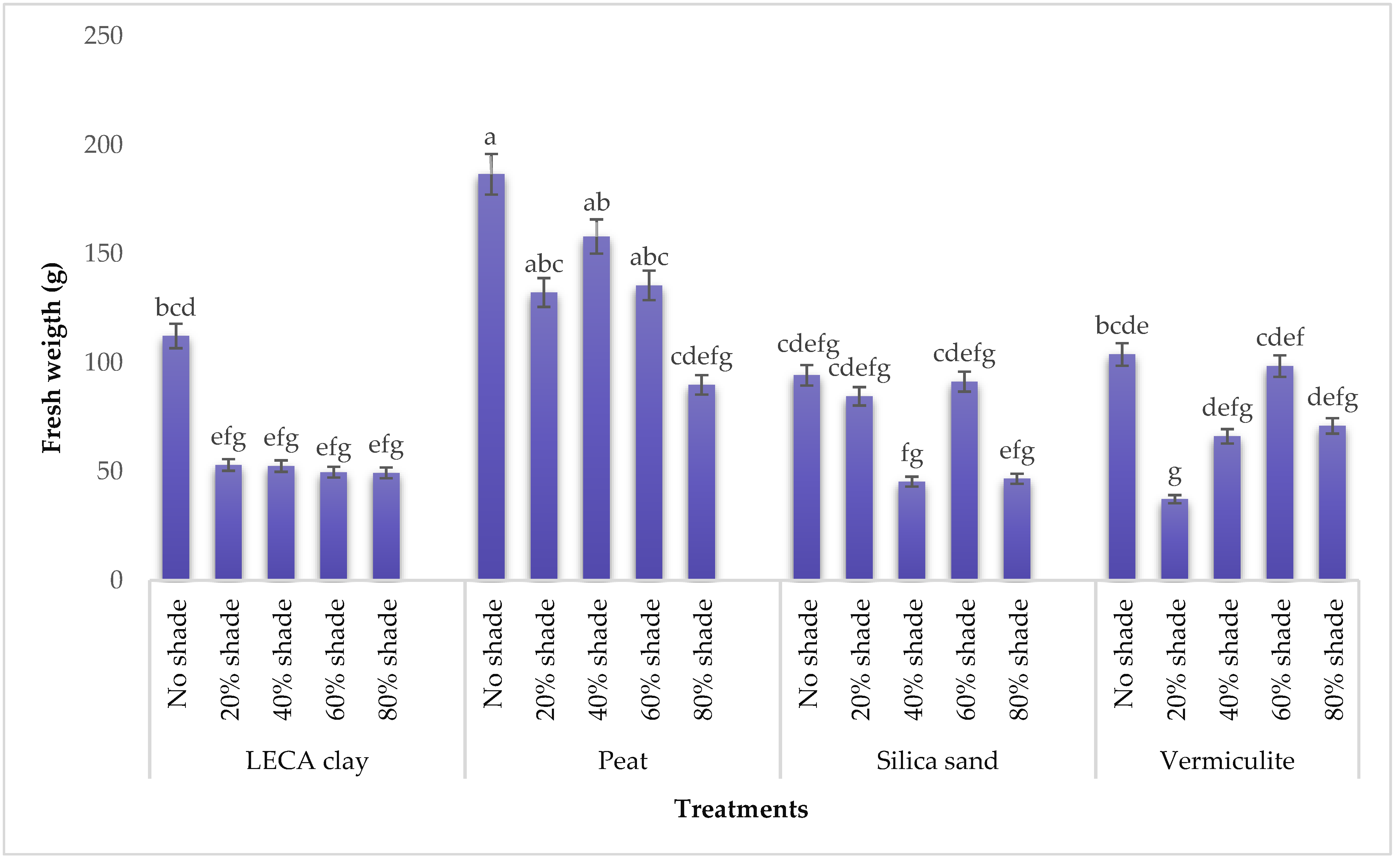
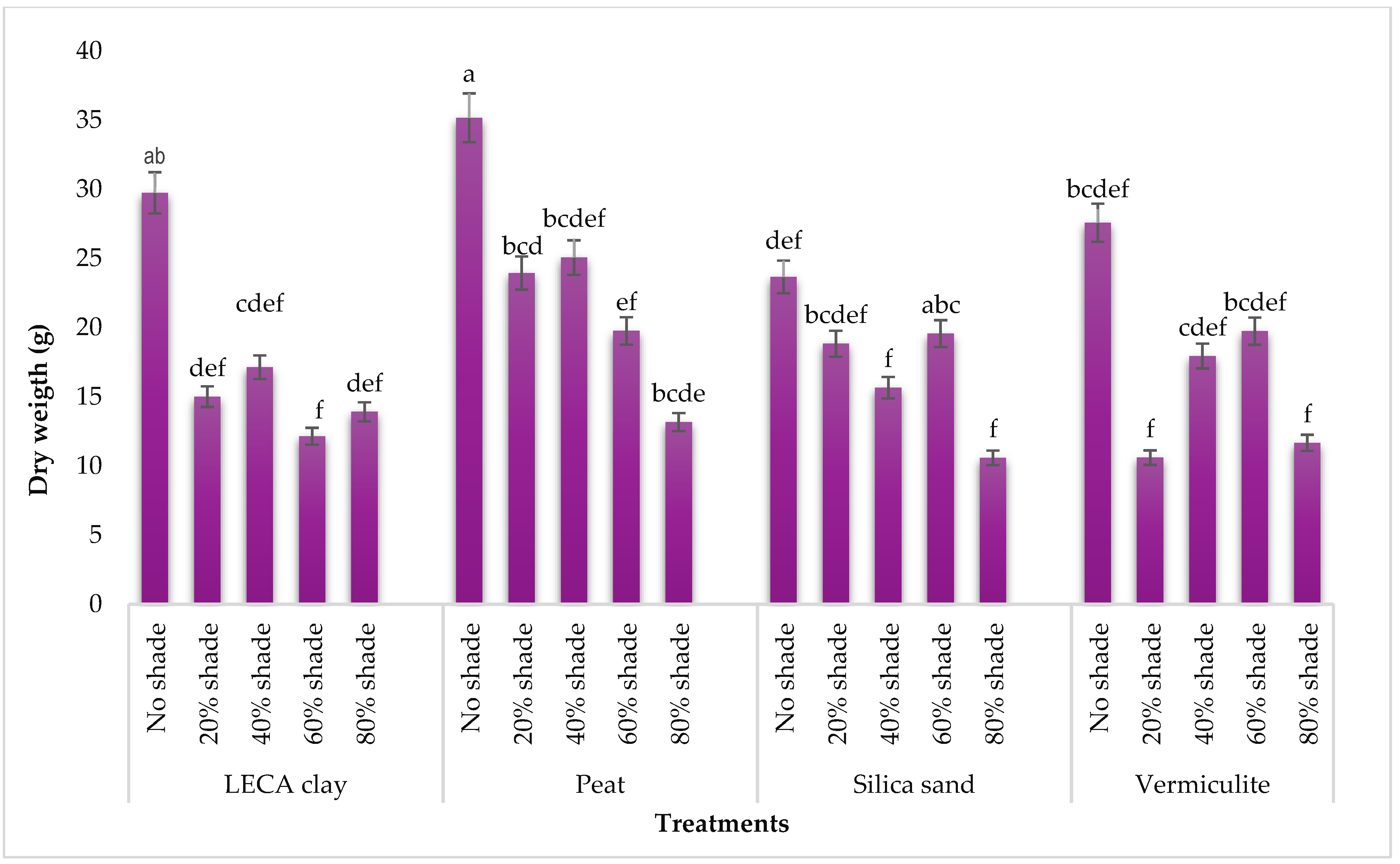
2.4.2. Plant Weight
2.5. Chlorophyll Concentration
2.6. Mineral Analysis of Flower Buds
2.7. Proximate Analysis of Flower Buds
2.7.1. Moisture Content
2.7.2. Crude Fat Content
2.7.3. Ash Content
2.7.4. Crude Protein
2.7.5. Neutral Detergent Fibre (NDF)
2.8. Phytochemicals and Antioxidant Assays
2.8.1. Preparation of Plant Samples
2.8.2. Polyphenol Assay
2.8.3. Determination of Flavonols
2.8.4. DPPH Free Radical Antioxidant Content
2.8.5. Ferric Reducing Antioxidant Power (FRAP) Assay
2.8.6. TEAC Free Radical Scavenging Activity
2.9. Statistical Analysis
3. Results
3.1. Effect of Growing Media and Shade Levels on Plant Growth
3.1.1. Leaf Length and Flower Bud Number
3.1.2. Total Fresh and Dry Weight of T. divaricata
3.1.3. Effect of Growing Media and Shade Levels on Chlorophyll Content
3.2. Effect of Growing Media and Shade Levels on Mineral Composition of Flower Buds
3.2.1. Macronutrients
3.2.2. Micronutrients
3.3. Effect of Growing Media and Shade Levels on Proximate Composition of Flower Buds
3.4. Effect of Growing Media and Shade on Phytochemicals and the Antioxidant Content of Flower Buds
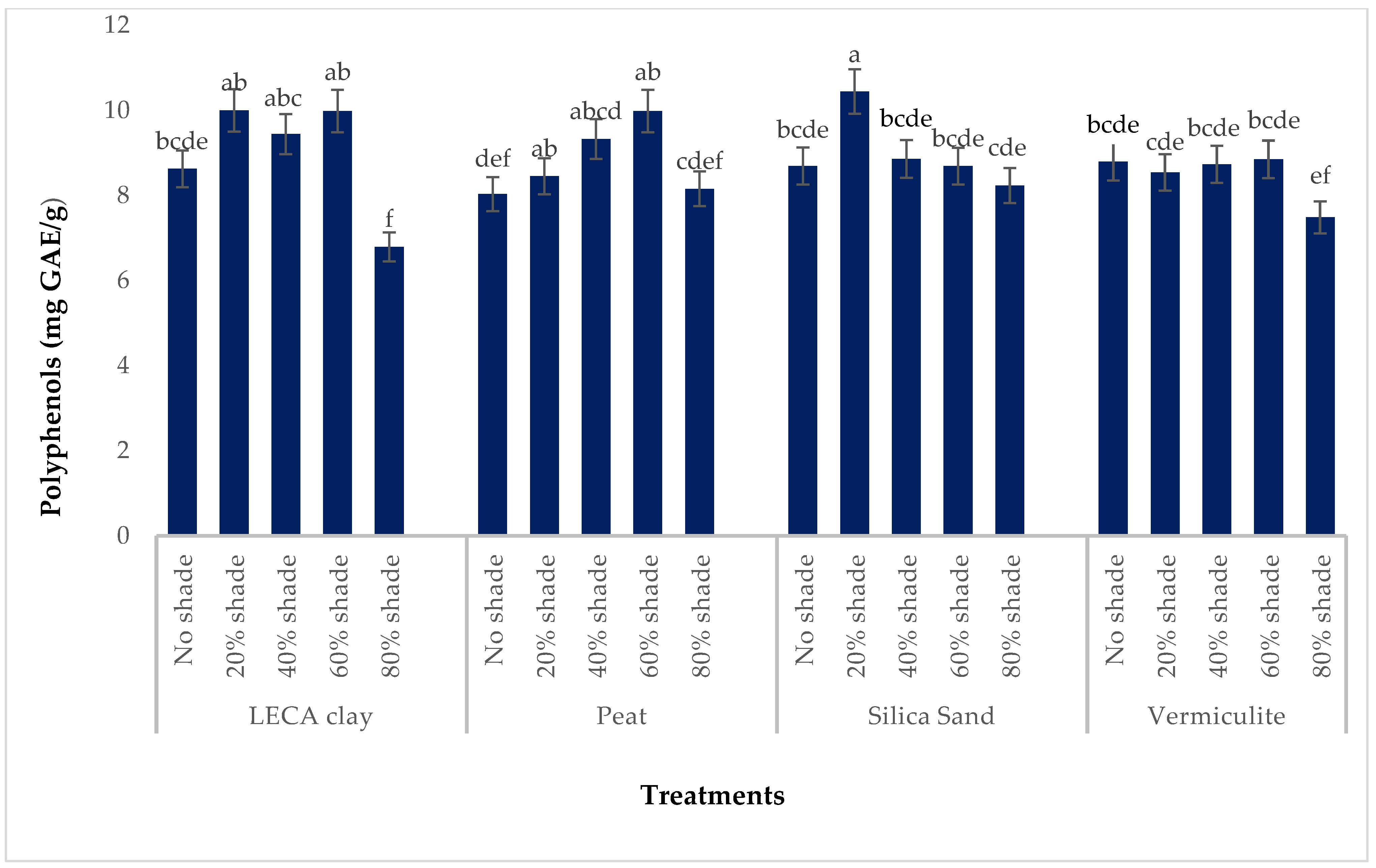
3.4.1. Total Polyphenols
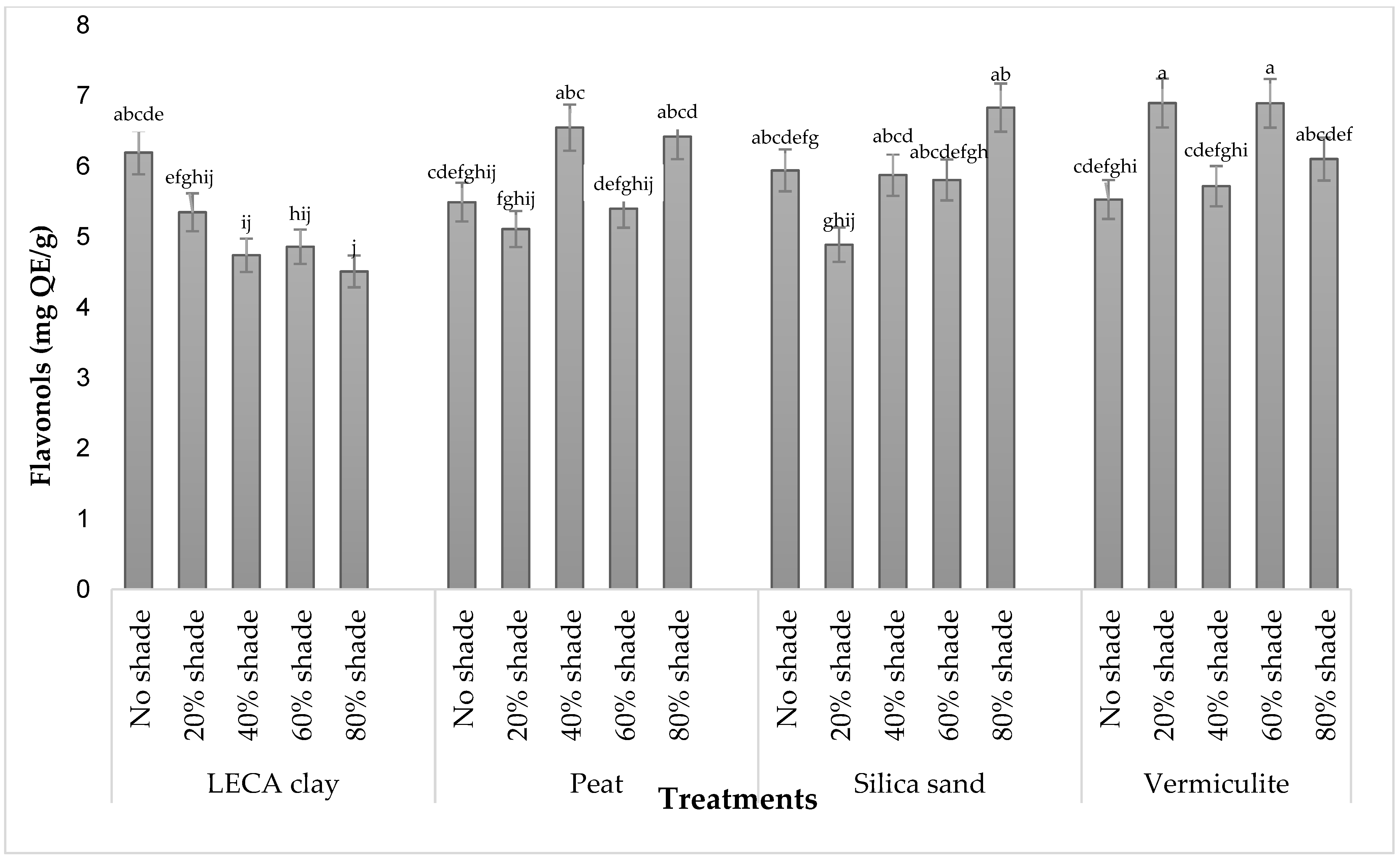
3.4.2. Total Flavonols
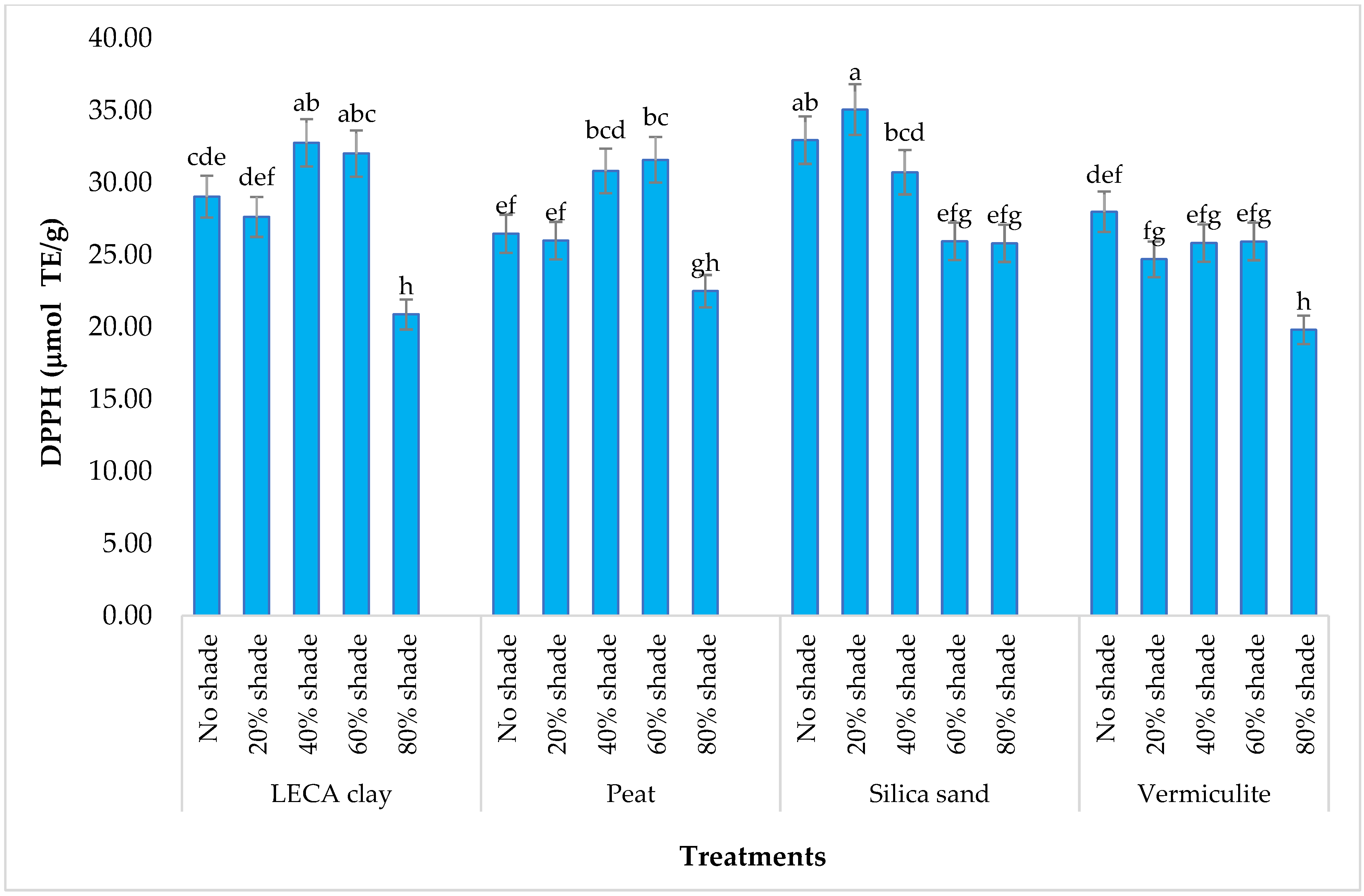
3.4.3. DPPH Antioxidant Content
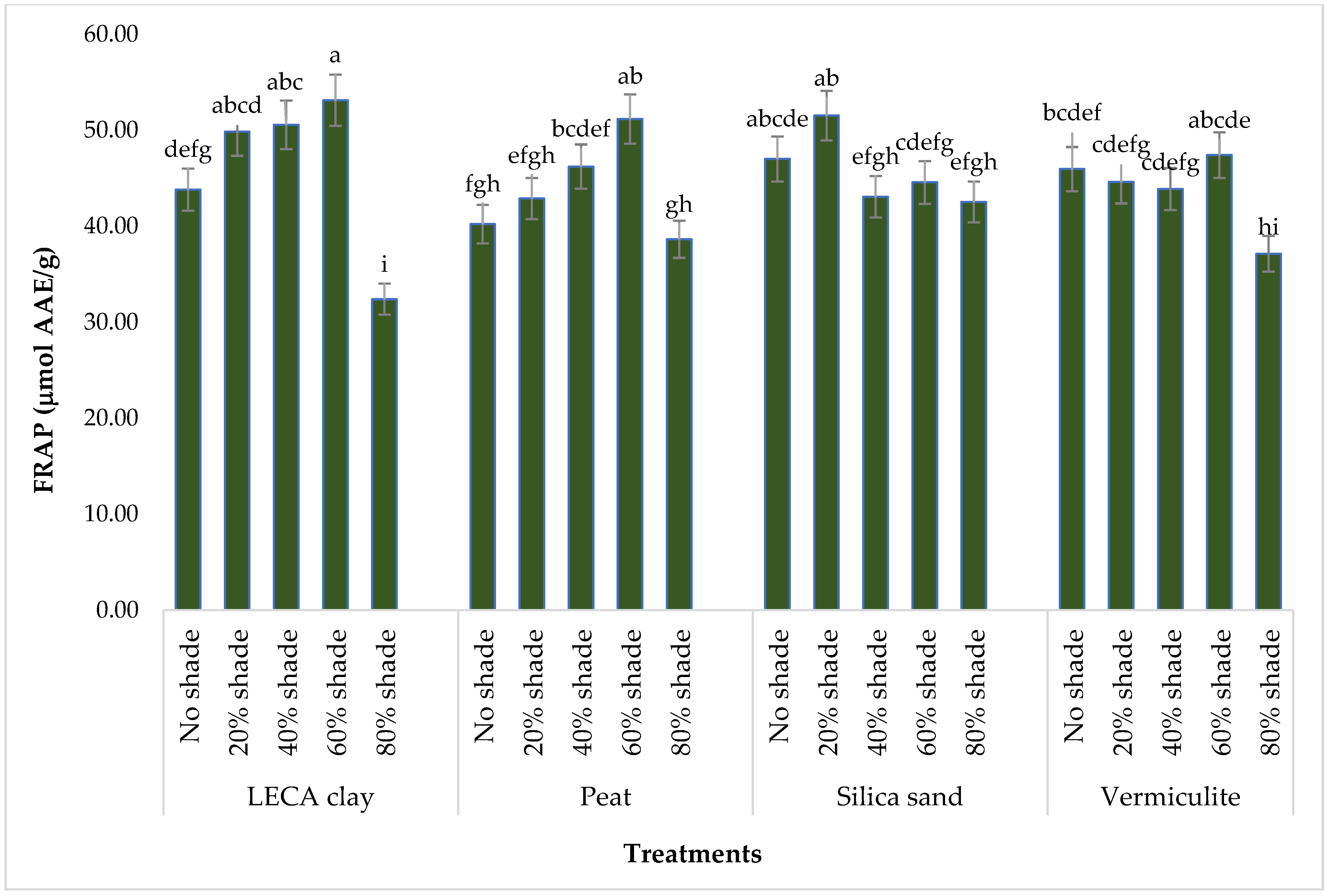
3.4.4. FRAP Antioxidant Content
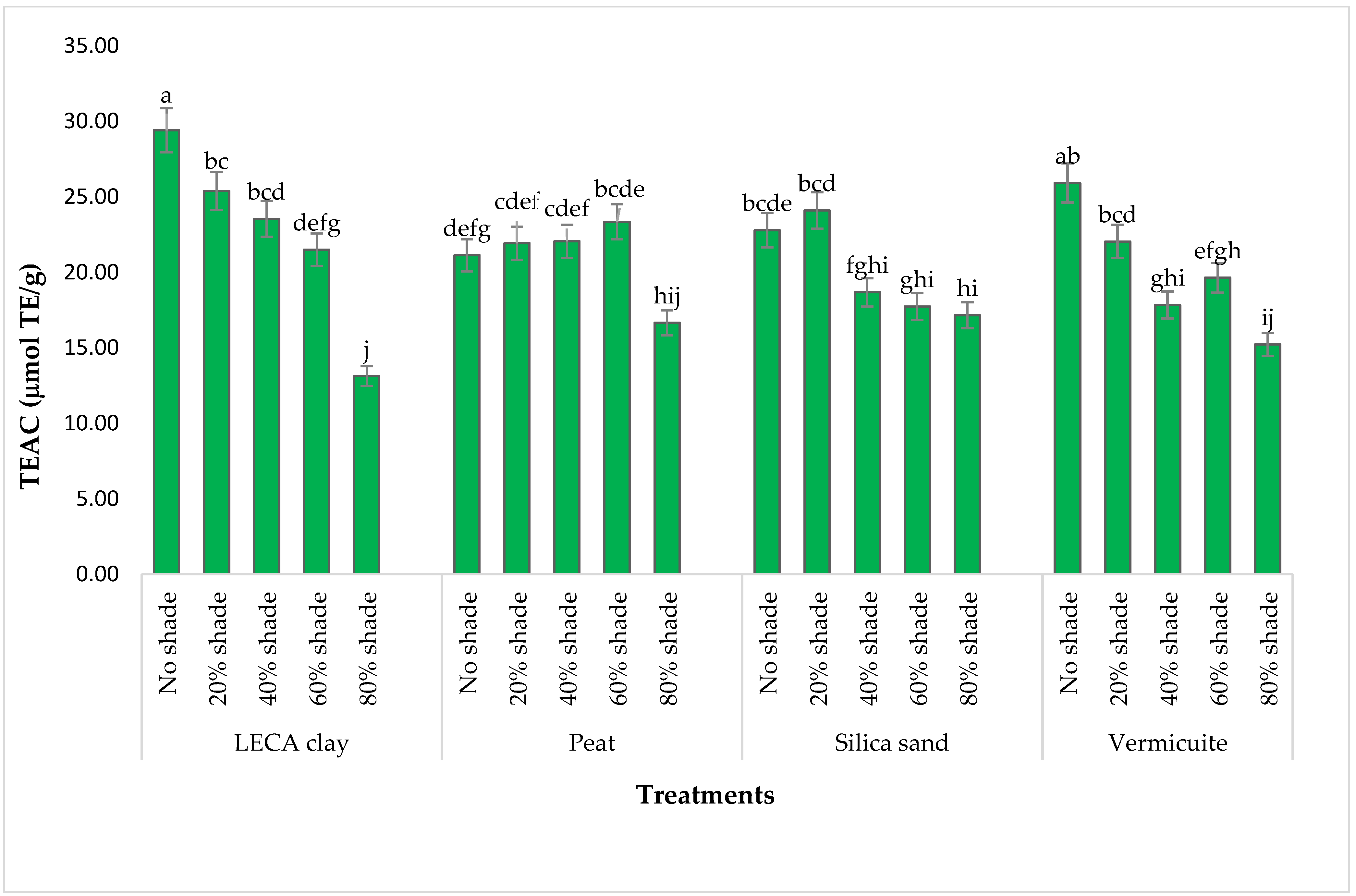
3.4.5. TEAC/ABTS Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, S.; Kumar, V.; Kapoor, D.; Kumar, S.; Singh, S.; Dhanjal, D.S.; Datta, S.; Samuel, J.; Dey, P.; Wang, S.; et al. Revealing on Hydrogen Sulfide and Nitric Oxide Signals Co-Ordination for Plant Growth under Stress Conditions. Physiol. Plant 2020, 168, 301–317. [Google Scholar] [CrossRef] [Green Version]
- Zahid, G.; Iftikhar, S.; Shimira, F.; Ahmad, H.M.; Aka Kaçar, Y. An Overview and Recent Progress of Plant Growth Regulators (PGRs) in the Mitigation of Abiotic Stresses in Fruits: A Review. Sci. Hortic. 2023, 309, 111621. [Google Scholar] [CrossRef]
- Huang, L.Z.; Zhou, M.; Ding, Y.F.; Zhu, C. Gene Networks Involved in Plant Heat Stress Response and Tolerance. Int. J. Mol. Sci. 2022, 23, 11970. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Webster, S.; He, S.Y. Growth–Defense Trade-Offs in Plants. Curr. Biol. 2022, 32, R634–R639. [Google Scholar] [CrossRef] [PubMed]
- Hazrati, S.; Tahmasebi-Sarvestani, Z.; Modarres-Sanavy, S.A.M.; Mokhtassi-Bidgoli, A.; Nicola, S. Effects of Water Stress and Light Intensity on Chlorophyll Fluorescence Parameters and Pigments of Aloe vera L. Plant Physiol. Biochem. 2016, 106, 141–148. [Google Scholar] [CrossRef]
- Ncise, W.; Daniels, C.W.; Nchu, F. Effects of Light Intensities and Varying Watering Intervals on Growth, Tissue Nutrient Content and Antifungal Activity of Hydroponic Cultivated Tulbaghia Violacea L. under Greenhouse Conditions. Heliyon 2020, 6, e03906. [Google Scholar] [CrossRef]
- Franklin, K.A. Light and Temperature Signal Crosstalk in Plant Development. Curr. Opin. Plant Biol. 2009, 12, 63–68. [Google Scholar] [CrossRef]
- Yang, L.Y.; Wang, L.T.; Ma, J.H.; Ma, E.D.; Li, J.Y.; Gong, M. Effects of Light Quality on Growth and Development, Photosynthetic Characteristics and Content of Carbohydrates in Tobacco (Nicotiana Tabacum L.) Plants. Photosynth. 2017, 55, 467–477. [Google Scholar] [CrossRef]
- Feng, L.; Raza, M.A.; Li, Z.; Chen, Y.; Bin Khalid, M.H.; Du, J.; Liu, W.; Wu, X.; Song, C.; Yu, L.; et al. The Influence of Light Intensity and Leaf Movement on Photosynthesis Characteristics and Carbon Balance of Soybean. Front. Plant Sci. 2019, 9, 1952. [Google Scholar] [CrossRef]
- Costa, L.C.D.B.; Pinto, J.E.B.P.; de Castro, E.M.; Alves, E.; Bertolucci, S.K.V.; Rosal, L.F. Effects of Coloured Shade Netting on the Vegetative Development and Leaf Structure of Ocimum selloi. Bragantia 2010, 69, 349–359. [Google Scholar] [CrossRef]
- Ptushenko, O.S.; Ptushenko, V.v.; Solovchenko, A.E. Spectrum of Light as a Determinant of Plant Functioning: A Historical Perspective. Life 2020, 10, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaitzoglou, P.; van Ieperen, W.; Harbinson, J.; van der Meer, M.; Martinakos, S.; Weerheim, K.; Nicole, C.C.S.; Marcelis, L.F.M. Effects of Continuous or End-of-Day Far-Red Light on Tomato Plant Growth, Morphology, Light Absorption, and Fruit Production. Front. Plant Sci. 2019, 10, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Sharma, A.; León, F.; Avery, B.; Kjelgren, R.; McCurdy, C.R.; Pearson, B.J. Plant Growth and Phytoactive Alkaloid Synthesis in Kratom [Mitragyna speciosa (Korth.)] in Response to Varying Radiance. PLoS ONE 2022, 17, e0259326. [Google Scholar] [CrossRef] [PubMed]
- Tran, T. The Effect of Light Exposure on the Total Chlorophyll Content, Chl a/b Ratio, and Car/Chl Ratio in the Barks of Fraxinus Latifolia Seedlings; Portland State University: Portland, OR, USA, 2018. [Google Scholar]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Germination Response of Amaranthus Caudatus L. to Soil Types and Environmental Conditions. Thaiszia J. Bot. 2019, 29, 085–100. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Ashraf, U.; Mo, Z.; Suo, H.; Li, G. Dynamics of Seed Germination, Seedling Growth and Physiological Responses of Sweet Corn under PEG-Induced Water Stress. Pak. J. Bot. 2017, 49, 639–646. [Google Scholar]
- Ngxabi, S.; Jimoh, M.O.; Kambizi, L.; Laubscher, C.P. Edibility of Trachyandra ciliata (L.F.) Kunth- An Underutilized Vegetable from South African Western Cape Coast: A Review. Afr. J. FoodAgric. Nutr. Dev. 2021, 21, 18644–18661. [Google Scholar] [CrossRef]
- Faber, R.J.; Laubscher, C.P.; Rautenbach, F.; Jimoh, M.O. Variabilities in Alkaloid Concentration of Sceletium Tortuosum (L.) N.E. Br in Response to Different Soilless Growing Media and Fertigation Regimes in Hydroponics. Heliyon 2020, 6, e05479. [Google Scholar] [CrossRef] [PubMed]
- Ngxabi, S.; Jimoh, M.O.; Kambizi, L.; Laubscher, C.P. Growth Characteristics, Phytochemical Contents, and Antioxidant Capacity of Trachyandra ciliata (L.f) Kunth Grown in Hydroponics under Varying Degrees of Salinity. Horticulturae 2021, 7, 244. [Google Scholar] [CrossRef]
- Bulawa, B.; Sogoni, A.; Jimoh, M.O.; Laubscher, C.P. Potassium Application Enhanced Plant Growth, Mineral Composition, Proximate and Phytochemical Content in Trachyandra divaricata Kunth (Sandkool). Plants 2022, 11, 3183. [Google Scholar] [CrossRef]
- Burchi, F.; Fanzo, J.; Frison, E. The Role of Food and Nutrition System Approaches in Tackling Hidden Hunger. Int. J. Environ. Res. Public Health 2011, 8, 358–373. [Google Scholar] [CrossRef]
- Noort, M.W.J.; Renzetti, S.; Linderhof, V.; du Rand, G.E.; Marx-Pienaar, N.J.M.M.; de Kock, H.L.; Magano, N.; Taylor, J.R.N. Towards Sustainable Shifts to Healthy Diets and Food Security in Sub-Saharan Africa with Climate-Resilient Crops in Bread-Type Products: A Food System Analysis. Foods 2022, 11, 135. [Google Scholar] [CrossRef] [PubMed]
- Chivenge, P.; Mabhaudhi, T.; Modi, A.T.; Mafongoya, P. The Potential Role of Neglected and Underutilised Crop Species as Future Crops under Water Scarce Conditions in Sub-Saharan Africa. Int. J. Environ. Res. Public Health 2015, 12, 5685–5711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bvenura, C.; Sivakumar, D. The Role of Wild Fruits and Vegetables in Delivering a Balanced and Healthy Diet. Food Res. Int. 2017, 99, 15–30. [Google Scholar] [CrossRef] [PubMed]
- De Vynck, J.C.; van Wyk, B.E.; Cowling, R.M. Indigenous Edible Plant Use by Contemporary Khoe-San Descendants of South Africa’s Cape South Coast. South Afr. J. Bot. 2016, 102, 60–69. [Google Scholar] [CrossRef]
- Van Wyk, B.E. The Potential of South African Plants in the Development of New Food and Beverage Products. South Afr. J. Bot. 2011, 77, 857–868. [Google Scholar] [CrossRef] [Green Version]
- Botha, M.S.; Cowling, R.M.; de Vynck, J.C.; Esler, K.J.; Potts, A.J. The Response of Geophytes to Continuous Human Foraging on the Cape South Coast, South Africa and Its Implications for Early Hunter-Gatherer Mobility Patterns. PeerJ 2022, 10, e13066. [Google Scholar] [CrossRef]
- Shew, A.M.; Tack, J.B.; Nalley, L.L.; Chaminuka, P. Yield Reduction under Climate Warming Varies among Wheat Cultivars in South Africa. Nat. Commun 2020, 11, 4408 . [Google Scholar] [CrossRef]
- Salami, S.O.; Adegbaju, O.D.; Idris, O.A.; Jimoh, M.O.; Olatunji, T.L.; Omonona, S.; Orimoloye, I.R.; Adetunji, A.E.; Olusola, A.; Maboeta, M.S.; et al. South African wild fruits and vegetables under a changing climate: The implications on health and economy. S. Afr. J. Bot. 2021, 145, 13–27. [Google Scholar] [CrossRef]
- Govender, L.; Pillay, K.; Siwela, M.; Modi, A.; Mabhaudhi, T. Food and Nutrition Insecurity in Selected Rural Communities of KwaZulu-Natal, South Africa—Linking Human Nutrition and Agriculture. Int. J. Env. Res. Public Health 2017, 14, 17. [Google Scholar] [CrossRef] [Green Version]
- Viljoen, C.C.; Jimoh, M.O.; Laubscher, C.P. Studies of Vegetative Growth, Inflorescence Development and Eco-Dormancy Formation of Abscission Layers in Streptocarpus formosus (Gesneriaceae). Horticulturae 2021, 7, 120. [Google Scholar] [CrossRef]
- Silber, A. Chemical Characteristics of Soilless Media. In Soilless Culture: Theory and Practice, 2nd ed.; Raviv, M., Lieth, J.H., Bar-Tal, A., Eds.; Academic Press: London, UK, 2019; pp. 113–148. [Google Scholar] [CrossRef]
- Bar-Tal, A.; Saha, U.K.; Raviv, M.; Tuller, M. Inorganic and Synthetic Organic Components of Soilless Culture and Potting Mixtures. In Soilless Culture: Theory and Practice, 2nd ed.; Raviv, M., Lieth, J.H., Bar-Tal, A., Eds.; Academic Press: London, UK, 2019; pp. 259–301. [Google Scholar] [CrossRef]
- Kazemi, F.; Mohorko, R. Review on the Roles and Effects of Growing Media on Plant Performance in Green Roofs in World Climates. Urban Urban Green 2017, 23, 13–26. [Google Scholar] [CrossRef]
- Mlih, R.; Bydalek, F.; Klumpp, E.; Yaghi, N.; Bol, R.; Wenk, J. Light-Expanded Clay Aggregate (LECA) as a Substrate in Constructed Wetlands–A Review. Ecol. Eng. 2020, 148, 105783. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, T.; Huang, Q.; Luo, Z.; Lu, A.; Zhu, L. Preparation, Properties Characterization and Structure Formation Mechanism of Silica Sand Tailings-Based Ceramic Materials. Mater. Chem. Phys. 2020, 255, 123611. [Google Scholar] [CrossRef]
- Katz, O.; Puppe, D.; Kaczorek, D.; Prakash, N.B.; Schaller, J. Silicon in the Soil–Plant Continuum: Intricate Feedback Mechanisms within Ecosystems. Plants 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Euliss, K.W.; Dorsey, B.L.; Benke, K.C.; Banks, M.K.; Schwab, A.P. The Use of Plant Tissue Silica Content for Estimating Transpiration. Ecol. Eng. 2005, 25, 343–348. [Google Scholar] [CrossRef]
- Kremenetskaya, I.; Ivanova, L.; Chislov, M.; Zvereva, I.; Vasilieva, T.; Marchevskaya, V.; Semushin, V.; Slukovskaya, M. Physicochemical Transformation of Expanded Vermiculite after Long-Term Use in Hydroponics. Appl. Clay Sci. 2020, 198, 105839. [Google Scholar] [CrossRef]
- Kern, J.; Tammeorg, P.; Shanskiy, M.; Sakrabani, R.; Knicker, H.; Kammann, C.; Tuhkanen, E.M.; Smidt, G.; Prasad, M.; Tiilikkala, K.; et al. Synergistic Use of Peat and Charred Material in Growing Media–an Option to Reduce the Pressure on Peatlands? J. Environ. Eng. Landsc. Manag. 2017, 25, 25–160. [Google Scholar] [CrossRef] [Green Version]
- Simons, K.L.; Ansar, A.; Kadali, K.; Bueti, A.; Adetutu, E.M.; Ball, A.S. Investigating the Effectiveness of Economically Sustainable Carrier Material Complexes for Marine Oil Remediation. Bioresour. Technol. 2012, 126, 202–207. [Google Scholar] [CrossRef]
- Brown, P.A.; Gill, S.A.; Allen, S.J. Metal Removal from Wastewater Using Peat. Water. Res. 2000, 34, 3907–3916. [Google Scholar] [CrossRef]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Nutrients and Antinutrient Constituents of Amaranthus caudatus L. Cultivated on Different Soils. Saudi. J. Biol. Sci. 2020, 27, 3570–3580. [Google Scholar] [CrossRef] [PubMed]
- Sogoni, A. The Effect of Salinity and Substrates on the Growth Parameters and Antioxidant Potential of Tetragonia decumbens (Dune Spinach) for Horticultural Applications. Master’s Dissertation, Cape Peninsula University of Technology, Cape Town, South Africa, 2020. [Google Scholar]
- Olatunji, T.L.; Afolayan, A.J. Comparison of nutritional, antioxidant vitamins and capsaicin contents in Capsicum annuum and C. frutescens. Int. J. Veg. Sci. 2020, 26, 190–207. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemist). Official Methods of Analysis; Latimer, G.W., Jr., Ed.; AOAC International: Washinton DC, USA, 2016; ISBN 0935584870. [Google Scholar]
- Idris, O.A.; Wintola, O.A.; Afolayan, A.J. Comparison of the Proximate Composition, Vitamins (Ascorbic Acid, α-Tocopherol and Retinol), Anti-Nutrients (Phytate and Oxalate) and the GC-MS Analysis of the Essential Oil of the Root and Leaf of Rumex crispus L. Plants 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sogoni, A.; Jimoh, M.O.; Kambizi, L.; Laubscher, C.P. The Impact of Salt Stress on Plant Growth, Mineral Composition, and Antioxidant Activity in Tetragonia decumbens Mill.: An Underutilized Edible Halophyte in South Africa. Horticulturae 2021, 7, 140. [Google Scholar] [CrossRef]
- Lim, S.H.; Kim, D.H.; Jung, J.A.; Hyung, N.I.; Youn, Y.; Lee, J.Y. Silencing of Dihydroflavonol 4-Reductase in Chrysanthemum Ray Florets Enhances Flavonoid Biosynthesis and Antioxidant Capacity. Plants 2022, 11, 1681. [Google Scholar] [CrossRef] [PubMed]
- Ohikhena, F.; Wintola, O.; Afolayan, A.J. Quantitative Phytochemical Constituents and Antioxidant Activities of the Mistletoe, Phragmanthera capitata (Sprengel) Balle Extracted with Different Solvents. Pharmacogn. Res. 2018, 10, 16–23. [Google Scholar] [CrossRef]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Antioxidant and Phytochemical Activities of Amaranthus caudatus L. Harvested from Different Soils at Various Growth Stages. Sci. Rep. 2019, 9, 12965. [Google Scholar] [CrossRef] [Green Version]
- Jimoh, M.A.; Idris, O.A.; Jimoh, M.O. Cytotoxicity, Phytochemical, Antiparasitic Screening, and Antioxidant Activities of Mucuna pruriens (Fabaceae). Plants 2020, 9, 1249. [Google Scholar] [CrossRef]
- Feller, U.; Rahman, S.; Marinello, F.; Lara, I.; Ortiz, R.; Bacenetti, J.; Cecchini, M.; Andolfi, A.; Ho Kim, K.; Rayhan Ahmed Shawon, M.; et al. Effect of Shade Screen on Sap Flow, Chlorophyll Fluorescence, NDVI, Plant Growth and Fruit Characteristics of Cultivated Paprika in Greenhouse. Agriculture 2022, 12, 1405. [Google Scholar] [CrossRef]
- Appolloni, E.; Pennisi, G.; Zauli, I.; Carotti, L.; Paucek, I.; Quaini, S.; Orsini, F.; Gianquinto, G. Beyond Vegetables: Effects of Indoor LED Light on Specialized Metabolite Biosynthesis in Medicinal and Aromatic Plants, Edible Flowers, and Microgreens. J. Sci. Food Agric. 2022, 102, 472–487. [Google Scholar] [CrossRef]
- Kitir, N.; Yildirim, E.; Şahin, Ü.; Turan, M.; Ekinci, M.; Ors, S.; Kul, R.; Ünlü, H.; Ünlü, H. Peat Use in Horticulture. In Peat; Topcuoğlu, B., Turan, M., Eds.; IntechOpen: London, UK, 2018; pp. 75–90. [Google Scholar] [CrossRef] [Green Version]
- Shao, Q.; Wang, H.; Guo, H.; Zhou, A.; Huang, Y.; Sun, Y.; Li, M. Effects of Shade Treatments on Photosynthetic Characteristics, Chloroplast Ultrastructure, and Physiology of Anoectochilus roxburghii. PLoS ONE 2014, 9, e85996. [Google Scholar] [CrossRef]
- Ogundola, A.F.; Bvenura, C.; Afolayan, A.J. Nutrient and Antinutrient Compositions and Heavy Metal Uptake and Accumulation in S. nigrum Cultivated on Different Soil Types. Sci. World J. 2018, Article ID 5703929, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Cope, D.G. Nursing Considerations for Androgen Deprivation Therapy and Bone Health in Men Affected by Prostate Cancer. Semin. Oncol. Nurs. 2022, 38, 151271. [Google Scholar] [CrossRef]
- Diviš, P.; Pořízka, J.; Vespalcov, M.; Matějíček, A.; Kaplan, J. Elemental Composition of Fruits from Different Black Elder (Sambucus nigra L.) Cultivars Grown in the Czech Republic. J. Elem. 2015, 20, 549–557. [Google Scholar] [CrossRef]
- Achigan-Dako, E.G.; Sogbohossou, O.E.D.; Maundu, P. Current Knowledge on Amaranthus Spp.: Research Avenues for Improved Nutritional Value and Yield in Leafy Amaranths in Sub-Saharan Africa. Euphytica 2014, 197, 303–317. [Google Scholar] [CrossRef]
- Gostin, A.I.; Waisundara, V.Y. Edible Flowers as Functional Food: A Review on Artichoke (Cynara cardunculus L.). Trends Food Sci. Technol. 2019, 86, 381–391. [Google Scholar] [CrossRef]
- Lau, B.F.; Kong, K.W.; Leong, K.H.; Sun, J.; He, X.; Wang, Z.; Mustafa, M.R.; Ling, T.C.; Ismail, A. Banana Inflorescence: Its Bio-Prospects as an Ingredient for Functional Foods. Trends Food Sci. Technol. 2020, 97, 14–28. [Google Scholar] [CrossRef]
- Li, H.; Xia, Y.; Liu, H.Y.; Guo, H.; He, X.Q.; Liu, Y.; Wu, D.T.; Mai, Y.H.; Li, H.-B.; Zou, L.; et al. Nutritional Values, Beneficial Effects, and Food Applications of Broccoli (Brassica oleracea Var. Italica Plenck). Trends Food Sci. Technol. 2022, 119, 288–308. [Google Scholar] [CrossRef]
- Barbagallo, M.; Veronese, N.; Dominguez, L.J. Magnesium in Aging, Health and Diseases. Nutrients 2021, 13, 463. [Google Scholar] [CrossRef]
- Serna, J.; Bergwitz, C. Importance of Dietary Phosphorus for Bone Metabolism and Healthy Aging. Nutrients 2020, 12, 3001. [Google Scholar] [CrossRef]
- Guo, Q.; Wang, N.; Liu, H.; Li, Z.; Lu, L.; Wang, C. The Bioactive Compounds and Biological Functions of Asparagus officinalis L.–A Review. J. Funct. Foods 2020, 65, 103727. [Google Scholar] [CrossRef]
- Navarro-González, I.; González-Barrio, R.; García-Valverde, V.; Bautista-Ortín, A.B.; Periago, M.J. Nutritional Composition and Antioxidant Capacity in Edible Flowers: Characterisation of Phenolic Compounds by HPLC-DAD-ESI/MSn. Int. J. Mol. Sci. 2014, 16, 805–822. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Arrieta, R.; Esquivel-Alvarado, D.; Alfaro-Viquez, E.; Alvarez-Valverde, V.; Krueger, C.G.; Reed, J.D. Nutritional and Bioactive Composition of Spanish, Valencia, and Virginia Type Peanut Skins. J. Food Compos. Anal. 2021, 98, 103816. [Google Scholar] [CrossRef]
- Fernandes, L.; Casal, S.; Pereira, J.A.; Saraiva, J.A.; Ramalhosa, E. Edible Flowers: A Review of the Nutritional, Antioxidant, Antimicrobial Properties and Effects on Human Health. J. Food Compos. Anal. 2017, 60, 38–50. [Google Scholar] [CrossRef]
- Radha; Kumar, M.; Puri, S.; Pundir, A.; Bangar, S.P.; Changan, S.; Choudhary, P.; Parameswari, E.; Alhariri, A.; Samota, M.K.; et al. Evaluation of Nutritional, Phytochemical, and Mineral Composition of Selected Medicinal Plants for Therapeutic Uses from Cold Desert of Western Himalaya. Plants 2021, 10, 1429. [Google Scholar] [CrossRef]
- Schroeder, B.O.; Bäckhed, F. Signals from the Gut Microbiota to Distant Organs in Physiology and Disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef]
- Kumar, M.; Zhang, B.; Nishad, J.; Verma, A.; Sheri, V.; Dhumal, S.; Radha; Sharma, N.; Chandran, D.; Senapathy, M.; et al. Jamun (Syzygium cumini (L.) Skeels) Seed: A Review on Nutritional Profile, Functional Food Properties, Health-Promoting Applications, and Safety Aspects. Processes 2022, 10, 2169. [Google Scholar] [CrossRef]
- Sotelo, A.; López-García, S.; Basurto-Peña, F. Content of Nutrient and Antinutrient in Edible Flowers of Wild Plants in Mexico. Plant Foods Hum. Nutr. 2007, 62, 133–138. [Google Scholar] [CrossRef]
- Pinto, T.; Vilela, A.; Cosme, F. Chemical and Sensory Characteristics of Fruit Juice and Fruit Fermented Beverages and Their Consumer Acceptance. Beverages 2022, 8, 33. [Google Scholar] [CrossRef]
- Akbari, B.; Baghaei-Yazdi, N.; Bahmaie, M.; Mahdavi Abhari, F. The Role of Plant-Derived Natural Antioxidants in Reduction of Oxidative Stress. BioFactors 2022, 48, 611–633. [Google Scholar] [CrossRef]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of Light Quality on the Accumulation of Phytochemicals in Vegetables Produced in Controlled Environments: A Review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef]
- Gao, M.; He, R.; Shi, R.; Zhang, Y.; Song, S.; Su, W.; Liu, H. Differential Effects of Low Light Intensity on Broccoli Microgreens Growth and Phytochemicals. Agronomy 2021, 11, 537. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NFT/Table | Gutter 1 | Gutter 2 | Gutter 3 | Gutter 4 | Gutter 5 |
|---|---|---|---|---|---|
| 1 | Leca clay + no shade | Leca clay + 20% shade | Leca clay + 40% shade | Leca clay + 60% shade | Leca clay + 80% shade |
| 2 | Peat + no shade | Peat + 20% shade | Peat + 40% shade | Peat + 60% shade | Peat + 80% shade |
| 3 | Sand + no shade (C) | Sand + 20% shade | Sand + 40% shade | Sand + 60% shade | Sand + 80% shade |
| 4 | Vermiculite + no shade | Vermiculite + 20% shade | Vermiculite + 40% shade | Vermiculite + 60% shade | Vermiculite + 80% shade |
| Media | Shade | Week 2 | Week 4 | Week 6 | Week 8 | Week 10 | Week 12 |
|---|---|---|---|---|---|---|---|
| No shade | 1.020 ± 0.25 c–g | 1.026 ± 0.41 d–g | 0.99 ± 0.02 d | 1.09 ± 0.23 b–e | 1.19 ± 0.02 ab | 1.15 ± 0.03 ab | |
| 20% shade | 0.961 ± 0.34 fg | 0.99 ± 0.37 efg | 1.32 ± 0.05 ab | 1.00 ± 0.02 cde | 1.27 ± 0.05 a | 1.02 ± 0.02 ab | |
| LECA clay | 40% shade | 0.980 ± 0.02 d–g | 1.02 ± 0.02 d-g | 1.09 ± 0.03 cd | 1.12 ± 0.26 ab | 1.06 ± 0.03 abc | 1.12 ± 0.03 ab |
| 60% shade | 1.20 ± 0.27 ab | 1.35 ± 0.03 ab | 1.41 ± 0.04 ab | 1.35 ± 0.04 a | 1.12 ± 0.02 ab | 1.03 ± 0.04 ab | |
| 80% shade | 1.32 ± 0.49 a | 1.34 ± 0.05 a | 1.43 ± 0.04 a | 0.96 ± 0.10 de | 0.96 ± 0.12 abc | 1.20 ± 0.14 ab | |
| No shade | 1.14 ± 0.02 b–e | 0.95 ± 0.02 g | 1.00 ± 0.02 d | 1.06 ± 0.03 b-e | 1.03 ± 0.19 abc | 0.99 ± 0.04 b | |
| 20% shade | 1.11 ± 0.03 b-f | 1.20 ± 0.07 bcd | 1.10 ± 0.04 cd | 1.09 ± 0.03 b-e | 1.14 ± 0.06 ab | 1.02 ± 0.04 ab | |
| Peat | 40% shade | 0.97 ± 0.02 efg | 1.04 ± 0.02 d-g | 1.11 ± 0.03 cd | 1.09 ± 0.02 b-e | 1.08 ± 0.04 abc | 1.16 ± 0.08 ab |
| 60% shade | 1.15 ± 0.02 a-d | 1.28 ± 0.05 abc | 1.40 ± 0.06 ab | 1.15 ± 0.01 a–d | 1.16 ± 0.03 ab | 1.11 ± 0.02 ab | |
| 80% shade | 1.17 ± 0.02 abc | 1.26 ± 0.04 abc | 1.19 ± 0.06 d | 1.11 ± 0.04 b–e | 1.19 ± 0.04 ab | 1.15 ± 0.13 ab | |
| No shade | 0.96 ± 0.03 fg | 1.00 ± 0.03 efg | 1.03 ± 0.03 d | 1.21 ± 0.03 abc | 1.16 ± 0.03 ab | 0.98 ± 0.03 b | |
| 20% shade | 1.13 ± 0.02 b–e | 0.99 ± 0.02 efg | 1.04 ± 0.02 d | 1.08 ± 0.03 b–e | 1.08 ± 0.03 abc | 1.01 ± 0.02 b | |
| Silica sand | 40% shade | 1.02 ± 0.03 c–g | 1.04 ± 0.02 d-g | 1.02 ± 0.03 d | 0.90 ± 0.04 e | 1.00 ± 0.12 bc | 0.88 ± 0.03 b |
| 60% shade | 1.17 ± 0.03 abc | 1.16 ± 0.05 b-e | 1.35 ± 0.04 ab | 1.25 ± 0.03 ab | 1.97 ± 0.22 ab | 1.13 ± 0.02 ab | |
| 80% shade | 1.23 ± 0.05 ab | 1.24 ± 0.03 abc | 1.33 ± 0.04 ab | 1.15 ± 0.13 a–d | 1.22 ± 0.14 ab | 1.29 ± 0.03 ab | |
| No shade | 1.00 ± 0.04 c–g | 1.14 ± 0.03 c-f | 1.11 ± 0.03 cd | 1.12 ± 0.02 b–e | 1.04 ± 0.02 abc | 1.16 ± 0.03 ab | |
| 20% shade | 0.99 ± 0.02 efg | 1.27 ± 0.05 abc | 1.10 ± 0.04 cd | 1.13 ± 0.03 a–e | 0.81 ± 0.21 c | 0.84 ± 0.18 b | |
| Vermiculite | 40% shade | 0.89 ± 0.04 g | 0.96 ± 0.02 fg | 1.01 ± 0.01 d | 0.98 ± 0.03 cde | 1.08 ± 0.03 abc | 1.07 ± 0.03 ab |
| 60% shade | 1.09 ± 0.04 b–f | 1.27 ± 0.05 abc | 1.24 ± 0.04 bc | 1.14 ± 0.03 a–d | 1.08 ± 0.02 abc | 1.04 ± 0.02 ab | |
| 80% shade | 1.22 ± 0.07 ab | 1.26 ± 0.05 abc | 1.09 ± 0.02 cd | 1.14 ± 0.02 a–d | 1.24 ± 0.04 ab | 1.42 ± 0.02 a | |
| Analysis of Variance | |||||||
| Growth media | 0.004 | 0.001 | 0.000 | 0.642 ns | 0.394 ns | 0.171 | |
| Shade levels | 0.000 | 0.000 | 0.000 | 0.000 | 0.394 ns | 0.000 | |
| Growth media × shade levels | 0.000 | 0.000 | 0.000 | 0.000 | 0.442 ns | 0.010 | |
| Shade | Media | Calcium | K/Ca + Mg | Potassium | Nitrogen | Magnesium | Phosphorus | Sodium |
|---|---|---|---|---|---|---|---|---|
| mg/100 g | mg/100 g | mg/100 g | mg/100 g | mg/100 g | mg/100 g | mg/100 g | ||
| No shade | LECA clay | 409.5 ± 0.5 i | 3145.0 ± 5.0 e | 5865.0 ± 5.0 h | 3019.0 ± 1.0 k | 324.5 ± 5.5 fg | 511.0 ± 1.0 ef | 139.0 ± 1.0 jk |
| Peat | 409.5 ± 0.5 i | 3419.5 ± 0.5 cd | 5926.5 ± 3.5 g | 2880.5 ± 0.5 n | 289.0 ± 1.0 h | 529.0 ± 1.0 e | 157.5 ± 2.5 gh | |
| Silica sand | 347.5 ± 2.5 m | 3079.0 ± 1.0 h | 4979.0 ± 1.0 m | 3257.5 ± 2.5 g | 286.0 ± 3.5 h | 444.5 ± 5.5 g | 169.0 ± 1.0 e | |
| Vermiculite | 327.5 ± 2.5 o | 3039.0 ± 1.0 j | 5570.0 ± 10.0 j | 2957.0 ± 2.5 l | 371.0 ± 1.0 cde | 489.0 ± 1.0 f | 137.5 ± 2.5 k | |
| 20% shade | LECA clay | 389.5 ± 0.5 k | 4039.0 ± 1.0 a | 6329.0 ± 1.0 d | 3579.0 ± 1.0 d | 249.0 ± 1.0 i | 427.5 ± 2.5 g | 147.5 ± 2.5 ij |
| Peat | 469.5 ± 0.5 f | 3619.0 ± 0.5 b | 7167.5 ± 2.5 a | 2867.5 ± 2.5 o | 329.0 ± 1.0 fg | 689.0 ± 1.0 bc | 149.0 ± 1.0 hi | |
| Silica sand | 347.5 ± 2.5 m | 3147.5 ± 2.5 e | 5390.5 ± 0.5 k | 3119.0 ± 1.0 h | 319.0 ± 1.0 gh | 511.5 ± 1.5 ef | 144.0 ± 4.0 ijk | |
| Vermiculite | 449.0 ± 1.0 h | 2839.0 ± 1.0 m | 6319.5 ± 0.5 d | 2939.0 ± 1.0 m | 419.0 ± 1.0 b | 528.5 ± 1.5 e | 149.0 ± 1.0 hi | |
| 40% shade | LECA clay | 689.0 ± 1.0 d | 1979.0 ± 1.0 p | 4961.0 ± 1.0 m | 2960.5 ± 0.5 l | 359.0 ± 1.0 def | 611.0 ± 1.0 d | 179.0 ± 1.0 d |
| Peat | 519.0 ± 1.0 e | 3019.5 ± 0.5 k | 6648.0 ± 2.0 b | 3047.5 ± 2.5 j | 369.5 ± 0.5 cde | 678.5 ± 1.5 c | 158.0 ± 2.0 gh | |
| Silica sand | 829.5 ± 0.5 c | 2219.0 ± 1.0 o | 6169.0 ± 1.0 f | 2720.0 ± 1.0 q | 357.5 ± 2.5 def | 419.0 ± 1.0 g | 230.5 ± 0.5 a | |
| Vermiculite | 409.0 ± 1.0 i | 2519.5 ± 0.5 n | 5239.5 ± 20.5 l | 3119.0 ± 1.0 h | 399.0 ± 1.0 bc | 748.5 ± 2.5 a | 147.5 ± 2.5 ij | |
| 60% shade | LECA clay | 359.5 ± 0.5 l | 2979.0 ± 1.0 l | 4865.0 ± 5.0 n | 2811.0 ± 1.0 p | 289.0 ± 1.0 h | 611.0 ± 1.0 d | 138.5 ± 1.5 jk |
| Peat | 458.5 ± 1.5 g | 3111.0 ± 1.0 f | 6189.0 ± 1.0 ef | 3412.0 ± 2.5 e | 337.5 ± 2.5 efg | 747.5 ± 2.5 a | 159.0 ± 1.0 fg | |
| Silica sand | 409.0 ± 1.0 i | 3428.0 ± 2.0 c | 6389.0 ± 1.0 c | 3599.0 ± 1.0 c | 328.0 ± 1.5 fg | 744.0 ± 26.0 a | 168.0 ± 2.0 ef | |
| Vermiculite | 360.5 ± 0.5 l | 3411.0 ± 1.0 d | 6223.5 ± 6.5 e | 3719.5 ± 0.5 a | 347.0 ± 3.0 defg | 716.0 ± 4.0 ab | 169.5 ± 0.5 de | |
| 80% shade | LECA clay | 1101.0 ± 1.0 a | 1536.5 ± 3.5 r | 5692.5 ± 7.5 i | 3018.5 ± 1.5 k | 478.5 ± 1.5 a | 529.0 ± 1.0 e | 219.5 ± 0.5 b |
| Peat | 339.0 ± 1.0 n | 3100.5 ± 0.5 g | 5228.0 ± 12.0 l | 3365.0 ± 5.0 f | 319.0 ± 1.0 gh | 678.5 ± 1.5 c | 79.0 ± 1.0 m | |
| Silica sand | 1059.0 ± 1.0 b | 1911.0 ± 1.0 q | 6392.5 ± 7.5 c | 3060.0 ± 0.5 p | 374.0 ± 26.0 cd | 516.0 ± 4.0 ef | 199.0 ± 1.0 c | |
| Vermiculite | 400.0 ± 1.0 j | 3069.0 ± 1.0 i | 5559.0 ± 1.0 j | 3689.0 ± 1.0 b | 318.50 ± 1.50 gh | 727.5 ± 2.5 a | 99.5 ± 0.5 l | |
| Analysis of Variance | ||||||||
| Growth media | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| Shade levels | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| Growth media × shade levels | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| Shade | Media | Copper (mg/100 g) | Iron (mg/100 g) | Manganese (mg/100 g) | Zinc (mg/100 g) |
|---|---|---|---|---|---|
| No shade | LECA clay | 0.40 ± 0.01 abcd | 7.50 ± 0.50 h | 3.69 ± 0.01 g | 5.81 ± 0.01 h |
| Peat | 0.00 ± 0.00 d | 6.25 ± 0.05 ij | 3.47 ± 0.03 g | 5.84 ± 0.06 h | |
| Silica sand | 0.40 ± 0.01 abcd | 9.25 ± 0.05 fg | 2.19 ± 0.01 h | 6.58 ± 0.02 g | |
| Vermiculite | 0.69 ± 0.01 ab | 9.45 ± 0.05 ef | 3.79 ± 0.02 g | 5.58 ± 0.02 i | |
| 20% shade | LECA clay | 0.53 ± 0.43 abcd | 5.75 ± 0.05 ij | 3.55 ± 0.30 g | 5.36 ± 0.04 j |
| Peat | 0.11 ± 0.01 cd | 5.85 ± 0.05 ij | 5.85 ± 0.05 cd | 7.58 ± 0.02 e | |
| Silica sand | 0.29 ± 0.01 bcd | 7.55 ± 0.05 h | 2.79 ± 0.02 h | 6.41 ± 0.01 g | |
| Vermiculite | 0.69 ± 0.01 ab | 15.75 ± 0.05 d | 5.48 ± 0.02 d | 6.47 ± 0.03 g | |
| 40% shade | LECA clay | 0.49 ± 0.02 abcd | 22.11 ± 0.01 c | 5.84 ± 0.05 cd | 7.59 ± 0.01 e |
| Peat | 0.00 ± 0.00 d | 6.55 ± 0.05 i | 2.80 ± 0.20 h | 6.38 ± 0.03 g | |
| Silica sand | 0.39 ± 0.01 abcd | 23.89 ± 0.01 b | 5.29 ± 0.02 de | 8.47 ± 0.03 c | |
| Vermiculite | 0.79 ± 0.01 ab | 6.20 ± 0.01 ij | 3.58 ± 0.02 g | 6.58 ± 0.02 g | |
| 60% shade | LECA clay | 0.29 ± 0.01 bcd | 6.29 ± 0.01 ij | 3.45 ± 0.35 g | 8.28 ± 0.02 c |
| Peat | 0.00 ± 0.00 d | 6.29 ± 0.01 ij | 6.47 ± 0.03 bc | 7.58 ± 0.02 e | |
| Silica sand | 0.49 ± 0.01 abcd | 8.50 ± 0.50 g | 3.93 ± 0.08 g | 6.57 ± 0.03 g | |
| Vermiculite | 0.80 ± 0.01 ab | 5.49 ± 0.01 j | 4.68 ± 0.02 ef | 7.95 ± 0.05 d | |
| 80% shade | LECA clay | 0.89 ± 0.01 a | 40.39 ± 0.01 a | 9.48 ± 0.03 a | 9.19 ± 0.01 b |
| Peat | 0.58 ± 0.03 abc | 5.68 ± 0.03 ij | 3.68 ± 0.02 g | 5.93 ± 0.07 h | |
| Silica sand | 0.58 ± 0.02 abc | 22.11 ± 0.01 c | 6.58 ± 0.03 b | 10.50 ± 0.10 a | |
| Vermiculite | 0.69 ± 0.02 ab | 10.29 ± 0.02 e | 4.65 ± 0.05 f | 7.11 ± 0.01 f | |
| Analysis of Variance | |||||
| Growth media | 0.000 | 0.000 | 0.000 | 0.000 | |
| Shade levels | 0.001 | 0.000 | 0.000 | 0.000 | |
| Growth media × shade levels | 0.056 ns | 0.000 | 0.000 | 0.000 | |
| Shade | Media | ADF % | Ash % | Fat % | Moisture % | NDF % | Protein % |
|---|---|---|---|---|---|---|---|
| No shade | LECA clay | 33.78 ± 1.16 bcd | 12.45 ± 0.15 bcd | 2.00 ± 0.05 a–e | 9.87 ± 0.51 abc | 47.93 ± 0.68 ab | 19.14 ± 0.23 bcd |
| Peat | 32.80 ± 0.29 bcd | 12.59 ± 0.66 bcd | 1.91 ± 0.03 b–f | 9.76 ± 0.31 abc | 44.3 ± 0.77 bc | 17.63 ± 0.38 cd | |
| Silica sand | 31.12 ± 0.87 def | 11.01 ± 0.32 d | 1.82 ± 0.05 bef | 10.62 ± 0.39 ab | 40.85 ± 1.23 b–e | 19.87 ± 0.51 a–d | |
| Vermiculite | 32.19 ± 0.32 cde | 11.92 ± 0.27 cd | 2.01 ± 0.02 a–e | 10.26 ± 0.70 abc | 40.55 ± 1.30 b–e | 18.04 ± 0.48 cd | |
| 20% shade | LECA clay | 35.91 ± 0.31 b | 13.76 ± 0.36 bcd | 1.73 ± 0.70 efg | 10.29 ± 0.21 abc | 44.45 ± 3.09 bc | 21.86 ± 0.50 ab |
| Peat | 41.17 ± 0,28 a | 14.97 ± 0.33 b | 1.56 ± 0.65 g | 10.32 ± 0.46 abc | 53.87 ± 0.57 a | 17.71 ± 0.26 cd | |
| Silica sand | 29.09 ± 0.25 ef | 11.91 ± 0.46 cd | 2.26 ± 0.65 ab | 10.29 ± 0.21 abc | 40.85 ± 0.53 bcde | 19.23 ± 0.24 a–d | |
| Vermiculite | 35.09 ± 1.54 bc | 14.01 ± 0.36 bc | 2.05 ± 0.55 a–d | 9.70 ± 0.15 abc | 43.98 ± 3.62 bc | 17.83 ± 0.58 cd | |
| 40% shade | LECA clay | 24.84 ± 0.18 ij | 14.34 ± 0.50 bc | 2.04 ± 0.15 a–e | 11.27 ± 0.41 a | 34.21 ± 0.65 ef | 18.02 ± 0.48 cd |
| Peat | 30.48 ± 0.41 d–g | 14.39 ± 0.37 bc | 1.90 ± 0.20 b–f | 11.12 ± 0.48 ab | 39.24 ± 1.04 cdef | 18.97 ± 0.11 bcd | |
| Silica sand | 31.67 ± 0.41 cde | 18.89 ± 0.34 a | 1.82 ± 0.45 def | 10.02 ± 0.66 abc | 39.53 ± 1.27 cdef | 16.88 ± 0.13 d | |
| Vermiculite | 28.23 ± 0.42 e–i | 13.14 ± 0.28 bcd | 2.14 ± 0.40 abc | 11.37 ± 0.38 a | 35.80 ± 1.55 def | 17.67 ± 1.80 cd | |
| 60% shade | LECA clay | 30.08 ± 0.78 d–g | 12.23 ± 0.34 bcd | 1.83 ± 0.01 def | 11.30 ± 0.52 a | 42.83 ± 0.47 bcd | 17.90 ± 0.35 cd |
| Peat | 27.81 ± 0.22 f–i | 14.38 ± 0.54 bc | 1.84 ± 0.02 c–f | 11.53 ± 0.23 a | 40.93 ± 0.67 bcde | 20.94 ± 0.38 abc | |
| Silica sand | 31.15 ± 0.49 def | 14.21 ± 0.76 bc | 1.68 ± 0.03 fg | 9.40 ± 0.75 abc | 43.78 ± 0.53 bc | 21.91 ± 0.61 ab | |
| Vermiculite | 31.94 ± 1.04 cde | 14.14 ± 0.69 bc | 1.79 ± 0.05 d–g | 8.81 ± 0.17 bc | 40.93 ± 0.57 bcde | 21.80 ± 1.43 ab | |
| 80% shade | LECA clay | 23.57 ± 1.07 j | 19.19 ± 0.54 a | 2.09 ± 0.01 a–d | 8.05 ± 0.18 cd | 32.91 ± 0.51 f | 18.26 ± 0.63 bcd |
| Peat | 24.84 ± 0.54 ij | 13.06 ± 0.50 bcd | 2.25 ± 0.02 a | 9.76 ± 0.40 abc | 32.91 ± 0.66 f | 20.65 ± 0.42 abc | |
| Silica sand | 26.31 ± 0.01 g–j | 20.86 ± 0.87 a | 1.81 ± 0.16 def | 9.88 ± 0.25 bcd | 35.81 ± 0.40 ef | 19.04 ± 0.08 bcd | |
| Vermiculite | 26.02 ± 0.52 hij | 13.76 ± 0.48 bcd | 1.93 ± 0.11 b–f | 6.25 ± 0.65 d | 34.28 ± 0.72 ef | 22.82 ± 0.26 a | |
| Analysis of Variance | |||||||
| Growth media | 0.002 | 0.000 | 0.009 | 0.001 | 0.013 | 0.493 ns | |
| Shade levels | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| Growth media × shade levels | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tshayingwe, A.; Jimoh, M.O.; Sogoni, A.; Wilmot, C.M.; Laubscher, C.P. Light Intensity and Growth Media Influence Growth, Nutrition, and Phytochemical Content in Trachyandra divaricata Kunth. Agronomy 2023, 13, 247. https://doi.org/10.3390/agronomy13010247
Tshayingwe A, Jimoh MO, Sogoni A, Wilmot CM, Laubscher CP. Light Intensity and Growth Media Influence Growth, Nutrition, and Phytochemical Content in Trachyandra divaricata Kunth. Agronomy. 2023; 13(1):247. https://doi.org/10.3390/agronomy13010247
Chicago/Turabian StyleTshayingwe, Ayakha, Muhali Olaide Jimoh, Avela Sogoni, Carolyn Margaret Wilmot, and Charles Petrus Laubscher. 2023. "Light Intensity and Growth Media Influence Growth, Nutrition, and Phytochemical Content in Trachyandra divaricata Kunth" Agronomy 13, no. 1: 247. https://doi.org/10.3390/agronomy13010247