
Rhizosphere Microbial Community Shows a Greater Response Than Soil Properties to Tea (Camellia sinensis L.) Cultivars
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Soil Sampling
2.4. Soil Chemical Property Analysis, and Soil Fertility Index Calculation
2.5. DNA Extraction, PCR Amplification, and Amplicon Sequencing
2.6. High-Throughput Sequencing Data Processing
2.7. Statistical Analysis
3. Results
3.1. Changes in Soil Properties and Fertility Index
3.2. Rhizosphere Microbial Richness and Diversity
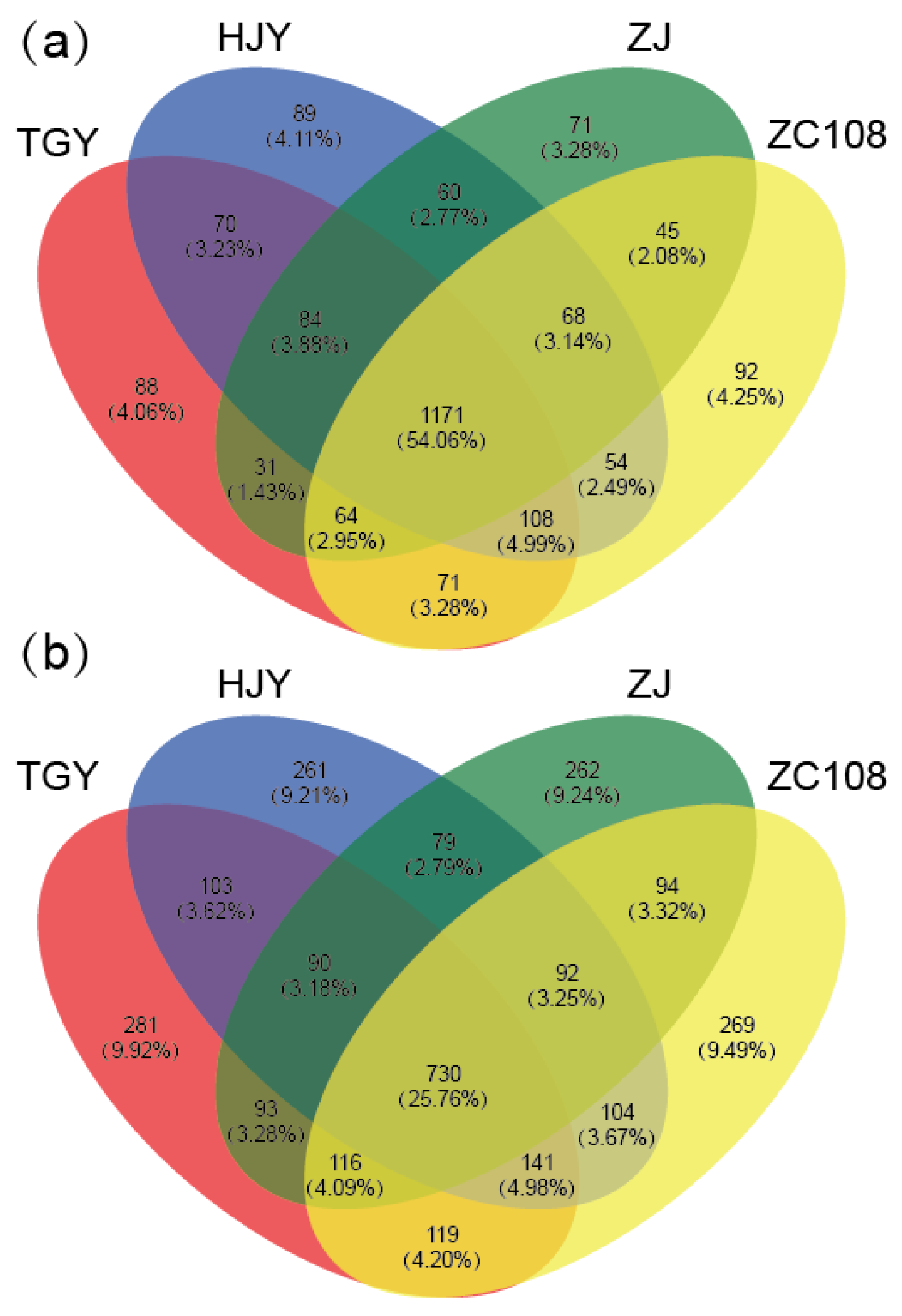
3.3. Rhizosphere Microbial Community Composition
3.4. Microbial Community of Soils with Different Tea Cultivars
3.5. Correlation between Microbial Communities and Soil Properties
3.6. Potential Functions of Microbial Community
4. Discussion
4.1. Changes in Soil Properties among Different Tea Cultivars
4.2. Culitvar-Associated Response Characteristics of Soil Microbial Diversity, Community Composition, and Their Potential Functions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deng, L.J.; Zeng, G.M.; Fan, C.Z.; Lu, L.H.; Chen, X.F.; Chen, M.; Wu, H.P.; He, X.X.; He, Y. Response of rhizosphere microbial community structure and diversity to heavy metal co-pollution in arable soil. Microbiol. Biotechnol. 2015, 99, 8259–8269. [Google Scholar] [CrossRef]
- Frenk, S.; Hadar, Y.; Minz, D. Resilience of soil bacterial community to irrigation with water of different qualities under Mediterranean climate. Environ. Microbiol. 2014, 16, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.D.; Ma, L.F.; Ji, L.F.; Shi, Y.Z.; Yi, X.Y.; Yang, Q.L.; Ni, K.; Ruan, J.Y. Long-term nitrogen fertilization indirectly affects soil fungi community structure by changing soil and pruned litter in a subtropical tea (Camellia sinensis L.) plantation in China. Plant Soil 2019, 444, 409–426. [Google Scholar] [CrossRef]
- Wu, H.M.; Wu, H.M.; Qin, X.J.; Lin, M.H.; Zhao, Y.L.; Rensing, C.; Lin, W.X. Replanting disease alters the faunal community composition and diversity in the rhizosphere soil of Radix pseudostellariae. Agric. Ecosyst. Environ. 2021, 310, 107304. [Google Scholar] [CrossRef]
- Zhao, M.L.; Wang, M.; Zhao, Y.T.; Wang, G.D.; Xue, Z.S.; Jiang, M. Variations in soil microbial communities in the sedge-dominated peatlands along an altitude gradient on the northern slope of Changbai Mountain, China. Ecol. Indic. 2021, 129, 107964. [Google Scholar] [CrossRef]
- Marschner, P.; Crowley, D.; Yang, C.H. Development of specific rhizosphere bacterial communities in relation to plant species, nutrition and soil type. Plant Soil 2004, 261, 199–208. [Google Scholar] [CrossRef]
- Shen, C.C.; Xiong, J.B.; Zhang, H.Y.; Feng, Y.Z.; Lin, X.G.; Li, X.Y.; Liang, W.J.; Chu, H.Y. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Kowalchuk, G.A.; Stephen, J.R.; DeBoer, W.; Prosser, J.I.; Embley, T.M.; Woldendorp, J.W. Analysis of ammonia-oxidizing bacteria of the beta subdivision of the class Proteobacteria in coastal sand dunes by denaturing gradient gel electrophoresis and sequencing of PCR-amplified 16S ribosomal DNA fragments. Appl. Environ. Microbiol. 1997, 63, 1489–1497. [Google Scholar] [CrossRef] [Green Version]
- Rousk, J.; Brookes, P.C.; Baath, E. Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Environ. Microbiol. 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Barcenas-Moreno, G.; Baath, E.; Rousk, J. Functional implications of the pH-trait distribution of the microbial community in a re-inoculation experiment across a pH gradient. Soil Biol. Biochem. 2016, 93, 69–78. [Google Scholar] [CrossRef]
- Lorenz, M.; Hofmann, D.; Steffen, B.; Fischer, K.; Thiele-Bruhn, S. The molecular composition of extractable soil microbial compounds and their contribution to soil organic matter vary with soil depth and tree species. Sci. Total Environ. 2021, 781, 146732. [Google Scholar] [CrossRef]
- Li, Y.B.; Bezemer, T.M.; Yang, J.J.; Lu, X.T.; Li, X.Y.; Liang, W.J.; Han, X.G.; Li, Q. Changes in litter quality induced by N deposition alter soil microbial communities. Soil Biol. Biochem. 2019, 130, 33–42. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, W.; Shen, Z.J.; Wang, J.J.; Chen, Y.J.; Wang, D.; Liu, G.; Han, M.Z. Comparison and interpretation of characteristics of Rhizosphere microbiomes of three blueberry varieties. BMC Microbiol. 2021, 21, 30. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, L.M.; Hattenschwiler, S.; Milcu, A.; Wambsganss, J.; Shihan, A.; Fromin, N. Tree species mixing affects soil microbial functioning indirectly via root and litter traits and soil parameters in European forests. Funct. Ecol. 2021, 35, 2190–2204. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Tkacz, A.; Cheema, J.; Chandra, G.; Grant, A.; Poole, P.S. Stability and succession of the rhizosphere microbiota depends upon plant type and soil composition. ISME J. 2015, 9, 2349–2359. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.; Haddix, M.L.; Byrne, P.F.; Cotrufo, M.F.; Schipanski, M.E.; Kallenbach, C.M.; Wallenstein, M.D.; Fonte, S.J. Long-term compost amendment modulates wheat genotype differences in belowground carbon allocation, microbial rhizosphere recruitment and nitrogen acquisition. Soil Biol. Biochem. 2022, 172, 108768. [Google Scholar] [CrossRef]
- Tang, D.D.; Liu, M.Y.; Zhang, Q.F.; Ma, L.F.; Shi, Y.Z.; Ruan, J.Y. Preferential assimilation of NH4+ over NO3− in tea plant associated with genes involved in nitrogen transportation, utilization and catechins biosynthesis. Plant Sci. 2020, 291, 110369. [Google Scholar] [CrossRef]
- Marschner, H.; Romheld, V.; Cakmak, I. Root-induced changes of nutrient availability in the rhizosphere. J. Plant Nutr. 1987, 10, 1175–1184. [Google Scholar] [CrossRef]
- Yang, X.D.; Ni, K.; Shi, Y.Z.; Yi, X.Y.; Zhang, Q.F.; Fang, L.; Ma, L.F.; Ruan, J.Y. Effects of long-term nitrogen application on soil acidification and solution chemistry of a tea plantation in China. Agric. Ecosyst. Environ. 2018, 252, 74–82. [Google Scholar] [CrossRef]
- Li, Y.C.; Li, Z.; Li, Z.W.; Jiang, Y.H.; Weng, B.Q.; Lin, W.X. Variations of rhizosphere bacterial communities in tea (Camellia sinensis L.) continuous cropping soil by high-throughput pyrosequencing approach. J. Appl. Microbiol. 2016, 121, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Zheng, Z.C.; Li, T.X.; Wang, Y.D.; Huang, H.G.; Yu, H.Y.; Ye, D.H.; Liu, T.; Yao, T.Y.; Zhang, X.Z. Variations of fungal communities within the soils of different tea varieties (Camellia sinensis L.) following long-term plantation. Plant Soil 2022, 477, 665–677. [Google Scholar] [CrossRef]
- Badri, D.V.; Weir, T.L.; van der Lelie, D.; Vivanco, J.M. Rhizosphere chemical dialogues: Plant-microbe interactions. Curr. Opin. Biotechnol. 2009, 20, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Lareen, A.; Burton, F.; Schafer, P. Plant root-microbe communication in shaping root microbiomes. Plant Mol. Biol. 2016, 90, 575–587. [Google Scholar] [CrossRef] [Green Version]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Shang, Q.Y.; Ling, N.; Feng, X.M.; Yang, X.X.; Wu, P.P.; Zou, J.W.; Shen, Q.R.; Guo, S.W. Soil fertility and its significance to crop productivity and sustainability in typical agroecosystem: A summary of long-term fertilizer experiments in China. Plant Soil 2014, 381, 13–23. [Google Scholar] [CrossRef]
- Shukla, M.K.; Lal, R.; Ebinger, M. Determining soil quality indicators by factor analysis. Soil Tillage Res. 2006, 87, 194–204. [Google Scholar] [CrossRef]
- Doran, J.W.; Parkin, T.B. Defining and assessing soil quality. In Defining soil quality for a sustainable environment; Doran, J.W., Coleman, D.C., Bezdick, D.F., Stewart, B.A., Eds.; Soil Science Society of America, Inc.: Madison, WI, USA, 1994; Volume 35, pp. 3–21. [Google Scholar]
- Beckers, B.; De Beeck, M.O.; Thijs, S.; Truyens, S.; Weyens, N.; Boerjan, W.; Vangronsveld, J. Performance of 16s rDNA Primer Pairs in the Study of Rhizosphere and Endosphere Bacterial Microbiomes in Metabarcoding Studies. Front. Microbiol. 2016, 7, 650. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Yu, G.; Shi, C.P.; Liu, L.M.; Guo, Q.; Han, C.; Zhang, D.; Zhang, L.; Liu, B.X.; Gao, H.; et al. Majorbio Cloud: A one-stop, comprehensive bioinformatic platform for multiomics analyses. Anal. iMeta 2022, 1, e12. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Reyon, D.; Tsai, S.Q.; Khayter, C.; Foden, J.A.; Sander, J.D.; Joung, J.K. FLASH assembly of TALENs for high-throughput genome editing. Nat. Biotechnol. 2012, 30, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Gloeckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.J.; Li, S.Z.; Li, R.P.; Zhang, J.; Liu, Y.H.; Lv, L.F.; Zhu, H.; Wu, W.L.; Li, W.L. Plant cultivars imprint the rhizosphere bacterial community composition and association networks. Soil Biol. Biochem. 2017, 109, 145–155. [Google Scholar] [CrossRef]
- Kong, X.; Han, Z.F.; Tai, X.; Jin, D.C.; Ai, S.; Zheng, X.X.; Bai, Z.H. Maize (Zea mays L. Sp.) varieties significantly influence bacterial and fungal community in bulk soil, rhizosphere soil and phyllosphere. FEMS Microbiol. Ecol. 2020, 96, fiaa020. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Yoshihashi, T.; Worthington, M.; Nakahara, K.; Ando, Y.; Sahrawat, K.L.; Rao, I.M.; Lata, J.C.; Kishii, M.; Braun, H.J. Suppression of soil nitrification by plants. Plant Sci. 2015, 233, 155–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vengavasi, K.; Pandey, R. Root exudation potential in contrasting soybean genotypes in response to low soil phosphorus availability is determined by photo-biochemical processes. Plant Physiol. Biochem. 2018, 124, 1–9. [Google Scholar] [CrossRef]
- Ghatak, A.; Schindler, F.; Bachmann, G.; Engelmeier, D.; Bajaj, P.; Brenner, M.; Fragner, L.; Varshney, R.K.; Subbarao, G.V.; Chaturvedi, P. Root exudation of contrasting drought-stressed pearl millet genotypes conveys varying biological nitrification inhibition (BNI) activity. Biol. Fertil. Soils 2022, 58, 291–306. [Google Scholar] [CrossRef]
- Kan, Z.R.; Liu, Q.Y.; Virk, A.L.; He, C.; Qi, J.Y.; Dang, Y.P.; Zhao, X.; Zhang, H.L. Effects of experiment duration on carbon mineralization and accumulation under no-till. Soil Tillage Res. 2021, 209, 104939. [Google Scholar] [CrossRef]
- Odlare, M.; Pell, M.; Arthurson, J.V.; Abubaker, J.; Nehrenheim, E. Combined mineral N and organic waste fertilization—Effects on crop growth and soil properties. J. Agric. Sci. 2014, 152, 134–145. [Google Scholar] [CrossRef]
- Bao, L.J.; Sun, B.; Wei, Y.X.; Xu, N.; Zhang, S.W.; Gu, L.K.; Bai, Z.H. Grape cultivar features differentiate the grape rhizosphere microbiota. Plants 2022, 11, 1111. [Google Scholar] [CrossRef]
- Zheng, Y.Y.; Li, X.Z.; Cao, H.L.; Lei, L.; Zhang, X.; Han, D.J.; Wang, J.M.; Yao, M.J. The assembly of wheat-associated fungal community differs across growth stages. Appl. Microbiol. Biotechnol. 2021, 105, 7427–7438. [Google Scholar] [CrossRef]
- Ndour, P.M.S.; Barry, C.M.; Tine, D.; De la Fuente Canto, C.; Gueye, M.; Barakat, M.; Ortet, P.; Achouak, W.; Ndoye, I.; Sine, B.; et al. Pearl millet genotype impacts microbial diversity and enzymatic activities in relation to root-adhering soil aggregation. Plant Soil 2021, 464, 109–129. [Google Scholar] [CrossRef]
- Ai, C.; Zhang, S.Q.; Zhang, X.; Guo, D.D.; Zhou, W.; Huang, S.M. Distinct responses of soil bacterial and fungal communities to changes in fertilization regime and crop rotation. Geoderma 2018, 319, 156–166. [Google Scholar] [CrossRef]
- Demoling, F.; Figueroa, D.; Baath, E. Comparison of factors limiting bacterial growth in different soils. Soil Biol. Biochem. 2007, 39, 2485–2495. [Google Scholar] [CrossRef]
- Lazcano, C.; Gomez-Brandon, M.; Revilla, P.; Dominguez, J. Short-term effects of organic and inorganic fertilizers on soil microbial community structure and function. Biol. Fertil. Soils 2013, 49, 723–733. [Google Scholar] [CrossRef]
- Nunes, C.A.; Barlow, J.; Franca, F.; Berenguer, E.; Solar, R.R.C.; Louzada, J.; Leitao, R.P.; Maia, L.F.; Oliveira, V.H.F.; Braga, R.F.; et al. Functional redundancy of Amazonian dung beetles confers community-level resistance to primary forest disturbance. Biotropica 2021, 53, 1510–1521. [Google Scholar] [CrossRef]
- Xiong, C.; He, J.Z.; Singh, B.K.; Zhu, Y.G.; Wang, J.T.; Li, P.P.; Zhang, Q.B.; Han, L.L.; Shen, J.P.; Ge, A.H.; et al. Rare taxa maintain the stability of crop mycobiomes and ecosystem functions. Environ. Microbiol. 2021, 23, 1907–1924. [Google Scholar] [CrossRef]
- Li, F.; Chen, L.; Zhang, J.B.; Yin, J.; Huang, S.M. Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Front. Microbiol. 2017, 8, 187. [Google Scholar] [CrossRef] [Green Version]
- Soman, C.; Li, D.F.; Wander, M.M.; Kent, A.D. Long-term fertilizer and crop-rotation treatments differentially affect soil bacterial community structure. Plant Soil 2017, 413, 145–159. [Google Scholar] [CrossRef]
- Yang, X.D.; Leng, Y.; Zhou, Z.Y.; Shang, H.G.; Ni, K.; Ma, L.F.; Yi, X.Y.; Cai, Y.J.; Ji, L.F.; Ruan, J.Y.; et al. Ecological management model for the improvement of soil fertility through the regulation of rare microbial taxa in tea (Camellia sinensis L.) plantation soils. J. Environ. Manag. 2022, 308, 114595. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.B.; Dsouza, M.; Gilbert, J.A.; Guo, X.S.; Wang, D.Z.; Guo, Z.B.; Ni, Y.Y.; Chu, H.Y. Fungal community composition in soils subjected to long-term chemical fertilization is most influenced by the type of organic matter. Environ. Microbiol. 2016, 18, 5137–5150. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Brookes, P.C.; Baath, E. Fungal and bacterial growth responses to N fertilization and pH in the 150-year ‘Park Grass’ UK grassland experiment. FEMS Microbiol. Ecol. 2011, 76, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Beales, N. Adaptation of microorganisms to cold temperatures, weak acid preservatives, low pH, and osmotic stress: A review. Compr. Rev. Food. Sci. Food Saf. 2004, 3, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Glassman, S.I.; Wang, I.J.; Bruns, T.D. Environmental filtering by pH and soil nutrients drives community assembly in fungi at fine spatial scales. Mol. Ecol. 2017, 26, 6960–6973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, D.; Bardgett, R.D.; Finlay, R.D.; Jones, D.L.; Philippot, L. A plant perspective on nitrogen cycling in the rhizosphere. Funct. Ecol. 2019, 33, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Liu, Y.; Wang, L.Y.; Bai, P.X.; Ruan, L.; Zhang, C.C.; Wei, K.; Cheng, H. Molecular cloning and expression analysis of ammonium transporters in tea plants (Camellia sinensis (L.) O. Kuntze) under different nitrogen treatments. Gene 2018, 658, 136–145. [Google Scholar] [CrossRef]
- Pajares, S.; Campo, J.; Bohannan, B.J.M.; Etchevers, J.D. Environmental controls on soil microbial communities in a seasonally dry tropical forest. Appl. Environ. Microbiol. 2018, 84, e00342-18. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.M.; Tan, Y.S.; Chen, G.W.; Wang, G.; Sun, J.X.; Ou, S.Y.; Chen, W.; Bai, W.B. Metabolism of anthocyanins and consequent effects on the gut microbiota. Crit. Rev. Food Sci. Nutr. 2019, 59, 982–991. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Tea Cultivars | |||
|---|---|---|---|---|
| HJY | TGY | ZC108 | ZJ | |
| pH (H2O) | 3.95 (0.10) a | 3.94 (0.06) a | 3.85 (0.07) a | 3.81 (0.07) a |
| NH4+ (mg kg−1) | 1.30 (0.29) a | 1.71 (0.31) a | 1.46 (0.25) a | 1.76 (0.45) a |
| NO3- (mg kg−1) | 4.42 (0.26) a | 4.64 (0.65) a | 4.47 (0.34) a | 4.24 (0.21) a |
| AP (mg kg−1) | 1.85 (0.54) a | 1.57 (0.34) a | 0.87 (0.18) a | 1.23 (0.30) a |
| AK (mg kg−1) | 286 (19) a | 252 (22) ab | 217 (14) bc | 197 (13) c |
| TP (mg kg−1) | 424 (30) a | 471 (28) a | 399 (36) a | 452 (38) a |
| TK (mg kg−1) | 5527 (498) a | 4983 (680) a | 5020 (704) a | 5316 (469) a |
| SOC (g kg−1) | 9.1 (0.5) a | 9.3 (0.5) a | 9.4 (0.5) a | 8.5 (0.4) a |
| TN (g kg−1) | 1.2 (0.1) a | 1.2 (0.1) a | 1.2 (0.0) a | 1.1 (0.0) a |
| C/N | 7.60 (0.24) a | 7.83 (0.16) a | 7.82 (0.20) a | 7.48 (0.20) a |
| SFI | 0.55 (0.07) a | 0.58 (0.04) a | 0.52 (0.05) a | 0.55 (0.07) a |
| Parameters | Tea Cultivars | |||
|---|---|---|---|---|
| HJY | TGY | ZC108 | ZJ | |
| Bacteria | ||||
| Phylum | 21 (0.4) a | 21 (0.5) a | 20 (0.4) a | 20 (0.4) a |
| Class | 42 (0.8) a | 42 (1.3) a | 40 (1.5) a | 40 (1.8) a |
| Order | 101 (1.4) a | 101 (2.0) a | 99 (3.3) a | 96 (1.9) a |
| Family | 159 (1.6) a | 158 (3.5) a | 155 (3.7) ab | 149 (3.0) b |
| Genus | 271 (5.1) a | 266 (6.8) ab | 439 (5.0) ab | 422 (5.8) b |
| Species | 452 (8.8)a | 445 (10.6) ab | 439 (9.5) ab | 422 (9.0) b |
| Chao1 | 1267 (26) a | 1267 (21) a | 1253 (38) a | 1202 (12) a |
| Shannon | 5.11 (0.02) a | 4.96 (0.07) ab | 5.05 (0.06) ab | 4.92 (0.05) b |
| Fungi | ||||
| Phylum | 12 (0.2) a | 12 (0.2) a | 12 (0.2) a | 12 (0.2) a |
| Class | 33 (0.7) a | 34 (0.3) a | 33 (0.7) a | 32 (1.0) a |
| Order | 73 (2.1) a | 75 (1.7) a | 72 (2.4) a | 71 (2.9) a |
| Family | 147 (4.2) a | 151 (4.4) a | 141 (4.3) a | 143 (2.9) a |
| Genus | 221 (8.1) a | 226 (6.7) a | 216 (7.7) a | 210 (11.2) a |
| Species | 283 (9.9) a | 285 (6.0) a | 273 (8.9) a | 367 (14.0) a |
| Chao1 | 691 (35) a | 734 (30) a | 700 (30) a | 637 (35) a |
| Shannon | 3.71 (0.14) a | 3.63 (0.13) a | 3.64 (0.09) a | 3.86 (0.10) a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, S.; Liu, B.; Ni, K.; Ma, L.; Shi, Y.; Leng, Y.; Zheng, S.; Gao, S.; Yang, X.; Ruan, J. Rhizosphere Microbial Community Shows a Greater Response Than Soil Properties to Tea (Camellia sinensis L.) Cultivars. Agronomy 2023, 13, 221. https://doi.org/10.3390/agronomy13010221
Wei S, Liu B, Ni K, Ma L, Shi Y, Leng Y, Zheng S, Gao S, Yang X, Ruan J. Rhizosphere Microbial Community Shows a Greater Response Than Soil Properties to Tea (Camellia sinensis L.) Cultivars. Agronomy. 2023; 13(1):221. https://doi.org/10.3390/agronomy13010221
Chicago/Turabian StyleWei, Sirou, Boheng Liu, Kang Ni, Lifeng Ma, Yuanzhi Shi, Yang Leng, Shenghong Zheng, Shuilian Gao, Xiangde Yang, and Jianyun Ruan. 2023. "Rhizosphere Microbial Community Shows a Greater Response Than Soil Properties to Tea (Camellia sinensis L.) Cultivars" Agronomy 13, no. 1: 221. https://doi.org/10.3390/agronomy13010221