Effects of the Continuous Cropping and Soilborne Diseases of Panax Ginseng C. A. Meyer on Rhizosphere Soil Physicochemical Properties, Enzyme Activities, and Microbial Communities

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sample Collection
2.2. Soil Physicochemical Analysis
2.3. Soil Enzyme Activities
2.4. DNA Extraction and MiSeq Sequencing
2.5. Sequencing Data Analysis
2.6. Statistical Analysis
3. Results
3.1. Soil Physicochemical Properties
3.2. Soil Enzyme Activity
3.3. Correlations between Soil Physicochemical Properties and Soil Enzyme Activities
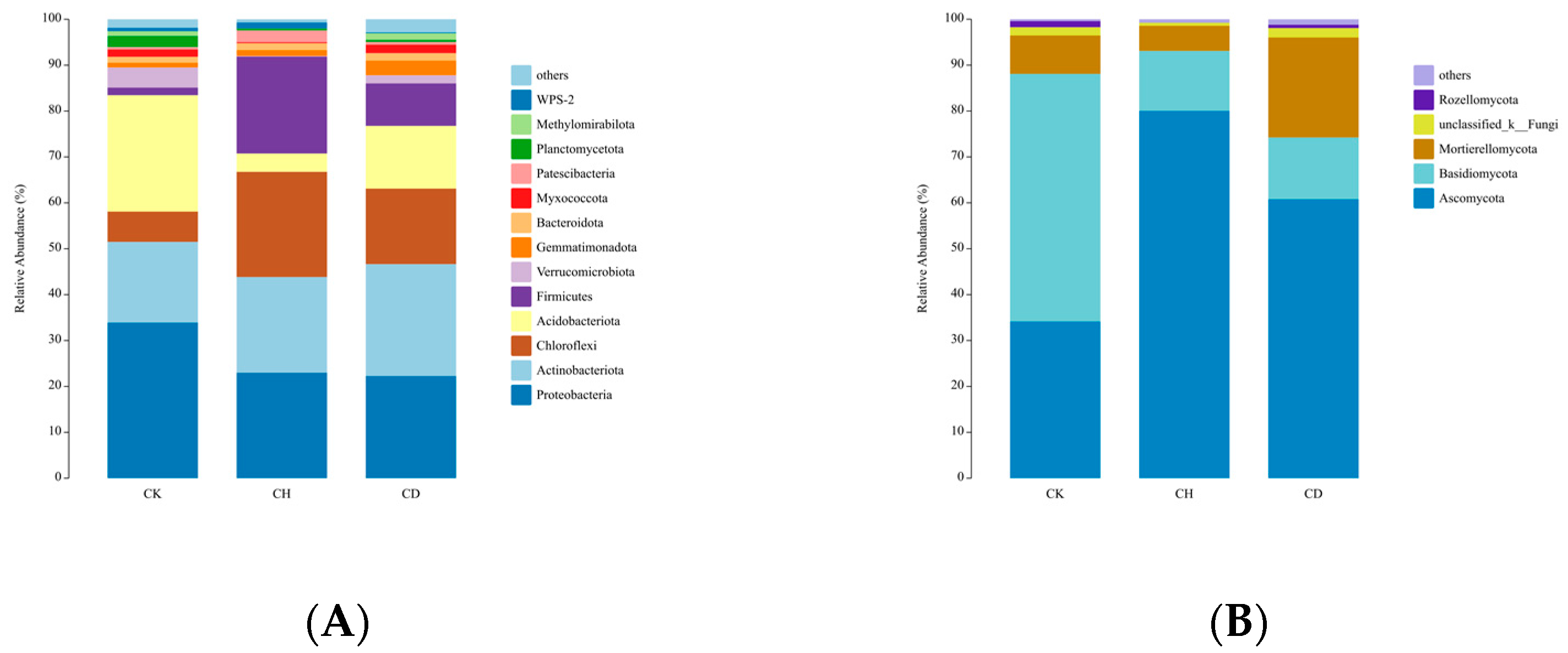
3.4. Soil Bacterial and Fungal Abundance
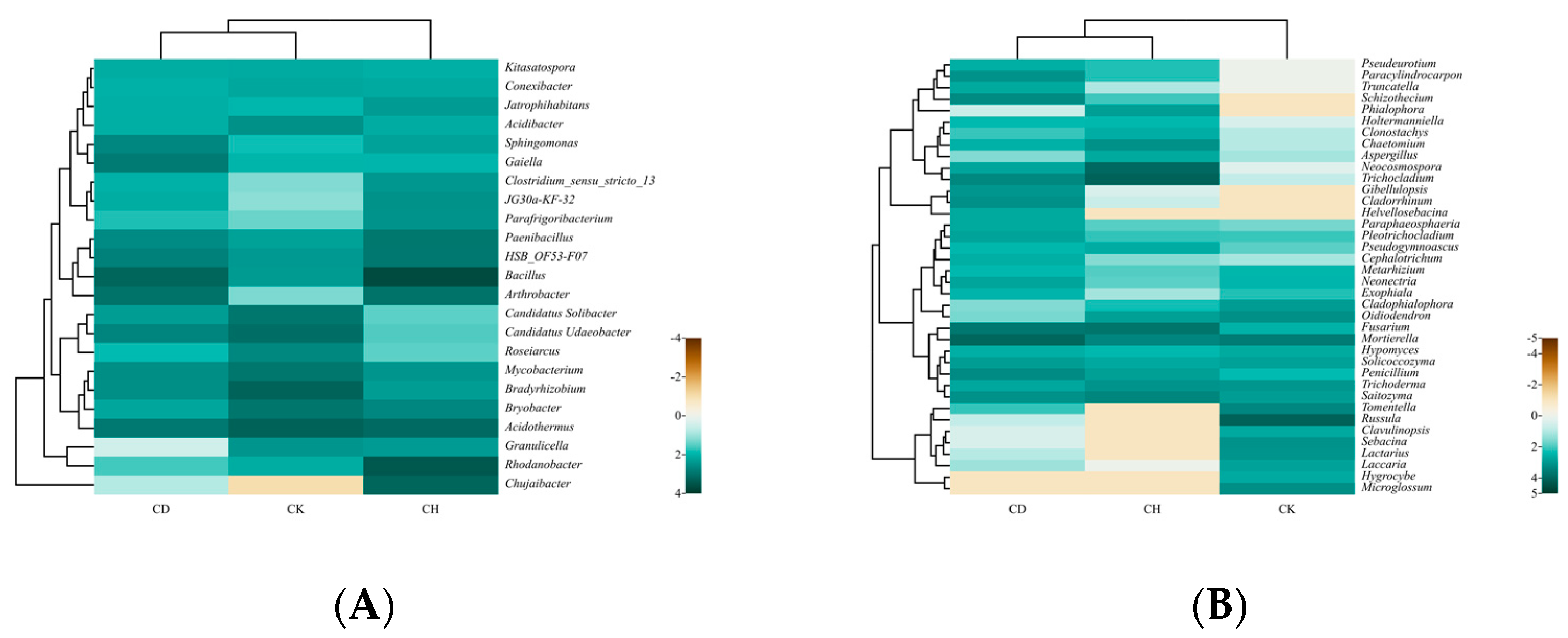
3.5. Soil Microbial Diversity
3.5.1. Alpha Diversity Indices of the Bacterial and Fungal Communities
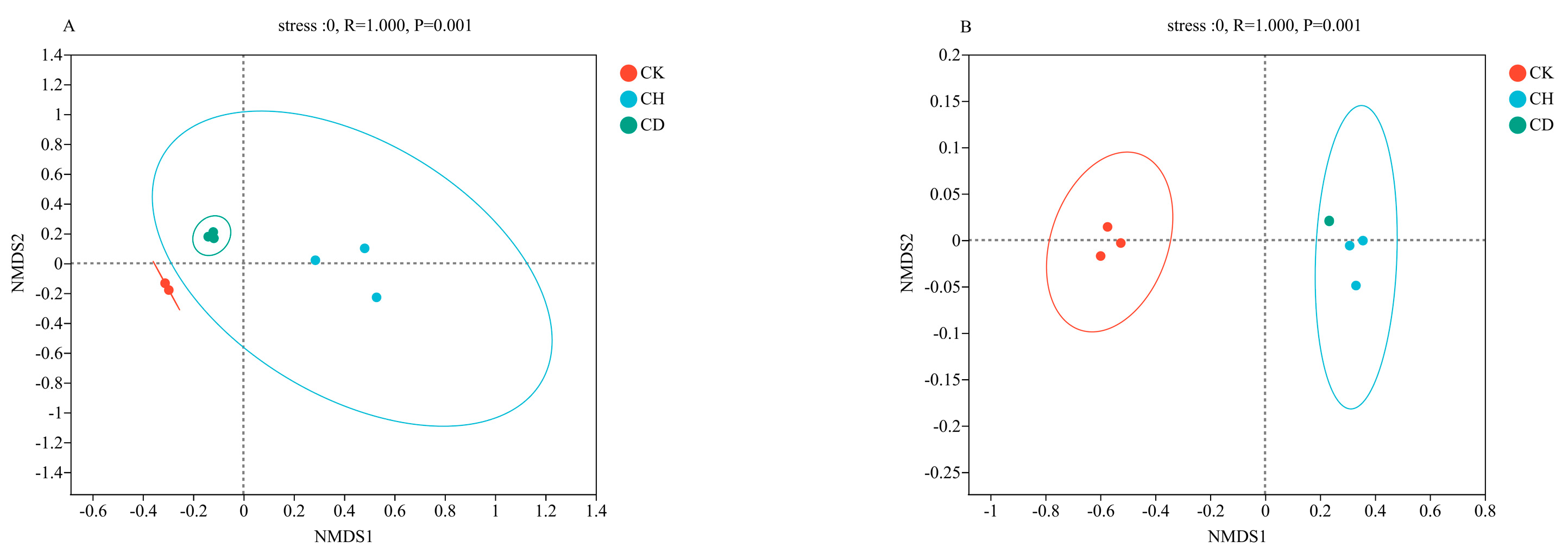
3.5.2. Beta Diversity Indices of the Bacterial and Fungal Communities
3.6. Effects of Environmental Factors on Bacterial and Fungal Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, X.-Y.; Deng, F.-Y.; Zhang, T.-M.; Huang, J.; Chen, C.-N. A new reaction system to determine nonlinear chemical fingerprint and its use in Panax ginseng identification method based on double reaction system. J. Cent. South Univ. 2018, 25, 1895–1903. [Google Scholar] [CrossRef]
- Qi, L.-W.; Wang, C.-Z.; Yuan, C.-S. Isolation and analysis of ginseng: Advances and challenges. Nat. Prod. Rep. 2011, 28, 467–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Yang, Y.; Liu, S.; Yang, S.; Chen, C.; Lin, M.; Zeng, Q.; Long, J.; Yao, J.; Yi, F.; et al. New Therapeutic Approaches to and Mechanisms of Ginsenoside Rg1 against Neurological Diseases. Cells 2022, 11, 2529. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, X.; Liu, X.; Zhang, C.; Shang, W.; Xue, J.; Chen, R.; Xing, Y.; Song, D.; Xu, R. Ginsenoside Rg1 promotes cerebral angiogenesis via the PI3K/Akt/mTOR signaling pathway in ischemic mice. Eur. J. Pharmacol. 2019, 856, 172418. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Wei, B.; Deng, J.; Mi, Y.; Dong, Y.; Zhu, C.; Fu, R.; Qu, L.; Fan, D. The anti-tumor effect of ginsenoside Rh4 in MCF-7 breast cancer cells in vitro and in vivo. Biochem. Biophys. Res. Commun. 2018, 499, 482–487. [Google Scholar] [CrossRef]
- Tong, A.-Z.; Liu, W.; Liu, Q.; Xia, G.-Q.; Zhu, J.-Y. Diversity and composition of the Panax ginseng rhizosphere microbiome in various cultivation modesand ages. BMC Microbiol. 2021, 21, 18. [Google Scholar] [CrossRef]
- Strange, R.N.; Scott, P.R. Plant Disease: A Threat to Global Food Security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef]
- Shin, J.-H.; Yun, B.-D.; Kim, H.-J.; Kim, S.-J.; Chung, D.-Y. Soil Environment and Soil-borne Plant Pathogen Causing Root Rot Disease of Ginseng. Korean J. Soil Sci. Fertil. 2012, 45, 370–376. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ma, Z.; Wang, X.; Sun, Q.; Dong, H.; Wang, G.; Chen, X.; Yin, C.; Han, Z.; Mao, Z. Effects of biochar on the growth of apple seedlings, soil enzyme activities and fungal communities in replant disease soil. Sci. Hortic. 2019, 256, 108641. [Google Scholar] [CrossRef]
- Fujii, T.; Minami, M.; Watanabe, T.; Sato, T.; Kumaishi, K.; Ichihashi, Y. Characterization of inter-annual changes in soil microbial flora of Panax ginseng cultivation fields in Shimane Prefecture of Western Japan by DNA metabarcoding using next-generation sequencing. J. Nat. Med. 2021, 75, 1067–1079. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhao, C.; Chen, Q.; Liu, T.; Li, L.; Liu, X.; Wang, X. Study on Microbial Community Structure and Soil Nitrogen Accumulation in Greenhouse Vegetable Fields with Different Planting Years. Agronomy 2022, 12, 1911. [Google Scholar] [CrossRef]
- Helaoui, S.; Mkhinini, M.; Boughattas, I.; Alphonse, V.; Giusti-Miller, S.; Livet, A.; Banni, M.; Bousserrhine, N. Assessment of Changes on Rhizospheric Soil Microbial Biomass, Enzymes Activities and Bacterial Functional Diversity under Nickel Stress in Presence of Alfafa Plants. Soil Sediment Contam. Int. J. 2020, 29, 823–843. [Google Scholar] [CrossRef]
- Zhang, N.; Nunan, N.; Hirsch, P.R.; Sun, B.; Zhou, J.; Liang, Y. Theory of microbial coexistence in promoting soil–plant ecosystem health. Biol. Fertil. Soils 2021, 57, 897–911. [Google Scholar] [CrossRef]
- Jiao, N.; Song, X.; Song, R.; Yin, D.; Deng, X. Diversity and structure of the microbial community in rhizosphere soil of Fritillaria ussuriensis at different health levels. Peerj 2022, 10, e12778. [Google Scholar] [CrossRef]
- Bao, S.D. Analysis Method of Soil and Agricultural Chemistry, 3rd ed.; China Agricultural Press: Beijing, China, 2000; pp. 25–108. [Google Scholar]
- NY/T53-1987; The Ministry of Agriculture of the People’s Republic of China. Method for the Determination of Soil Total Nitrogen (Semi-Micro Kjeldahl Method). Standards Press of China: Beijing, China, 1987; pp. 157–158.
- NY/T88-1988; The Ministry of Agriculture of the People’s Republic of China. Method for Determination of Soil Total Phosphorus. China Agriculture Press: Beijing, China, 1988; p. 9.
- NY/T87-1988; The Ministry of Agriculture of the People’s Republic of China. Method for Determination of Soil Total Potassium. China Agriculture Press: Beijing, China, 1988; pp. 266–269.
- NY/T1121.7-2014; The Ministry of Agriculture of the People’s Republic of China. Soil Testing-Part 7: Method for Determination of Available Phosphorus in Soil. China Agriculture Press: Beijing, China, 2015; pp. 1–4.
- LY/T1234-2015; National Health and Family Planning Commission of the People’s Republic of China, Forestry Department of People’s Republic of China. Forestry Industry Standards of the People’s Republic of China: Potassium Determination Methods of Forest Soils. Standards Press of China: Beijing, China, 2015; pp. 5–8.
- NY/T1121.6-2006; The Ministry of Agriculture of the People’s Republic of China. Soil Testing-Part 6: Method for Determination of Soil Organic Matter. Standards Press of China: Beijing, China, 2006; pp. 1–3.
- LY/T1228-2015; National Health and Family Planning Commission of the People’s Republic of China, Forestry Department of People’s Republic of China. Forestry Industry Standards of the People’s Republic of China: Nitrogen Determination Methods of Forest Soils. Standards Press of China: Beijing, China, 2015; pp. 7–8.
- LY/T1239-1999; National Health and Family Planning Commission of the People’s Republic of China, Forestry Department of People’s Republic of China. Forestry Industry Standards of the People’s Republic of China: Determination of pH Value in Forest Soils. Standards Press of China: Beijing, China, 1999; pp. 114–115.
- Gao, M.; Song, W.; Zhou, Q.; Ma, X.; Chen, X. Interactive effect of oxytetracycline and lead on soil enzymatic activity and microbial biomass. Environ. Toxicol. Pharmacol. 2013, 36, 667–674. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Witte, C.-P.; Medina-Escobar, N. In-Gel Detection of Urease with Nitroblue Tetrazolium and Quantification of the Enzyme from Different Crop Plants Using the Indophenol Reaction. Anal. Biochem. 2001, 290, 102–107. [Google Scholar] [CrossRef]
- Johansson, L.H.; Borg, L.A.H. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem. 1988, 174, 331–336. [Google Scholar] [CrossRef]
- Sinegani, A.A.S.; Sinegani, M.S. The effects of carbonates removal on adsorption, immobilization and activity of cellulase in a calcareous soil. Geoderma 2012, 173–174, 145–151. [Google Scholar] [CrossRef]
- Guan, S.Y. Soil Enzyme and Its Study Method; China Agriculture Press: Beijing, China, 1986; pp. 270–285. [Google Scholar]
- Dogan, S.; Dogan, M. Determination of kinetic properties of polyphenol oxidase from Thymus (Thymus longicaulis subsp. chaubardii var. chaubardii). Food Chem. 2004, 88, 69–77. [Google Scholar] [CrossRef]
- Liang, Z.; Liu, F.; Wang, W.; Zhang, P.; Sun, X.; Wang, F.; Kell, H. High-throughput sequencing revealed differences of microbial community structure and diversity between healthy and diseased Caulerpa lentillifera. BMC Microbiol. 2019, 19, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, Z.; Arafat, Y.; Lin, W. Studies on fungal communities and functional guilds shift in tea continuous cropping soils by high-throughput sequencing. Ann. Microbiol. 2020, 70, 7. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Li, Y.; Hu, X.; Zang, Q.; Zhuang, H.; Huang, L. The Influence of Organic and Conventional Cultivation Patterns on Physicochemical Property, Enzyme Activity and Microbial Community Characteristics of Paddy Soil. Agriculture 2022, 12, 121. [Google Scholar] [CrossRef]
- Wang, X.; Jia, Z.; Liang, L.; Zhao, Y.; Yang, B.; Ding, R.; Wang, J.; Nie, J. Changes in soil characteristics and maize yield under straw returning system in dryland farming. Field Crop. Res. 2018, 218, 11–17. [Google Scholar] [CrossRef]
- Dong, L.; Xu, J.; Li, Y.; Fang, H.; Niu, W.; Li, X.; Zhang, Y.; Ding, W.; Chen, S. Manipulation of microbial community in the rhizosphere alleviates the replanting issues in Panax ginseng. Soil Biol. Biochem. 2018, 125, 64–74. [Google Scholar] [CrossRef]
- Hao, T.; Zhu, Q.; Zeng, M.; Shen, J.; Shi, X.; Liu, X.; Zhang, F.; de Vries, W. Impacts of nitrogen fertilizer type and application rate on soil acidification rate under a wheat-maize double cropping system. J. Environ. Manag. 2020, 270, 110888. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Xiao, S.; Zhao, Y.; Xu, Y.; Yang, H.; Zhang, L. Comparative analysis of rhizosphere soil physiochemical characteristics and microbial communities between rusty and healthy ginseng root. Sci. Rep. 2020, 10, 15756. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.-M.; Zhang, M.; Zhang, K.-K.; Yang, X.-W.; He, D.-X.; Yin, J.; Wang, C.-Y. Effects of reduced nitrogen and suitable soil moisture on wheat (Triticum aestivum L.) rhizosphere soil microbiological, biochemical properties and yield in the Huanghuai Plain, China. J. Integr. Agric. 2020, 19, 234–250. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, J.; Zhou, X.; Li, F. Effects of shallow groundwater table and fertilization level on soil physico-chemical properties, enzyme activities, and winter wheat yield. Agric. Water Manag. 2018, 208, 307–317. [Google Scholar] [CrossRef]
- Zhao, Q.; Tang, J.; Li, Z.; Yang, W.; Duan, Y. The Influence of Soil Physico-Chemical Properties and Enzyme Activities on Soil Quality of Saline-Alkali Agroecosystems in Western Jilin Province, China. Sustainability 2018, 10, 1529. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Du, J.; Li, Y.; Tang, H.; Yin, Z.; Yang, L.; Ding, X. Evolutions and Managements of Soil Microbial Community Structure Drove by Continuous Cropping. Front. Microbiol. 2022, 13, 839494. [Google Scholar] [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Lee, I.-K.; Yun, B.-S. Antagonistic Effect of Streptomyces sp. BS062 against Botrytis Diseases. Mycobiology 2015, 43, 339–342. [Google Scholar] [CrossRef] [Green Version]
- Bandara, T.; Krohn, C.; Jin, J.; Chathurika, J.; Franks, A.; Xu, J.; Potter, I.D.; Tang, C. The effects of biochar aging on rhizosphere microbial communities in cadmium-contaminated acid soil. Chemosphere 2022, 303, 135153. [Google Scholar] [CrossRef]
- Li, Y.; Dang, H.; Lv, X.; Wang, Z.; Pu, X.; Zhuang, L. High-throughput sequencing reveals rhizosphere fungal community composition and diversity at different growth stages of Populus euphratica in the lower reaches of the Tarim River. Peerj 2022, 10, e13552. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Jeon, E.K.; Hwang, C.-W. Characteristic Analysis of Soil-Isolated Bacillus velezensis HY-3479 and Its Antifungal Activity Against Phytopathogens. Curr. Microbiol. 2022, 79, 357. [Google Scholar] [CrossRef] [PubMed]
- Durairaj, K.; Velmurugan, P.; Park, J.-H.; Chang, W.-S.; Park, Y.-J.; Senthilkumar, P.; Choi, K.-M.; Lee, J.-H.; Oh, B.-T. An investigation of biocontrol activity Pseudomonas and Bacillus strains against Panax ginseng root rot fungal phytopathogens. Biol. Control. 2018, 125, 138–146. [Google Scholar] [CrossRef]
- Kim, S.Y.; An, J.-H.; Park, K.H.; Lee, S.Y.; Weon, H.-Y.; Sang, M.-K.; Lee, J.-H.; Song, J. Effect of Bacillus CC112 Inoculation on Fungal Pathogens and Soil Microbial Community in a Ginseng-Cultivated Soil. Korean J. Soil Sci. Fertil. 2020, 53, 128–139. [Google Scholar] [CrossRef]
- Kim, Y.; Balaraju, K.; Jeon, Y. Biological characteristics of Bacillus amyloliquefaciens AK-0 and suppression of ginseng root rot caused by Cylindrocarpon destructans. J. Appl. Microbiol. 2017, 122, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Naing, K.W.; Anees, M.; Kim, S.J.; Nam, Y.; Kim, Y.C.; Kim, K.Y. Characterization of antifungal activity of Paenibacillus ehimensis KWN38 against soilborne phytopathogenic fungi belonging to various taxonomic groups. Ann. Microbiol. 2014, 64, 55–63. [Google Scholar] [CrossRef]
- Tran-Cong, N.M.; Mándi, A.; Kurtán, T.; Müller, W.E.G.; Kalscheuer, R.; Lin, W.; Liu, Z.; Proksch, P. Induction of cryptic metabolites of the endophytic fungus Trichocladium sp. through OSMAC and co-cultivation. RSC Adv. 2019, 9, 27279–27288. [Google Scholar] [CrossRef] [Green Version]
- Ratledge, C.; Wynn, J.P. The Biochemistry and Molecular Biology of Lipid Accumulation in Oleaginous Microorganisms. Adv. Appl. Microbiol. 2002, 51, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Zhang, N.; Liang, X.; Huang, T.; Li, B. Bacillus aryabhattai LAD impacts rhizosphere bacterial community structure and promotes maize plant growth. J. Sci. Food Agric. 2022, 102, 6650–6657. [Google Scholar] [CrossRef] [PubMed]
- Cagide, C.; Riviezzi, B.; Minteguiaga, M.; Morel, M.A.; Castro-Sowinski, S. Identification of Plant Compounds Involved in the Microbe-Plant Communication during the Coinoculation of Soybean with Bradyrhizobium elkanii and Delftia sp. strain JD2. Mol. Plant-Microbe Interact. 2018, 31, 1192–1199. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Lin, X.; Guan, S.; Dou, S. Deep incorporation of corn straw benefits soil organic carbon and microbial community composition in a black soil of Northeast China. Soil Use Manag. 2022, 38, 1266–1279. [Google Scholar] [CrossRef]
- Guan, Y.M.; Lu, B.H.; Wang, Y.; Gao, J.; Wu, L.J. First Report of Root Rot Caused by Fusarium redolens on Ginseng (Panax ginseng) in Jilin Province of China. Plant Dis. 2014, 98, 844. [Google Scholar] [CrossRef] [PubMed]
- Chacón, F.I.; Sineli, P.E.; Mansilla, F.I.; Pereyra, M.M.; Diaz, M.A.; Volentini, S.I.; Poehlein, A.; Meinhardt, F.; Daniel, R.; Dib, J.R. Native Cultivable Bacteria from the Blueberry Microbiome as Novel Potential Biocontrol Agents. Microorganisms 2022, 10, 969. [Google Scholar] [CrossRef]
- Li, Q.; Yan, N.; Miao, X.; Zhan, Y.; Chen, C. The potential of novel bacterial isolates from healthy ginseng for the control of ginseng root rot disease (Fusarium oxysporum). PLoS ONE 2022, 17, e0277191. [Google Scholar] [CrossRef]
- Kim, S.G.; Khan, Z.; Jeon, Y.H.; Kim, Y.H. Inhibitory Effect of Paenibacillus polymyxa GBR-462 on Phytophthora capsica Causing Phytophthora Blight in Chili Pepper. J. Phytopathol. 2009, 157, 329–337. [Google Scholar] [CrossRef]
- Abbas, A.; Duan, J.; Abdoulaye, A.H.; Fu, Y.; Lin, Y.; Xie, J.; Cheng, J.; Jiang, D. Deciphering Bacterial Community of the Fallow and Paddy Soil Focusing on Possible Biocontrol Agents. Agronomy 2022, 12, 431. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Soil Color | Soil Structure | Soil Texture | Soil Layer | Topographic Position | Soil Taxonomy |
|---|---|---|---|---|---|---|
| CK | dark brown | granular soil | clay | humus layer | plain | phaeozem |
| CH | dark brown | granular soil | clay | humus layer | plain | phaeozem |
| CD | dark brown | granular soil | clay | humus layer | plain | phaeozem |
| Samples | Available Iron (mg·kg−1) | Available Manganese (mg·kg−1) | Cu (mg·kg−1) | Zn (mg·kg−1) | Available Boron (mg·kg−1) | Mo (mg·kg−1) |
|---|---|---|---|---|---|---|
| CK | 233.67 ± 3.51 a | 21.73 ± 0.85 b | 15.50 ± 0.46 b | 55.17 ± 1.46 c | 0.97 ± 0.05 b | 0.55 ± 0.04 c |
| CH | 167.00 ± 3.61 b | 35.67 ± 1.40 a | 14.80 ± 0.20 b | 63.33 ± 0.67 b | 1.38 ± 0.02 a | 0.69 ± 0.02 b |
| CD | 85.70 ± 1.14 c | 23.60 ± 0.36 b | 18.53 ± 0.57 a | 80.30 ± 1.87 a | 0.23 ± 0.02 c | 0.83 ± 0.04 a |
| Samples | TN (mg·kg−1) | TP (mg·kg−1) | TK (mg·kg−1) | AP (mg·kg−1) | AK (mg·kg−1) | HN (mg·kg−1) | OM (mg·kg−1) | pH |
|---|---|---|---|---|---|---|---|---|
| CK | 6100.00 ± 0.00 a | 700.00 ± 0.00 b | 18,600.00 ± 0.00 c | 6.60 ± 0.11 c | 210.67 ± 9.65 c | 398.47 ± 184.17 a | 145,970.00 ± 7.37 a | 7.47 ± 0.07 a |
| CH | 2300.00 ± 0.00 c | 900.00 ± 0.00 a | 27,600.00 ± 0.00 a | 39.25 ± 0.76 a | 545.17 ± 7.20 a | 368.53 ± 5.56 b | 61,930.00 ± 2.88 c | 4.52 ± 0.00 c |
| CD | 4600.00 ± 0.00 b | 1000.00 ± 0.00 a | 26,300.00 ± 0.00 b | 32.05 ± 0.41 b | 287.63 ± 31.70 b | 343.03 ± 13.90 c | 104,230.00 ± 0.26 b | 5.48 ± 0.11 b |
| Samples | S-UE (U·g−1) | S-CAT (U·g−1) | S-CL (U·g−1) | S-SC (U·g−1) | S-ACP (U·g−1) | S-AKP (U·g−1) | S-PPO (U·g−1) |
|---|---|---|---|---|---|---|---|
| CK | 1166.50 ± 186.58 a | 2.88 ± 0.00 a | 6.59 ± 0.01 b | 110.63 ± 3.50 a | 35.34 ± 1.01 a | 30.76 ± 1.23 a | 36.53 ± 0.67 a |
| CH | 557.22 ± 1.40 c | 0.69 ± 0.02 b | 8.32 ± 0.23 a | 15.47 ± 0.01 c | 29.31 ± 0.64 b | 23.07 ± 0.15 c | 28.34 ± 0.30 b |
| CD | 806.13 ± 9.82 b | 2.08 ± 0.00 a | 6.46 ± 0.09 b | 19.96 ± 0.03 b | 34.36 ± 0.39 a | 28.40 ± 0.84 b | 25.85 ± 0.41 c |
| Correlation Coefficient | TN | TP | TK | AP | AK | HN | OM | pH |
|---|---|---|---|---|---|---|---|---|
| S-SC | 0.827 ** | −0.936 ** | −0.994 ** | −0.985 ** | −0.708 * | 0.836 ** | 0.883 ** | 0.946 ** |
| S-UE | 0.975 ** | −0.787 * | −0.956 ** | −0.978 ** | −0.918 ** | 0.605 | 0.993 ** | 0.980 ** |
| S-CL | −0.853 ** | 0.213 | 0.530 | 0.598 | 0.911 ** | 0.017 | −0.796 * | −0.662 |
| S-ACP | 0.939 ** | −0.393 | −0.699 * | −0.748 * | −0.965 ** | 0.151 | 0.904 ** | 0.807 ** |
| S-AKP | 0.973 ** | −0.574 | −0.796 * | −0.847 ** | −0.973 ** | 0.360 | 0.954 ** | 0.871 ** |
| S-CAT | 0.946 ** | −0.382 | −0.670 * | −0.730 * | −0.986 ** | 0.135 | 0.903 ** | 0.787 * |
| S-PPO | 0.645 | −0.960 ** | −0.932 ** | −0.900 ** | −0.495 | 0.908 ** | 0.727 * | 0.852 ** |
| Microbial Community | Kingdom | Phyla | Classes | Orders | Families | Genera | Species |
|---|---|---|---|---|---|---|---|
| bacteria | 1 | 35 | 94 | 243 | 377 | 361 | 338 |
| fungi | 1 | 14 | 50 | 131 | 267 | 431 | 671 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Xie, Y.; Jia, Q.; Li, S.; Li, S.; Shen, N.; Jiang, M.; Wang, Y. Effects of the Continuous Cropping and Soilborne Diseases of Panax Ginseng C. A. Meyer on Rhizosphere Soil Physicochemical Properties, Enzyme Activities, and Microbial Communities. Agronomy 2023, 13, 210. https://doi.org/10.3390/agronomy13010210
Chen F, Xie Y, Jia Q, Li S, Li S, Shen N, Jiang M, Wang Y. Effects of the Continuous Cropping and Soilborne Diseases of Panax Ginseng C. A. Meyer on Rhizosphere Soil Physicochemical Properties, Enzyme Activities, and Microbial Communities. Agronomy. 2023; 13(1):210. https://doi.org/10.3390/agronomy13010210
Chicago/Turabian StyleChen, Fuhui, Yongjun Xie, Qingwen Jia, Shuyan Li, Shiyong Li, Naikun Shen, Mingguo Jiang, and Yibing Wang. 2023. "Effects of the Continuous Cropping and Soilborne Diseases of Panax Ginseng C. A. Meyer on Rhizosphere Soil Physicochemical Properties, Enzyme Activities, and Microbial Communities" Agronomy 13, no. 1: 210. https://doi.org/10.3390/agronomy13010210