

From Plant Nursery to Field: Persistence of Mycorrhizal Symbiosis Balancing Effects on Growth-Defence Tradeoffs Mediated by Rootstock



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. RNA Isolation and RT-qPCR Analysis
2.3. Leaf Gas Exchange Measurements
2.4. HPLC-DAD Targeted Metabolites Analysis
2.5. Statistics
3. Results
3.1. Expression Analysis of Nitrogen-Related Genes in Cuttings
3.2. Leaf Gas Exchange Performances in Field
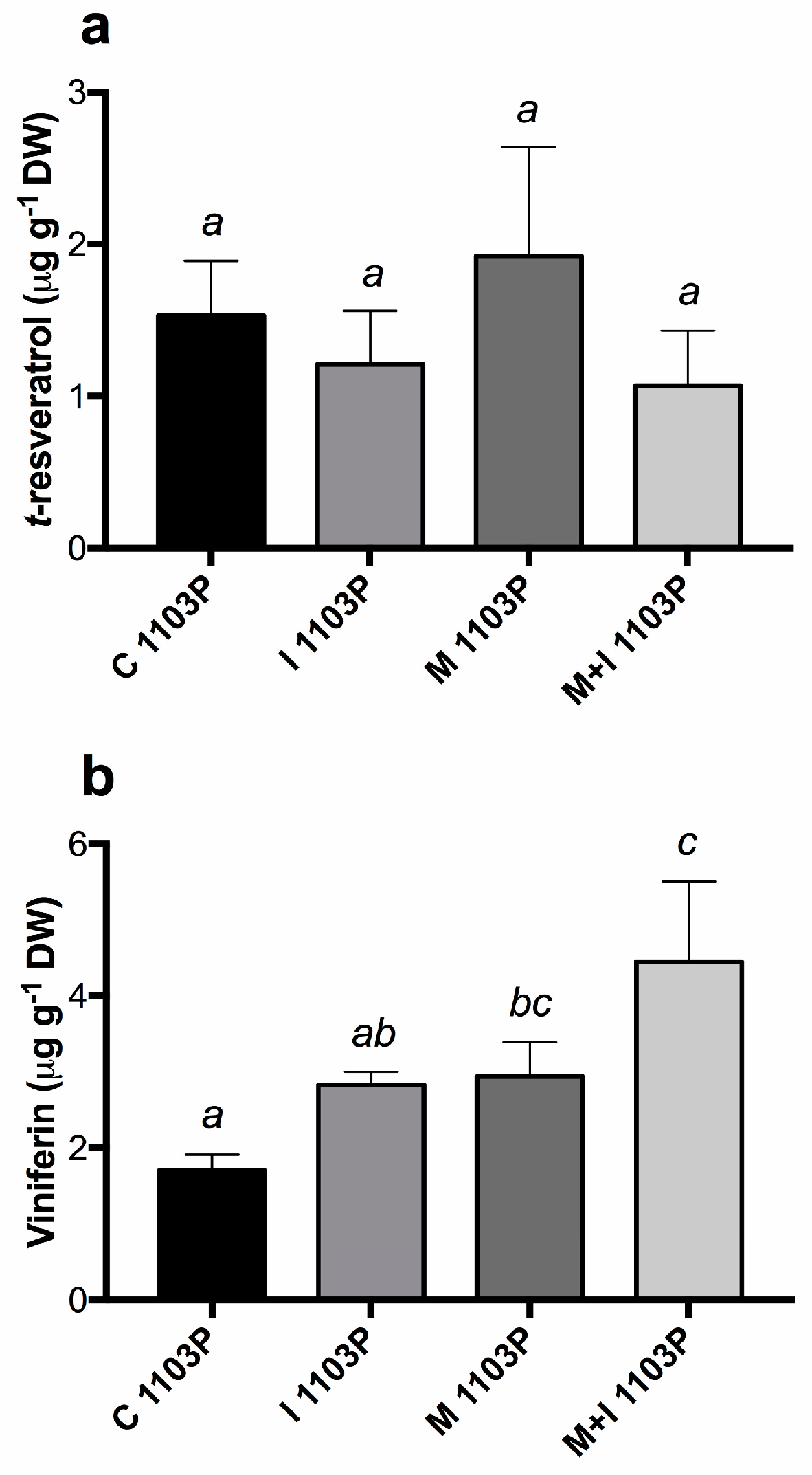
3.3. Growth—And Defence—Related Metabolites Content in Leaves
4. Discussion
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alagna, F.; Balestrini, R.; Chitarra, W.; Marsico, A.; Nerva, L. Getting Ready with the Priming: Innovative Weapons against Biotic and Abiotic Crop Enemies in a Global Changing Scenario. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Elsevier: Amsterdam, The Netherlands, 2020; pp. 35–56. [Google Scholar]
- Nerva, L.; Sandrini, M.; Moffa, L.; Velasco, R.; Balestrini, R.; Chitarra, W. Breeding toward Improved Ecological Plant–Microbiome Interactions. Trends Plant Sci. 2022, 27, 1134–1143. [Google Scholar] [CrossRef] [PubMed]
- Giudice, G.; Moffa, L.; Niero, M.; Duso, C.; Sandrini, M.; Vazzoler, L.F.; Luison, M.; Pasini, E.; Chitarra, W.; Nerva, L. Novel Sustainable Strategies to Control Plasmopara viticola in Grapevine Unveil New Insights on Priming Responses and Arthropods Ecology. Pest Manag. Sci. 2022, 78, 2342–2356. [Google Scholar] [CrossRef] [PubMed]
- Balestrini, R.; Chitarra, W.; Antoniou, C.; Ruocco, M.; Fotopoulos, V. Improvement of Plant Performance under Water Deficit with the Employment of Biological and Chemical Priming Agents. J. Agric. Sci. 2018, 156, 680–688. [Google Scholar] [CrossRef]
- Balestrini, R.; Brunetti, C.; Chitarra, W.; Nerva, L. Photosynthetic Traits and Nitrogen Uptake in Crops: Which Is the Role of Arbuscular Mycorrhizal Fungi? Plants 2020, 9, 1105. [Google Scholar] [CrossRef]
- Irankhah, S.; Sillo, F.; Nerva, L.; Ganjeali, A.; Balestrini, R.; Chitarra, W. Combined Effects of Water Deficit, Exogenous Ethylene Application and Root Symbioses on Trigonelline and ABA Accumulation in Fenugreek. Appl. Sci. 2020, 10, 2338. [Google Scholar] [CrossRef] [Green Version]
- Balestrini, R.; Nerva, L.; Sillo, F.; Girlanda, M.; Perotto, S. Plant and Fungal Gene Expression in Mycorrhizal Protocorms of the Orchid Serapias vomeracea Colonized by Tulasnella calospora. Plant Signal. Behav. 2014, 9, e977707. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.A.; van der Lelie, D.; Zarraonaindia, I. Microbial Terroir for Wine Grapes. Proc. Natl. Acad. Sci. USA 2014, 111, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Nerva, L.; Moffa, L.; Giudice, G.; Giorgianni, A.; Tomasi, D.; Chitarra, W. Microscale Analysis of Soil Characteristics and Microbiomes Reveals Potential Impacts on Plants and Fruit: Vineyard as a Model Case Study. Plant Soil 2021, 462, 525–541. [Google Scholar] [CrossRef]
- Sandrini, M.; Nerva, L.; Sillo, F.; Balestrini, R.; Chitarra, W.; Zampieri, E. Abiotic Stress and Belowground Microbiome: The Potential of Omics Approaches. Int. J. Mol. Sci. 2022, 23, 1091. [Google Scholar] [CrossRef]
- Schüβler, A.; Schwarzott, D.; Walker, C. A New Fungal Phylum, the Glomeromycota: Phylogeny and Evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef]
- Nerva, L.; Giudice, G.; Quiroga, G.; Belfiore, N.; Lovat, L.; Perria, R.; Volpe, M.G.; Moffa, L.; Sandrini, M.; Gaiotti, F. Mycorrhizal Symbiosis Balances Rootstock-Mediated Growth-Defence Tradeoffs. Biol. Fertil. Soils 2021, 58, 17–34. [Google Scholar] [CrossRef]
- Walder, F.; Niemann, H.; Natarajan, M.; Lehmann, M.F.; Boller, T.; Wiemken, A. Mycorrhizal Networks: Common Goods of Plants Shared under Unequal Terms of Trade. Plant Physiol. 2012, 159, 789–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balestrini, R.; Lumini, E. Focus on Mycorrhizal Symbioses. Appl. Soil Ecol. 2018, 123, 299–304. [Google Scholar] [CrossRef]
- Volpe, V.; Chitarra, W.; Cascone, P.; Volpe, M.G.; Bartolini, P.; Moneti, G.; Pieraccini, G.; Di Serio, C.; Maserti, B.; Guerrieri, E. The Association with Two Different Arbuscular Mycorrhizal Fungi Differently Affects Water Stress Tolerance in Tomato. Front. Plant Sci. 2018, 9, 1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balestrini, R.; Magurno, F.; Walker, C.; Lumini, E.; Bianciotto, V. Cohorts of Arbuscular Mycorrhizal Fungi (AMF) in Vitis vinifera, a Typical Mediterranean Fruit Crop. Environ. Microbiol. Rep. 2010, 2, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Trouvelot, S.; Bonneau, L.; Redecker, D.; Van Tuinen, D.; Adrian, M.; Wipf, D. Arbuscular Mycorrhiza Symbiosis in Viticulture: A Review. Agron. Sustain. Dev. 2015, 35, 1449–1467. [Google Scholar] [CrossRef] [Green Version]
- Valat, L.; Deglène-Benbrahim, L.; Kendel, M.; Hussenet, R.; Le Jeune, C.; Schellenbaum, P.; Maillot, P. Transcriptional Induction of Two Phosphate Transporter 1 Genes and Enhanced Root Branching in Grape Plants Inoculated with Funneliformis mosseae. Mycorrhiza 2018, 28, 179–185. [Google Scholar] [CrossRef]
- Nogales, A.; Ribeiro, H.; Nogales-Bueno, J.; Hansen, L.D.; Gonçalves, E.F.; Coito, J.L.; Rato, A.E.; Peixe, A.; Viegas, W.; Cardoso, H. Response of Mycorrhizal ‘Touriga Nacional’ Variety Grapevines to High Temperatures Measured by Calorespirometry and Near-Infrared Spectroscopy. Plants 2020, 9, 1499. [Google Scholar] [CrossRef]
- Cameron, D.D.; Neal, A.L.; van Wees, S.C.; Ton, J. Mycorrhiza-Induced Resistance: More than the Sum of Its Parts? Trends Plant Sci. 2013, 18, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Miozzi, L.; Vaira, A.M.; Brilli, F.; Casarin, V.; Berti, M.; Ferrandino, A.; Nerva, L.; Accotto, G.P.; Lanfranco, L. Arbuscular Mycorrhizal Symbiosis Primes Tolerance to Cucumber mosaic virus in Tomato. Viruses 2020, 12, 675. [Google Scholar] [CrossRef]
- Bruisson, S.; Maillot, P.; Schellenbaum, P.; Walter, B.; Gindro, K.; Deglène-Benbrahim, L. Arbuscular Mycorrhizal Symbiosis Stimulates Key Genes of the Phenylpropanoid Biosynthesis and Stilbenoid Production in Grapevine Leaves in Response to Downy Mildew and Grey Mould Infection. Phytochemistry 2016, 131, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Silva, A.; Figueiredo, A.; Sebastiana, M. First Insights into the Effect of Mycorrhizae on the Expression of Pathogen Effectors during the Infection of Grapevine with Plasmopara viticola. Sustainability 2021, 13, 1226. [Google Scholar] [CrossRef]
- Goddard, M.-L.; Belval, L.; Martin, I.R.; Roth, L.; Laloue, H.; Deglène-Benbrahim, L.; Valat, L.; Bertsch, C.; Chong, J. Arbuscular Mycorrhizal Symbiosis Triggers Major Changes in Primary Metabolism Together with Modification of Defense Responses and Signaling in Both Roots and Leaves of Vitis vinifera. Front. Plant Sci. 2021, 12, 1675. [Google Scholar] [CrossRef] [PubMed]
- Nogales, A.; Camprubí, A.; Estaún, V.; Marfà, V.; Calvet, C. In Vitro Interaction Studies between Glomus intraradices and Armillaria mellea in Vines. Span. J. Agric. Res. 2010, 8, 62–68. [Google Scholar] [CrossRef]
- Petit, E.; Gubler, W.D. Influence of Glomus intraradices on Black Foot Disease Caused by Cylindrocarpon macrodidymum on Vitis rupestris under Controlled Conditions. Plant Dis. 2006, 90, 1481–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.-Y.; Yang, G.-D.; Shu, H.-R.; Yang, Y.-T.; Ye, B.-X.; Nishida, I.; Zheng, C.-C. Colonization by the Arbuscular Mycorrhizal Fungus Glomus versiforme Induces a Defense Response against the Root-Knot Nematode Meloidogyne Incognita in the Grapevine (Vitis amurensis Rupr.), Which Includes Transcriptional Activation of the Class III Chitinase Gene VCH3. Plant Cell Physiol. 2006, 47, 154–163. [Google Scholar] [PubMed]
- Hao, Z.; Fayolle, L.; van Tuinen, D.; Chatagnier, O.; Li, X.; Gianinazzi, S.; Gianinazzi-Pearson, V. Local and Systemic Mycorrhiza-Induced Protection against the Ectoparasitic Nematode Xiphinema index Involves Priming of Defence Gene Responses in Grapevine. J. Exp. Bot. 2012, 63, 3657–3672. [Google Scholar] [CrossRef]
- Martignoni, M.M.; Garnier, J.; Hart, M.M.; Tyson, R.C. Investigating the Impact of the Mycorrhizal Inoculum on the Resident Fungal Community and on Plant Growth. Ecol. Model. 2020, 438, 109321. [Google Scholar] [CrossRef]
- Bastías, D.A.; Balestrini, R.; Pollmann, S.; Gundel, P.E. Environmental Interference of Plant-microbe Interactions. Plant Cell Environ. 2022, 45, 3387–3398. [Google Scholar] [CrossRef]
- Lovisolo, C.; Lavoie-Lamoureux, A.; Tramontini, S.; Ferrandino, A. Grapevine Adaptations to Water Stress: New Perspectives about Soil/Plant Interactions. Theor. Exp. Plant Physiol. 2016, 28, 53–66. [Google Scholar] [CrossRef]
- Warschefsky, E.J.; Klein, L.L.; Frank, M.H.; Chitwood, D.H.; Londo, J.P.; von Wettberg, E.J.; Miller, A.J. Rootstocks: Diversity, Domestication, and Impacts on Shoot Phenotypes. Trends Plant Sci. 2016, 21, 418–437. [Google Scholar] [CrossRef] [PubMed]
- Chitarra, W.; Perrone, I.; Avanzato, C.G.; Minio, A.; Boccacci, P.; Santini, D.; Gilardi, G.; Siciliano, I.; Gullino, M.L.; Delledonne, M. Grapevine Grafting: Scion Transcript Profiling and Defense-Related Metabolites Induced by Rootstocks. Front. Plant Sci. 2017, 8, 654. [Google Scholar] [CrossRef] [PubMed]
- Marasco, R.; Rolli, E.; Fusi, M.; Michoud, G.; Daffonchio, D. Grapevine Rootstocks Shape Underground Bacterial Microbiome and Networking but Not Potential Functionality. Microbiome 2018, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedini, A.; Mercy, L.; Schneider, C.; Franken, P.; Lucic-Mercy, E. Unraveling the Initial Plant Hormone Signaling, Metabolic Mechanisms and Plant Defense Triggering the Endomycorrhizal Symbiosis Behavior. Front. Plant Sci. 2018, 9, 1800. [Google Scholar] [CrossRef] [Green Version]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in Action: Signalling, Transport and the Control of Plant Growth and Development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Paque, S.; Weijers, D. Q&A: Auxin: The Plant Molecule That Influences Almost Anything. BMC Biol. 2016, 14, 67. [Google Scholar]
- Wasilewska, A.; Vlad, F.; Sirichandra, C.; Redko, Y.; Jammes, F.; Valon, C.; dit Frey, N.F.; Leung, J. An Update on Abscisic Acid Signaling in Plants and More…. Mol. Plant 2008, 1, 198–217. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Li, G.; Bressan, R.A.; Song, C.; Zhu, J.; Zhao, Y. Abscisic Acid Dynamics, Signaling, and Functions in Plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [Green Version]
- Schnee, S.; Viret, O.; Gindro, K. Role of Stilbenes in the Resistance of Grapevine to Powdery Mildew. Physiol. Mol. Plant Pathol. 2008, 72, 128–133. [Google Scholar] [CrossRef]
- Viret, O.; Spring, J.-L.; Gindro, K. Stilbenes: Biomarkers of Grapevine Resistance to Fungal Diseases. Oeno One 2018, 52, 235–241. [Google Scholar]
- Zanin, L.; Nerva, L.; Alessandrini, M.; Tomasi, D.; Pinton, R.; Lucchetta, M.; Chitarra, W.; Gaiotti, F. Effects of the Fractionation of the Nitrogen Fertilization on Root Nitrate Uptake and Vine Agronomic Performance in Pinot Gris Grapevines in a Temperate Climate. J. Soil Sci. Plant Nutr. 2022, 22, 4996–5008. [Google Scholar] [CrossRef]
- Balestrini, R.; Salvioli, A.; Dal Molin, A.; Novero, M.; Gabelli, G.; Paparelli, E.; Marroni, F.; Bonfante, P. Impact of an Arbuscular Mycorrhizal Fungus versus a Mixed Microbial Inoculum on the Transcriptome Reprogramming of Grapevine Roots. Mycorrhiza 2017, 27, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Nerva, L.; Guaschino, M.; Pagliarani, C.; De Rosso, M.; Lovisolo, C.; Chitarra, W. Spray-induced Gene Silencing Targeting a Glutathione S-transferase Gene Improves Resilience to Drought in Grapevine. Plant Cell Environ. 2022, 45, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Belfiore, N.; Nerva, L.; Fasolini, R.; Gaiotti, F.; Lovat, L.; Chitarra, W. Leaf Gas Exchange and Abscisic Acid in Leaves of Glera Grape Variety during Drought and Recovery. Theor. Exp. Plant Physiol. 2021, 33, 261–270. [Google Scholar] [CrossRef]
- Belfiore, N.; Vinti, R.; Lovat, L.; Chitarra, W.; Tomasi, D.; de Bei, R.; Meggio, F.; Gaiotti, F. Infrared Thermography to Estimate Vine Water Status: Optimizing Canopy Measurements and Thermal Indices for the Varieties Merlot and Moscato in Northern Italy. Agronomy 2019, 9, 821. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Gulías, J.; Jonasson, S.; Medrano, H.; Mus, M. Seasonal Patterns and Control of Gas Exchange in Local Populations of the Mediterranean Evergreen Shrub Pistacia lentiscus L. Acta Oecologica 2001, 22, 33–43. [Google Scholar] [CrossRef]
- He, Z.; Webster, S.; He, S.Y. Growth–Defense Trade-Offs in Plants. Curr. Biol. 2022, 32, R634–R639. [Google Scholar] [CrossRef]
- Sandrini, M.; Moffa, L.; Velasco, R.; Balestrini, R.; Chitarra, W.; Nerva, L. Microbe-Assisted Crop Improvement: A Sustainable Weapon to Restore Holobiont Functionality and Resilience. Hortic. Res. 2022, 9. [Google Scholar] [CrossRef]
- Jacott, C.N.; Murray, J.D.; Ridout, C.J. Trade-Offs in Arbuscular Mycorrhizal Symbiosis: Disease Resistance, Growth Responses and Perspectives for Crop Breeding. Agronomy 2017, 7, 75. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Pieterse, C.M.; Bakker, P.A.; Berendsen, R.L. Beneficial Microbes Going Underground of Root Immunity. Plant Cell Environ. 2019, 42, 2860–2870. [Google Scholar] [CrossRef] [Green Version]
- Barea, J.-M.; Azcón, R.; Azcón-Aguilar, C. Mycorrhizosphere Interactions to Improve Plant Fitness and Soil Quality. Antonie Van Leeuwenhoek 2002, 81, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Bücking, H.; Kafle, A. Role of Arbuscular Mycorrhizal Fungi in the Nitrogen Uptake of Plants: Current Knowledge and Research Gaps. Agronomy 2015, 5, 587–612. [Google Scholar] [CrossRef]
- Guether, M.; Balestrini, R.; Hannah, M.; He, J.; Udvardi, M.K.; Bonfante, P. Genome-wide Reprogramming of Regulatory Networks, Transport, Cell Wall and Membrane Biogenesis during Arbuscular Mycorrhizal Symbiosis in Lotus japonicus. New Phytol. 2009, 182, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Augé, R.M.; Toler, H.D.; Saxton, A.M. Mycorrhizal Stimulation of Leaf Gas Exchange in Relation to Root Colonization, Shoot Size, Leaf Phosphorus and Nitrogen: A Quantitative Analysis of the Literature Using Meta-Regression. Front. Plant Sci. 2016, 7, 1084. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd_Allah, E.F.; Hashem, A. Phytohormones and Beneficial Microbes: Essential Components for Plants to Balance Stress and Fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef]
- Chaves, M.; Flexas, J.; Pinheiro, C. Photosynthesis under Drought and Salt Stress: Regulation Mechanisms from Whole Plant to Cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Chitarra, W.; Pagliarani, C.; Maserti, B.; Lumini, E.; Siciliano, I.; Cascone, P.; Schubert, A.; Gambino, G.; Balestrini, R.; Guerrieri, E. Insights on the Impact of Arbuscular Mycorrhizal Symbiosis on Tomato Tolerance to Water Stress. Plant Physiol. 2016, 171, 1009–1023. [Google Scholar] [CrossRef] [Green Version]
- Barros, V.; Frosi, G.; Santos, M.; Ramos, D.G.; Falcão, H.M.; Santos, M.G. Arbuscular Mycorrhizal Fungi Improve Photosynthetic Energy Use Efficiency and Decrease Foliar Construction Cost under Recurrent Water Deficit in Woody Evergreen Species. Plant Physiol. Biochem. 2018, 127, 469–477. [Google Scholar] [CrossRef]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Regulation of Root Water Uptake under Abiotic Stress Conditions. J. Exp. Bot. 2012, 63, 43–57. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Ding, L.; Chaumont, F.; Aroca, R.; Ruiz-Lozano, J.M. The Arbuscular Mycorrhizal Symbiosis Regulates Aquaporins Activity and Improves Root Cell Water Permeability in Maize Plants Subjected to Water Stress. Plant Cell Environ. 2019, 42, 2274–2290. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, W.; Chang, S.; Anyia, A. Phosphorus Fertilization and Fungal Inoculations Affected the Physiology, Phosphorus Uptake and Growth of Spring Wheat under Rainfed Conditions on the Canadian Prairies. J. Agron. Crop Sci. 2013, 199, 85–93. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of Arbuscular Mycorrhizae on Photosynthesis and Water Status of Maize Plants under Salt Stress. Mycorrhiza 2008, 18, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O. The Power of Auxin in Plants. Plant Physiol. 2010, 154, 501–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig-Müller, J.; Güther, M. Auxins as Signals in Arbuscular Mycorrhiza Formation. Plant Signal. Behav. 2007, 2, 194–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vos, C.; Schouteden, N.; van Tuinen, D.; Chatagnier, O.; Elsen, A.; De Waele, D.; Panis, B.; Gianinazzi-Pearson, V. Mycorrhiza-Induced Resistance against the Root–Knot Nematode Meloidogyne incognita Involves Priming of Defense Gene Responses in Tomato. Soil Biol. Biochem. 2013, 60, 45–54. [Google Scholar] [CrossRef]
- Campos-Soriano, L.; García-Martínez, J.; Segundo, B.S. The Arbuscular Mycorrhizal Symbiosis Promotes the Systemic Induction of Regulatory Defence-related Genes in Rice Leaves and Confers Resistance to Pathogen Infection. Mol. Plant Pathol. 2012, 13, 579–592. [Google Scholar] [CrossRef]
- Bettenfeld, P.; i Canals, J.C.; Jacquens, L.; Fernandez, O.; Fontaine, F.; van Schaik, E.; Courty, P.-E.; Trouvelot, S. The Microbiota of the Grapevine Holobiont: A Key Component of Plant Health. J. Adv. Res. 2021, 40, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Carlström, C.I.; Field, C.M.; Bortfeld-Miller, M.; Müller, B.; Sunagawa, S.; Vorholt, J.A. Synthetic Microbiota Reveal Priority Effects and Keystone Strains in the Arabidopsis Phyllosphere. Nat. Ecol. Evol. 2019, 3, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, C.; Loverock, L.; Kokkoris, V.; Holland, T.; Bowen, P.A.; Hart, M. Commercial Arbuscular Mycorrhizal Fungal Inoculant Failed to Establish in a Vineyard despite Priority Advantage. PeerJ 2021, 9, e11119. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nerva, L.; Balestrini, R.; Chitarra, W. From Plant Nursery to Field: Persistence of Mycorrhizal Symbiosis Balancing Effects on Growth-Defence Tradeoffs Mediated by Rootstock. Agronomy 2023, 13, 229. https://doi.org/10.3390/agronomy13010229
Nerva L, Balestrini R, Chitarra W. From Plant Nursery to Field: Persistence of Mycorrhizal Symbiosis Balancing Effects on Growth-Defence Tradeoffs Mediated by Rootstock. Agronomy. 2023; 13(1):229. https://doi.org/10.3390/agronomy13010229
Chicago/Turabian StyleNerva, Luca, Raffaella Balestrini, and Walter Chitarra. 2023. "From Plant Nursery to Field: Persistence of Mycorrhizal Symbiosis Balancing Effects on Growth-Defence Tradeoffs Mediated by Rootstock" Agronomy 13, no. 1: 229. https://doi.org/10.3390/agronomy13010229