
Response of White Cabbage (Brassica oleracea var. capitata) to Single and Repeated Short-Term Waterlogging

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials Used for the Experimental Set-Up
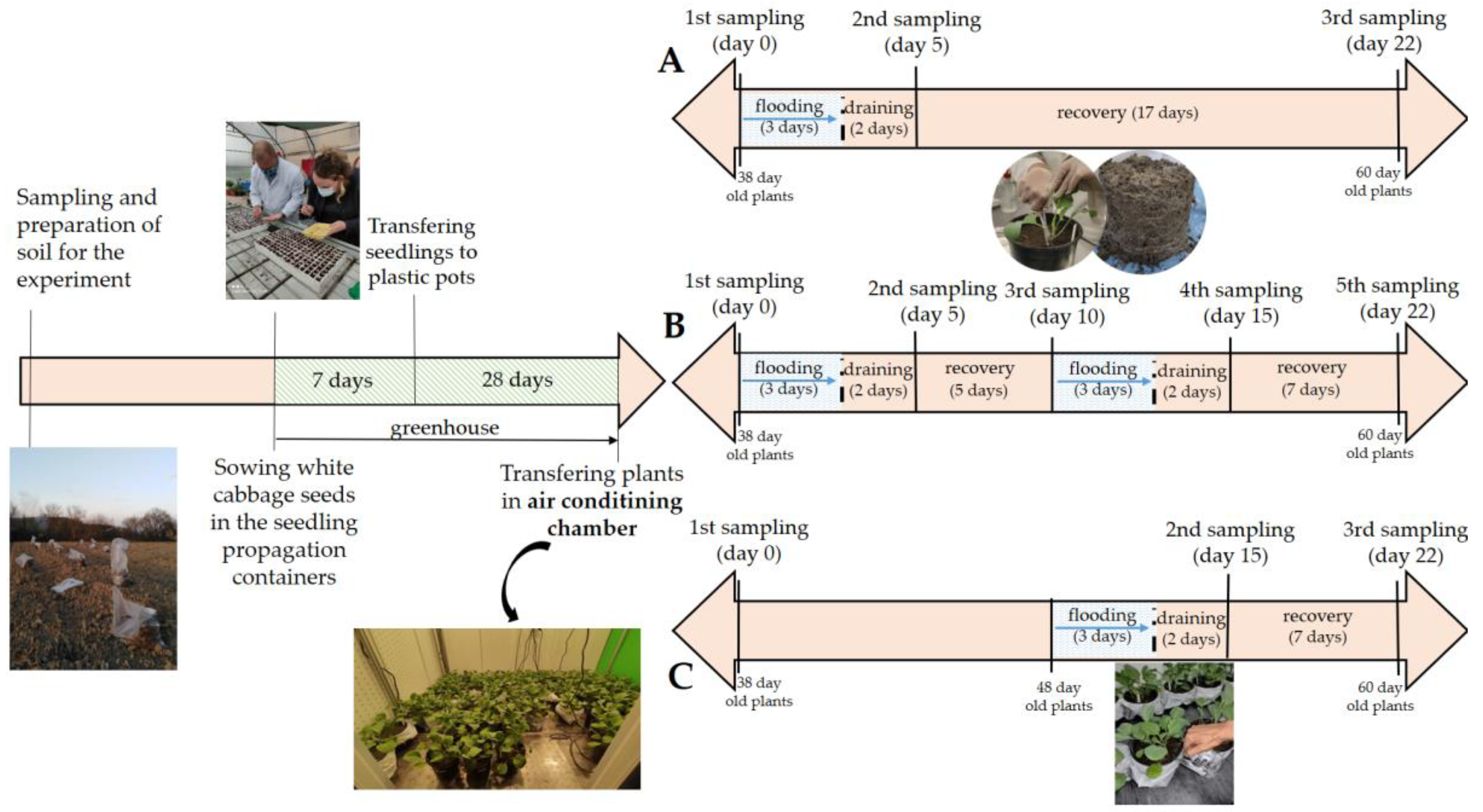
2.2. Experimental Design
2.3. Plant Biomass and Morphological Characteristics
2.4. Chlorophyll Fluorescence and Multispectral Analysis
2.5. Stress Parameter Analysis
2.5.1. Proline
2.5.2. Total Polyphenolic Content
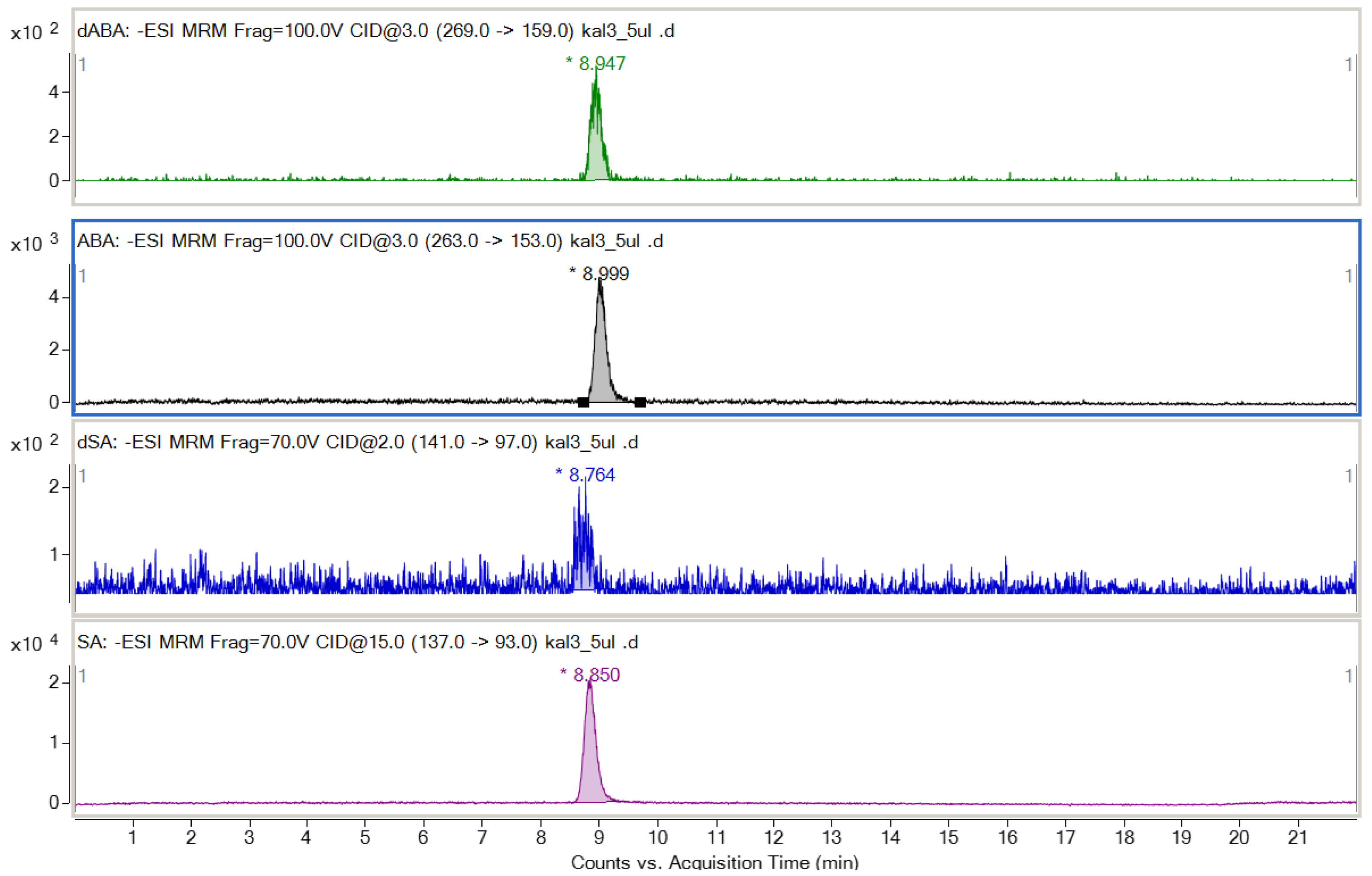
2.5.3. Salicylic and Abscisic Acids
Sample Preparation and Analysis
Preparation of Standard and Calibrant Solutions
2.6. Elements and Mineral Composition Analysis
2.7. Statistical Analysis
3. Results
3.1. Waterlogging Effects on Plant Biomass and Morphological Characteristics
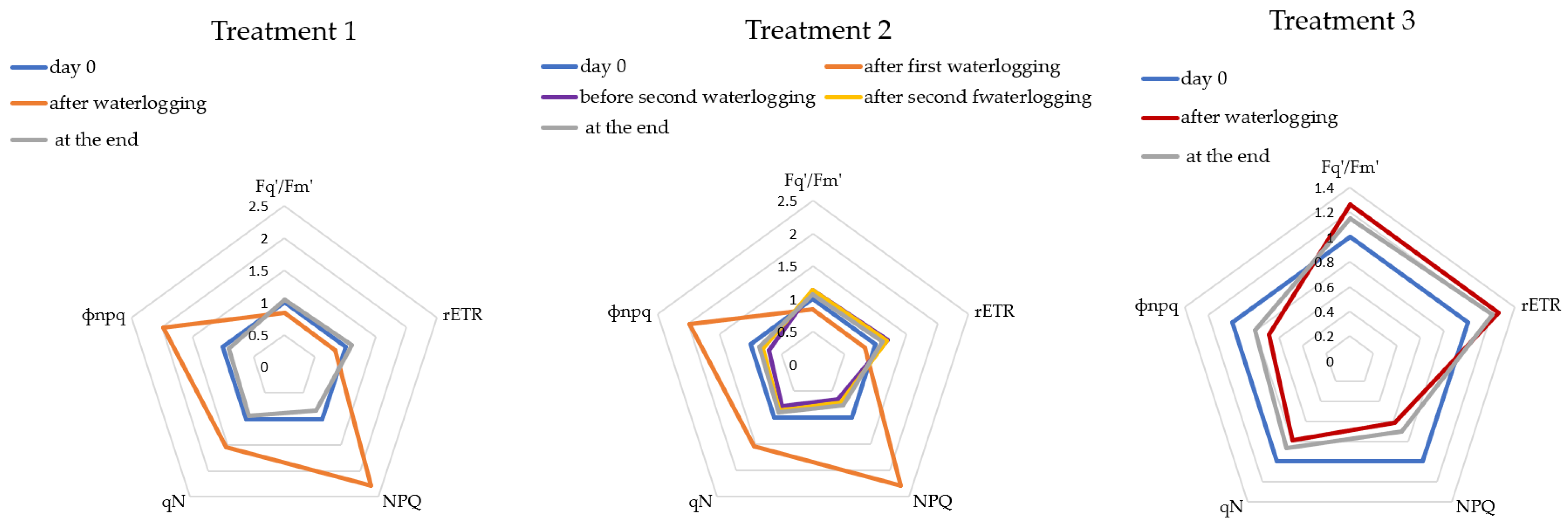
3.2. Waterlogging Effects on Chlorophyll Fluorescence and Multispectral Parameters
3.3. Waterlogging Effects on Plant Molecular Stress Parameters
3.4. Waterlogging Effects on the Concentration of Elements and Minerals in Plants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sly, P.D. Health impacts of climate change and biosecurity in the Asian Pacific region. Rev. Environ. Health 2011, 26, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, D.K.; West, P.C.; Clark, M.; Gerber, J.S.; Prishchepov, A.V.; Chatterjee, S. Climate change has likely already affected global food production. PLoS ONE 2019, 14, e0217148. [Google Scholar] [CrossRef] [PubMed]
- Westra, S.; Fowler, H.J.; Evans, J.P.; Alexander, L.V.; Berg, P.; Johnson, F.; Kendon, E.J.; Lenderink, G.; Roberts, N.M. Future changes tothe intensity and frequency of short-duration extreme rainfall. Rev. Geophys. 2014, 52, 522–555. [Google Scholar] [CrossRef] [Green Version]
- Masson-Delmotte, P.; Zhai, H.O.; Pörtner, D.; Roberts, J.; Skea, P.R.; Shukla, A.; Pirani, W.; Moufouma-Okia, C.; Péan, R.; Pidcock, S.; et al. IPCC Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty. 2018. Available online: https://www.ipcc.ch/sr15/ (accessed on 25 October 2022).
- Mancuso, S.; Shabala, S. Waterlogging Signalling and Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar] [CrossRef]
- Olesen, J.E.; Trnka, M.; Kersebaum, K.C.; Skjelvåg, A.O.; Seguin, B.; Peltonen-Sainio, P.; Rossi, F.; Kozyra, J.; Micale, F. Impacts and adaptation of European crop production systems to climate change. Eur. J. Agron. 2011, 34, 96–112. [Google Scholar] [CrossRef]
- Visser, E.J.W.; Voesenek, L.A.C.J.; Vartapetian, B.B.; Jackson, M.B. Flooding and Plant Growth. Ann. Bot. 2003, 91, 107–109. [Google Scholar] [CrossRef] [Green Version]
- Mustroph, A. Improving Flooding Tolerance of Crop Plants. Agronomy 2018, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Balakhnina, T.I. Plant Responses to Soil Flooding. In Stress Responses in Plants—Mechanisms of Toxicity and Tolerance; Tripathi, B.N., Muller, M., Eds.; Springer International: Cham, Switzerland, 2015; pp. 115–143. [Google Scholar]
- Arduini, I.; Baldanzi, M.; Pampana, S. Reduced Growth and Nitrogen Uptake During Waterlogging at Tillering Permanently Affect Yield Components in Late Sown Oats. Front. Plant Sci. 2019, 12, 1087. [Google Scholar] [CrossRef] [Green Version]
- Petrić, I.; Šamec, D.; Karalija, E.; Salopek-Sondi, B. Beneficial Microbes and Molecules for Mitigation of Soil Salinity in Brassica Species: A Review. Soil Syst. 2022, 6, 18. [Google Scholar] [CrossRef]
- Pavlović, I.; Petřík, I.; Tarkowská, D.; Lepeduš, H.; Vujčić Bok, V.; Radić Brkanac, S.; Novák, O.; Salopek-Sondi, B. Correlations between Phytohormones and Drought Tolerance in Selected Brassica Crops: Chinese Cabbage, White Cabbage and Kale. Int. J. Mol. Sci. 2018, 19, 2866. [Google Scholar] [CrossRef]
- Linić, I.; Šamec, D.; Grúz, J.; Vujčić Bok, V.; Strnad, M.; Salopek-Sondi, B. Involvement of Phenolic Acids in Short-Term Adaptation to Salinity Stress is Species-Specific among Brassicaceae. Plants 2019, 8, 155. [Google Scholar] [CrossRef] [Green Version]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Pavlović, I.; Salopek-Sondi, B. White cabbage (Brassica oleracea var. capitata f. alba): Botanical, phytochemical and pharmacological overview. Phytochem. Rev. 2017, 16, 117–135. [Google Scholar] [CrossRef]
- Food and Organization of the United Nations (FAO). Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 6 December 2022).
- Casierra-Posada, F.; Cutler, J. Photosystem II fluorescence and growth in cabbage plants (Brassica oleracea var. capitata) grown under waterlogging stress. Rev. UDCA Act. Div. Cient. 2017, 20, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Senko, H.; Pole, L.; Mesšić, A.; Šamec, D.; Petek, M.; Pohajda, I.; Rajnović, I.; Udoković-Kolić, N.; Brkljačić, L.; Palijan, G.; et al. Farmers observations on the impact of excessive rain and flooding on agricultural land in Croatia. J. Cent. Eur. Agric. 2022, 23, 125–137. [Google Scholar] [CrossRef]
- HRN ISO 10390:2005; Soil Quality—Determination of pH (ISO 10390:2005). Hrvatski Zavod za Normizaciju: Zagreb, Croatia, 2005.
- Škorić, A. Priručnik za Pedološka Istraživanja; Fakultet Poljoprivrednih Znanosti: Zagreb, Croatia, 1982. [Google Scholar]
- HRN ISO 11261:2004; Soil Quality—Determination of Total Nitrogen—Modified Kjeldahl Method (ISO 11261:1995). Hrvatski Zavod za Normizaciju: Zagreb, Croatia, 2004.
- Egner, H.; Riehm, H.; Domingo, W.R. Untersuchungen uber die chemische Bodenanalyse als Grundlage fur die Beurteilung des Nahrstoffzustandes der Boden II. Chemische Extractionsmethoden zu Phosphor- und Kaliumbestimmung. Kungl. Lantbr. Ann. 1960, 26, 199–215. [Google Scholar]
- Petek, M.; Rehak, M.; Karažija, T.; Viduka, A.; Markoski, M.; Veres, S.; Fruk, G. Potassium and magnesium content in mandarinon market of city of Zagreb. AgroLife Sci. J. 2022, 11, 170–175. [Google Scholar]
- HRN ISO 11277:2011; Soil Quality—Determination of Particle Size Distribution in Mineral Soil Material—Method by Sieving and Sedimentation (ISO 11277:2009). Hrvatski Zavod za Normizaciju: Zagreb, Croatia, 2011.
- HRN EN ISO 11274:2014; Soil Quality—Determination of the Water-Retention Characteristic—Laboratory Methods (ISO 11274:1998+Cor 1:2009; EN ISO 11274:2014). Hrvatski Zavod za Normizaciju: Zagreb, Croatia, 2014.
- Kitajima, M.; Butler, W.L. Fluorescence quenching in photosystem II of chloroplasts. Biochim. Biophys. Acta 1975, 376, 116–125. [Google Scholar]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]
- Schreiber, U.; Schliwa, W.; Bilger, U. Continious recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photsynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Genty, B.; Harbinson, J. Regulation of light utilisation for photosynthetic electron transport. In Environmental Stress and Photosynthesis; Baker, N.R., Ed.; Environmental Stress and Photosynthesis; Kluwer Academic Press: Dordrecht, The Netherlands, 1996; pp. 67–99. [Google Scholar]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for nondestructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Gitelson, A.A.; Merzlyak, M.N.; Chivkunova, O.B. Optical properties and nondestructive estimation of anthocyanin content in plant leaves. Photochem. Photobiol. 2001, 74, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the great plains with ERTS. NASA Spec. Publ. 1974, 1, 309–317. [Google Scholar]
- Carillo, P.; Gibon, Z.; PrometheusWiki Contributors. Extraction and Determination of Proline. Available online: https://www.researchgate.net/publication/211353600_PROTOCOL_Extraction_and_determination_of_proline (accessed on 21 December 2022).
- Waterhouse, A.L. Determination of Total Phenolics. Curr. Protoc. Food Anal. Chem. 2001, 6, I1.1.1–I1.1.8. [Google Scholar]
- HRN ISO 11465:2004; Soil Quality—Determination of Dry Matter and Water Content on a Mass Basis—Gravimetric Method (ISO 11465:1993+Cor 1:1994). Hrvatski Zavod za Normizaciju: Zagreb, Croatia, 2004.
- AOAC. Official Method of Analysis of AOAC International; AOAC: Gaithersburg, MD, USA, 2015. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Paul, P.L.C.; Bell, R.W.; Barrett-Lennard, E.G.; Kabir, E.; Mainuddin, M.; Sarker, K.K. Short-Term Waterlogging Depresses Early Growth of Sunflower (Helianthus annuus L.) on Saline Soils with a Shallow Water Table in the Coastal Zone of Bangladesh. Soil Syst. 2021, 5, 68. [Google Scholar] [CrossRef]
- Lenssen, J.P.M.; Menting, F.B.J.; Van der Putten, W.H. Plant responses to simultaneous stress of waterlogging and shade: Amplified or hierarchical effects? New Phytol. 2003, 157, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Noguchi, K. Tolerant mechanisms to O2 deficiency under submergence conditions in plants. J. Plant Res. 2020, 133, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Vashisht, D.; Hesselink, A.; Pierik, R.; Ammerlaan, J.M.H.; Bailey-Serres, J.; Visser, E.J.W.; Pedersen, O.; van Zanten, M.; Vreugdenhil, D.; Jamar, D.C.L.; et al. Natural variation of submergence tolerance among Arabidopsis thaliana accessions. New Phytol. 2011, 190, 299–310. [Google Scholar] [CrossRef]
- Dubois, M.; Van den Broeck, L.; Inzé, D. The Pivotal Role of Ethylene in Plant Growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanwar, R.S.; Baker, J.L.; Mukhtar, S. Excessive soilwater effects at various stages of development on the growth and yield of corn. Trans. ASAE 1988, 31, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Ren, B.; Zhang, J.; Li, X.; Fan, X.; Dong, S.; Liu, P.; Zhao, B. Effects of waterlogging on the yield and growth of summer maize under field conditions. Can. J. Plant Sci. 2014, 94, 23–31. [Google Scholar] [CrossRef]
- Mcdaniel, V.; Skaggs, R.W.; Negm, L. Injury and Recovery of Maize Roots Affected by Flooding. Appl. Eng. Agric. 2016, 32, 627–638. [Google Scholar] [CrossRef]
- Yamauchi, T.; Colmer, T.D.; Pedersen, O.; Nakazono, M. Regulation of Root Traits for Internal Aeration and Tolerance to Soil Waterlogging—Flooding Stress. Plant Physiol. 2018, 176, 1118–1130. [Google Scholar] [CrossRef]
- Wang, X.; Deng, Z.; Zhang, W.; Meng, Z.; Chang, X.; Lv, M. Effect of Waterlogging Duration at Different Growth Stages on the Growth, Yield and Quality of Cotton. PLoS ONE 2017, 12, e0169029. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Mustroph, A.; Sasidharan, R.; Vashisht, D.; Pedersen, O.; Oosumi, T.; Voesenek, L.A.C.J.; Bailey-Serres, J. Molecular characterization of the submergence response of the Arabidopsis thaliana ecotype Columbia. New Phytol. 2011, 190, 457–471. [Google Scholar] [CrossRef]
- Vriezen, W.H.; Zhou, Z.; Van der Straeten, D. Regulation of Submergence-induced Enhanced Shoot Elongation in Oryza sativa L. Ann. Bot. 2003, 91, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Šamec, D.; Linić, I.; Salopek-Sondi, B. Salinity Stress as an Elicitor for Phytochemicals and Minerals Accumulation in Selected Leafy Vegetables of Brassicaceae. Agronomy 2021, 11, 361. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Caudle, K.L.; Maricle, B.R. Effects of Flooding on Photosynthesis, Chlorophyll Fluorescence, and Oxygen Stress in Plants of Varying Flooding Tolerance. Trans. Kans. Acad. Sci. 2012, 115, 5–18. Available online: http://www.jstor.org/stable/23264070 (accessed on 15 November 2022).
- Zahra, N.; Hafeez, M.B.; Shaukat, K.; Shaukat, K.; Wahid, A.; Hussain, S.; Naseer, R.; Raza, A.; Iqbal, S.; Farooq, M. Hypoxia and Anoxia Stress: Plant responses and tolerance mechanisms. J. Agron. Crop. Sci. 2021, 207, 249–284. [Google Scholar] [CrossRef]
- Mommer, L.; Visser, E.J.W. Underwater photosynthesis in flooded terrestrial plants: A matter of leaf plasticity. Ann. Bot. 2005, 96, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological Responses to Drought, Salinity, and Heat Stress in Plants: A Review. Stresses 2022, 2, 9. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol Res. 2019, 52, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljubej, V.; Karalija, E.; Salopek-Sondi, B.; Šamec, D. Effects of Short-Term Exposure to Low Temperatures on Proline, Pigments, and Phytochemicals Level in Kale (Brassica oleracea var. acephala). Horticulturae 2021, 7, 341. [Google Scholar] [CrossRef]
- Balakhnina, T.; Bennicelli, R.; Stêpniewska, Z.; Stêpniewski, W.; Borkowska, A.; Fomina, I. Stress responses of spring rape plants to soil flooding. Int. Agrophys. 2012, 26, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Teoh, E.Y.; Teo, C.H.; Baharum, N.A.; Pua, T.-L.; Tan, B.C. Waterlogging Stress Induces Antioxidant Defense Responses, Aerenchyma Formation and Alters Metabolisms of Banana Plants. Plants 2022, 11, 2052. [Google Scholar] [CrossRef]
- Yiu, J.C.; Liu, C.W.; Fang, D.Y.-T.; Lai, Y.S. Waterlogging tolerance of Welsh onion (Allium fistulosum L.) enhanced by exogenous spermidine and spermine. Plant Physiol. Biochem. 2009, 47, 710–716. [Google Scholar] [CrossRef]
- Khan, M.; Iqbal, R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Hwang, S.J.; Waqas, M.; Khan, A.L.; Lee, J.H. Comparative analysis of endogenous hormones level in two soybean (Glycine max L.) lines differing in waterlogging tolerance. Front. Plant Sci. 2015, 6, 714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, U.; Gulati, I.; Rathiya, G.; Singh, P. Effect of Saline Water Irrigation, Humic Acid and Salicylic Acid on Soil Properties, Yield Attributes and Yield of Tomato (Lycopersicon esculentum Mill.). Environ. Ecol. 2019, 35, 449–453. [Google Scholar]
- Wang, J.; Shi, S.H.; Wang, D.Y.; Sun, Y.; Zhu, M.; Li, F.H. Exogenous salicylic acid ameliorates waterlogging stress damages and improves photosynthetic efficiency and antioxidative defense system in waxy corn. Photosynthetica 2021, 59, 84–94. [Google Scholar] [CrossRef]
- Hasanuzzamana, M.; Ahmeda, N.; Sahaa, T.; Rahmana, M.; Rahmana, K.; Alama, M.M.; Rohman, M.; Nahar, K. Exogenous salicylic acid and kinetin modulate reactive oxygen species metabolism and glyoxalase system to confer waterlogging stress tolerance in soybean (Glycine max L.). Plant Stress 2022, 3, 100057. [Google Scholar] [CrossRef]
- Sharma, A.; Thakur, S.; Kumar, V.; Kanwar, M.K.; Kesavan, A.K.; Thukral, A.K.; Bhardwaj, R.; Alam, P.; Ahmad, P. Pre-sowing Seed Treatment with 24-Epibrassinolide Ameliorates Pesticide Stress in Brassica juncea L. through the Modulation of Stress Markers. Front. Plant Sci. 2016, 7, 1569. [Google Scholar] [CrossRef] [Green Version]
- Ljubej, V.; Radojčić Redovniković, I.; Salopek-Sondi, B.; Smolko, A.; Roje, S.; Šamec, D. Chilling and Freezing Temperature Stress Differently Influence Glucosinolates Content in Brassica oleracea var. acephala. Plants 2021, 10, 1305. [Google Scholar] [CrossRef]
- Šola, I.; Stić, P.; Rusak, G. Effect of flooding and drought on the content of phenolics, sugars, photosynthetic pigments and vitamin C, and antioxidant potential of young Chinese cabbage. Eur. Food Res. Technol. 2021, 247, 1913–1920. [Google Scholar] [CrossRef]
- Zaidi, P.H.; Rafique, S.; Singh, N.N. Response of maize (Zea mays L.) genotypes to excess moisture stress: Morpho-physiological effects and basis of tolerance. Eur. J. Agron. 2003, 19, 383–399. [Google Scholar]
- Abiko, T.; Kotula, L.; Shiono, K.; Malik, A.I.; Colmer, T.D.; Nakazono, M. Enhanced formation of aerenchyma and induction of a barrier to radial oxygen loss in adventitious roots of Zea nicaraguensis contribute to its waterlogging tolerance as compared with maize (Zea mays ssp. mays). Plant Cell Environ. 2012, 35, 1618–1630. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Fukao, T.; Gibbs, D.J.; Holdsworth, M.J.; Lee, S.C.; Licausi, F.; Perata, P.; Voesenek, L.A.; van Dongen, J.T. Making sense of low oxygen sensing. Trends Plant Sci. 2012, 17, 129–138. [Google Scholar] [CrossRef]
- Setter, T.; Waters, I.; Sharma, S.; Singh, K.N.; Kulshreshtha, N.; Yaduvanshi, N.P.S.; Ram, P.C.; Singh, B.N.; Rane, J.; McDonald, G.; et al. Review of wheat improvement for waterlogging tolerance in Australia and India: The importance of anaerobiosis and element toxicities associated with different soils. Ann. Bot. 2008, 103, 221–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzog, M.; Striker, G.G.; Colmer, T.D.; Pedersen, O. Mechanisms of waterlogging tolerance in wheat—A review of root and shoot physiology. Plant Cell Environ. 2016, 39, 1068–1086. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| pH | % | AL-mg/100 g | % | |||
|---|---|---|---|---|---|---|
| H2O | nKCl | Humus | N | P2O5 | K2O | CaCO3 |
| 7.55 | 7.25 | 5.58 | 0.332 | 44.1 | 47.5 | 14.8 |
| Content of Mechanical Particles in the Soil (%) | Textural Class | ||||
|---|---|---|---|---|---|
| Coarse Sand 2.0–0.2 mm | Fine Sand 0.2–0.063 mm | Coarse Silt 0.063–0.02 mm | Fine Silt 0.02–0.002 Mm | Clay <0.002 mm | |
| 1.7 | 4.8 | 29.5 | 49.0 | 15.0 | Silty loam |
| Soil Water Retention (% Weight) at | Physiologically Active Moisture 3 | |
|---|---|---|
| 0.33 bar 1 | 15.0 bar 2 | % weight |
| 25.4 | 14.4 | 11.0 |
| Mass of Aerial Parts (g) | Average Leaf Surface (cm2) | Average Leaf Blade Width (cm) | Average Leaf Length (cm) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Day 0 | 17.70 ± 1.25 | 41.80 ± 1.58 | 9.10 ± 0.30 | 10.91 ± 0.79 | |||||
| Control | Waterlogged Samples | Control | Waterlogged Samples | Control | Waterlogged Samples | Control | Waterlogged Samples | ||
| Treatment 1 | After waterlogging | 22.14 ± 2.79 | 23.22 ± 3.83 | 41.00 ± 4.82 | 38.60 ± 8.35 | 9.50 ± 0.30 | 9.50 ± 1.07 | 9.91 ± 0.71 | 9.42 ± 1.27 |
| At the end | 26.32 ± 2.54 | 28.75 ± 5.03 | 43.50 ± 5.92 | 42.70 ± 5.57 | 10.30 ± 0.46 | 11.70 ± 3.62 | 9.40 ± 1.12 | 8.00 ± 0.75 | |
| Treatment 2 | Before the second waterlogging | 26.04 ± 3.28 | 23.58 ± 2.72 | 46.21 ± 2.94 | 35.15 ± 4.23 * | 10.62 ± 0.43 | 9.69 ± 0.43 | 10.11 ± 1.21 | 8.60 ± 1.46 |
| After the second waterlogging | 24.99 ± 2.21 | 25.12 ± 1.47 | 40.56 ± 4.32 | 35.85 ± 3.84 | 10.50 ± 0.51 | 9.96 ± 0.44 | 9.53 ± 0.32 | 8.64 ± 0.53 | |
| At the end | 26.32 ± 2.54 | 28.09 ± 2.20 | 43.50 ± 5.92 | 43.31 ± 1.99 | 10.30 ± 0.46 | 10.69 ± 0.88 | 9.40 ± 1.12 | 8.95 ± 0.36 | |
| Treatment 3 | After waterlogging | 24.99 ± 2.21 | 28.99 ± 4.65 | 40.56 ± 4.32 | 46.80 ± 5.45 | 10.50 ± 0.51 | 10.70 ± 0.73 | 9.53 ± 0.32 | 9.48 ± 0.49 |
| At the end | 26.32 ± 2.54 | 27.92 ± 2.78 | 43.50 ± 5.92 | 46.70 ± 1.31 | 10.30 ± 0.46 | 10.20 ± 0.23 | 9.40 ± 1.12 | 9.70 ± 0.18 | |
| Proline (µmol/g DW) | Total Polyphenols (mg GAE/g DW) | ABA (ng/g DW) | SA (ng/g DW) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Day 0 | 3.35 ± 0.03 | 17.68 ± 0.88 | 87.30 ± 16.85 | 468.21 ± 30.22 | |||||
| Control | Waterlogged Samples | Control | Waterlogged Samples | Control | Waterlogged Samples | Control | Waterlogged Samples | ||
| Treatment 1 | After waterlogging | 5.28 ± 1.94 | 3.95 ± 0.87 | 15.82 ± 2.44 | 15.81 ± 0.84 | 206.72 ± 52.71 | 299.09 ± 65.20 | 2232.27 ± 1161.77 | 1167.69 ± 522.91 |
| At the end | 3.18 ± 0.51 | 3.73 ± 1.51 | 12.75 ± 0.80 | 9.33 ± 2.61 | 184.89 ± 123.43 | 286.58 ± 132.00 | 1622.09 ± 525.56 | 1861.55 ± 280.73 | |
| Treatment 2 | Before the second waterlogging | 4.33 ± 0.94 | 5.96 ± 1.24 | 13.90 ± 0.80 | 16.92 ± 1.28 * | 211.63 ± 39.42 | 203.75 ± 49.83 | 1054.88 ± 65.91 | 946.42 ± 119.46 |
| After the second waterlogging | 8.40 ± 1.50 | 4.51 ± 0.90 ** | 12.73 ± 2.99 | 14.48 ± 1.24 | 778.88 ± 6.45 | 359.68 ± 40.95 * | 1115.15 ± 303.97 | 2283.27 ± 432.92 * | |
| At the end | 3.18 ± 0.51 | 3.23 ± 0.54 | 9.65 ± 0.67 | 11.47 ± 0.80 * | 184.89 ± 123.43 | 347.47 ± 67.19 | 1622.089 ± 525.56 | 1538.58 ± 165.92 | |
| Treatment 3 | After waterlogging | 8.40 ± 1.50 | 4.77 ± 0.61 ** | 12.73 ± 2.99 | 13.57 ± 0.70 | 778.88 ± 6.45 | 309.27 ± 165.58 | 1115.15 ± 303.97 | 198.12 ± 51.22 ** |
| At the end | 3.18 ± 0.51 | 3.99 ± 0.44 | 9.65 ± 0.67 | 9.75 ± 1.41 | 184.89 ± 123.43 | 149.16 ± 61.63 | 1622.089 ± 525.56 | 890.89 ± 211.33 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huđ, A.; Šamec, D.; Senko, H.; Petek, M.; Brkljačić, L.; Pole, L.; Lazarević, B.; Rajnović, I.; Udiković-Kolić, N.; Mešić, A.; et al. Response of White Cabbage (Brassica oleracea var. capitata) to Single and Repeated Short-Term Waterlogging. Agronomy 2023, 13, 200. https://doi.org/10.3390/agronomy13010200
Huđ A, Šamec D, Senko H, Petek M, Brkljačić L, Pole L, Lazarević B, Rajnović I, Udiković-Kolić N, Mešić A, et al. Response of White Cabbage (Brassica oleracea var. capitata) to Single and Repeated Short-Term Waterlogging. Agronomy. 2023; 13(1):200. https://doi.org/10.3390/agronomy13010200
Chicago/Turabian StyleHuđ, Anastazija, Dunja Šamec, Helena Senko, Marko Petek, Lidija Brkljačić, Lucia Pole, Boris Lazarević, Ivana Rajnović, Nikolina Udiković-Kolić, Armin Mešić, and et al. 2023. "Response of White Cabbage (Brassica oleracea var. capitata) to Single and Repeated Short-Term Waterlogging" Agronomy 13, no. 1: 200. https://doi.org/10.3390/agronomy13010200