
Agro-Physiological Indices and Multidimensional Analyses for Detecting Heat Tolerance in Wheat Genotypes

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Plant Materials
2.2. Measurements of Traits and Data Collection
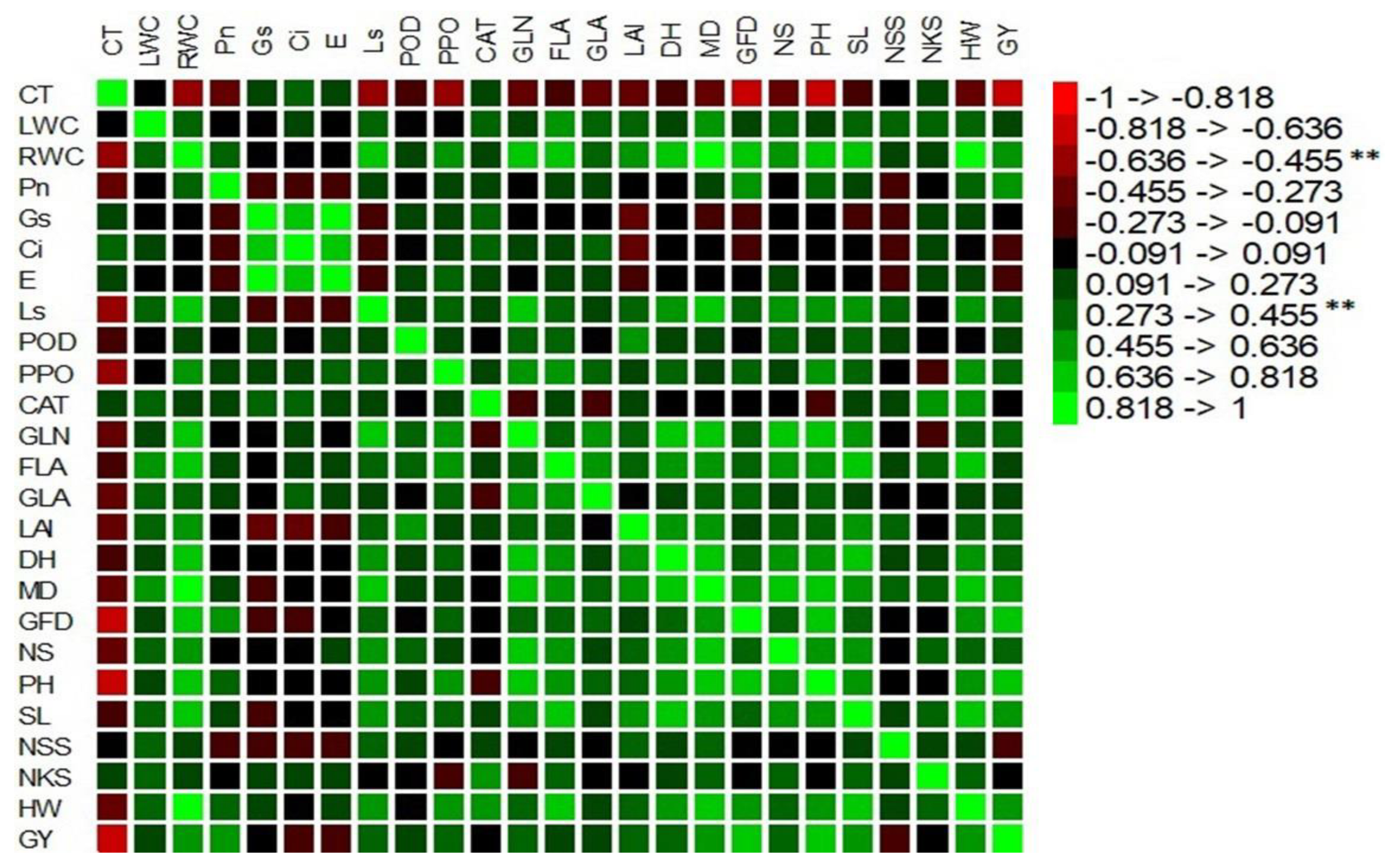
2.3. Statistical Analysis of Evaluated Data
3. Results
3.1. Phenotypic Analysis of Heat Tolerance Index
3.1.1. Analysis of Variance and Genetic Parameters of the Studied Indices
3.1.2. Heat Tolerance Index of Measured Parameters
3.2. Multidimensional Analyses in the Classification of Heat-Tolerant Genotypes
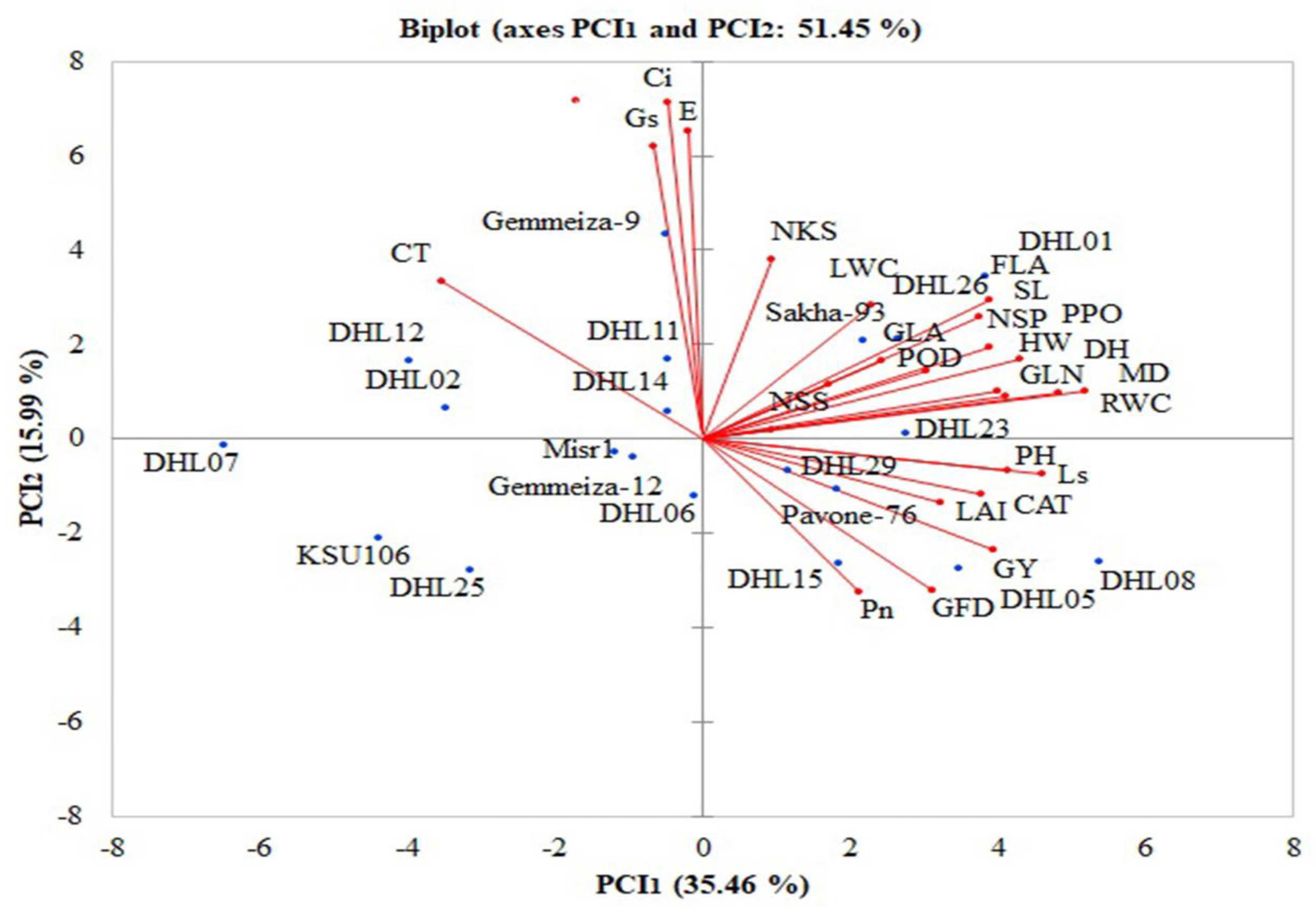
3.2.1. Principal Component Analysis
3.2.2. Identification of Indices Related to Yield Tolerance Index
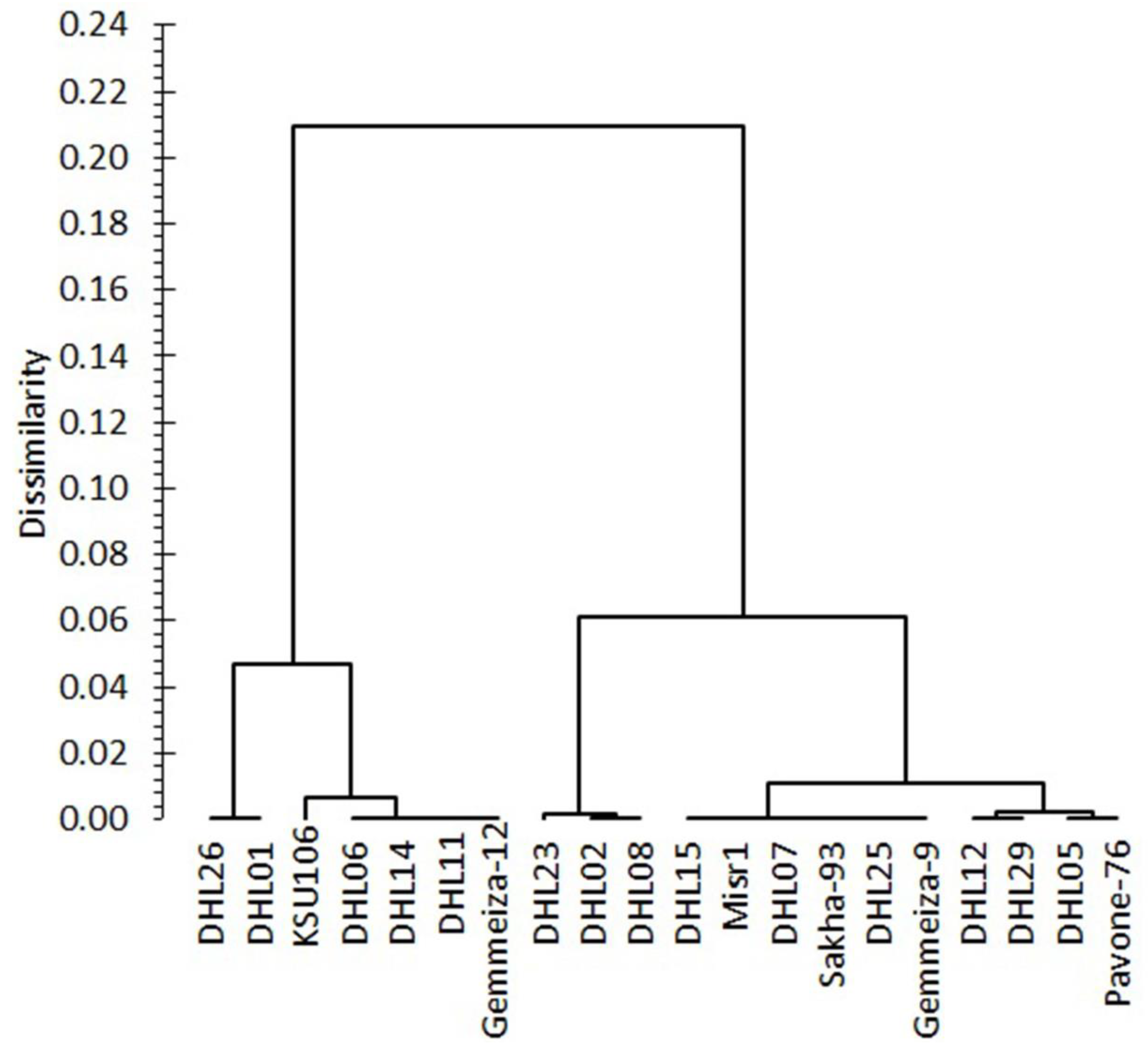
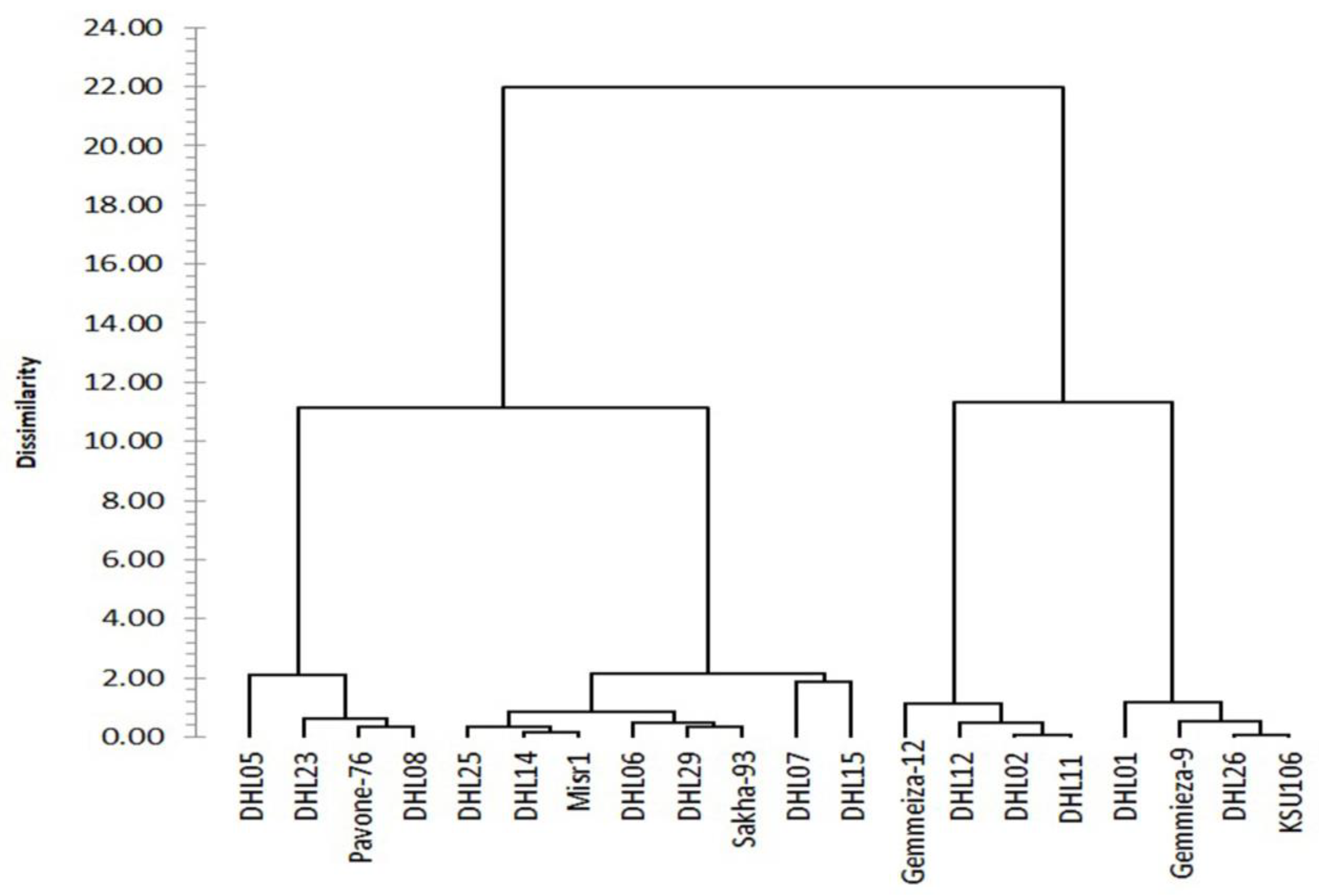
3.2.3. Clustering and Genetic Relationships between the Genotypes for Heat Tolerance
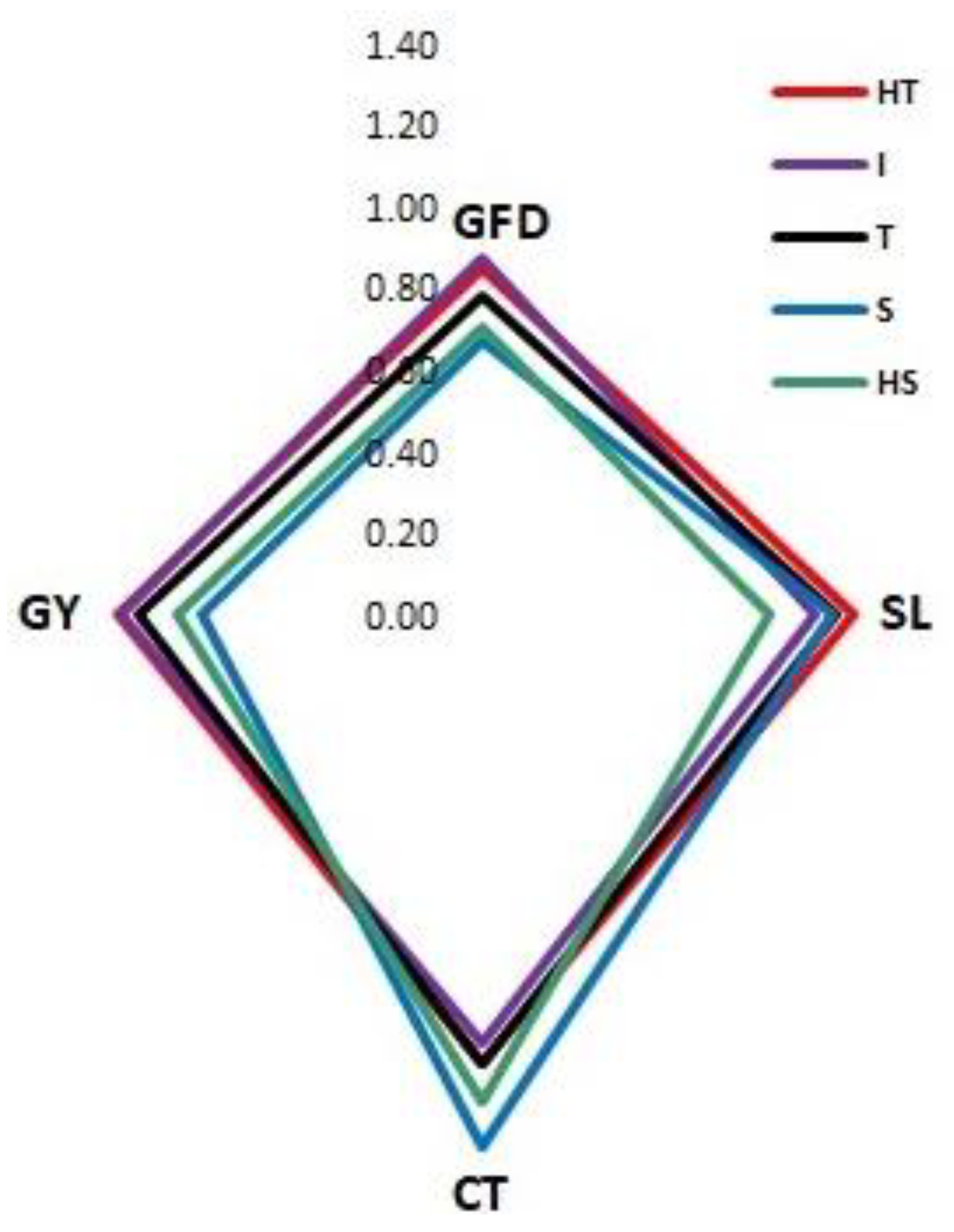
3.2.4. Differentiation of Heat Groups by Discriminant Function Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Ashkar, I.; Alotaibi, M.; Refay, Y.; Ghazy, A.; Zakri, A.; Al-Doss, A. Selection criteria for high-yielding and early-flowering bread wheat hybrids under heat stress. PLoS ONE 2020, 15, e0236351. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.W.; Yang, L.; Yousaf, M.I.; Sami, A.; Mei, X.D.; Shah, L.; Rehman, S.; Xue, L.; Si, H.; Ma, C. Effects of Heat Stress on Growth, Physiology of Plants, Yield and Grain Quality of Different Spring Wheat (Triticum aestivum L.) Genotypes. Sustainability 2021, 13, 2972. [Google Scholar] [CrossRef]
- Teixeira, E.I.; Fischer, G.; van Velthuizen, H.; Walter, C.; Ewert, F. Global hot-spots of heat stress on agricultural crops due to climate change. Agric. Forest Meteorol. 2013, 170, 206–215. [Google Scholar] [CrossRef]
- Dubey, R.; Pathak, H.; Chakrabarti, B.; Singh, S.; Gupta, D.K.; Harit, R.C. Impact of terminal heat stress on wheat yield in India and options for adaptation. Agric. Syst. 2020, 181, 102826. [Google Scholar] [CrossRef]
- Fernie, E.; Tan, D.K.; Liu, S.Y.; Ullah, N.; Khoddami, A.J.A. Post-Anthesis Heat Influences Grain Yield, Physical and Nutritional Quality in Wheat: A Review. Agriculture 2022, 12, 886. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Sallam, M.; Al-Suhaibani, N.; Ibrahim, A.; Alsadon, A.; Al-Doss, A.J.A. Multiple Stresses of Wheat in the Detection of Traits and Genotypes of High-Performance and Stability for a Complex Interplay of Environment and Genotypes. Agronomy 2022, 12, 2252. [Google Scholar] [CrossRef]
- Tariq, A.; Ashraf, I.; Ahmed, M.; Muscolo, A.; Basra, S.; Reynolds, M. Evaluation of Physiological and Morphological Traits for Improving Spring Wheat Adaptation to Terminal Heat Stress. Plants 2021, 10, 455. [Google Scholar]
- Djanaguiraman, M.; Narayanan, S.; Erdayani, E.; Prasad, P.V.V. Effects of high temperature stress during anthesis and grain filling periods on photosynthesis, lipids and grain yield in wheat. BMC Plant Biol. 2020, 20, 268. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.K.; Mishra, B.; Chatrath, R.; Ferrara, G.O.; Singh, R.P. Wheat improvement in India: Present status, emerging challenges and future prospects. Euphytica 2007, 157, 431–446. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef]
- Asseng, S.; Foster, I.; Turner, N.C. The impact of temperature variability on wheat yields. Glob. Chang. Biol. 2011, 17, 997–1012. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Boyle, D.L.; Welti, R.; Jagadish, S.V.K.; Prasad, P.V.V. Decreased photosynthetic rate under high temperature in wheat is due to lipid desaturation, oxidation, acylation, and damage of organelles. BMC Plant Biol. 2018, 18, 55. [Google Scholar] [CrossRef] [Green Version]
- Yadav, M.R.; Choudhary, M.; Singh, J.; Lal, M.K.; Jha, P.K.; Udawat, P.; Gupta, N.K.; Rajput, V.D.; Garg, N.K.; Maheshwari, C.; et al. Impacts, Tolerance, Adaptation, and Mitigation of Heat Stress on Wheat under Changing Climates. Int. J. Mol. Sci. 2022, 23, 2838. [Google Scholar] [CrossRef]
- Tashiro, T.; Wardlaw, I.F. The Response to High-Temperature Shock and Humidity Changes Prior to and during the Early Stages of Grain Development in Wheat. Aust. J. Plant Physiol. 1990, 17, 551–561. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P. Stay-green in spring wheat can be determined by spectral reflectance measurements (normalized difference vegetation index) independently from phenology. J. Exp. Bot. 2012, 63, 3789–3798. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Singh, R.P.; Crossa, J.; Huerta-Espino, J.; Sharma, I.; Chatrath, R.; Singh, G.P.; Sohu, V.S.; Mavi, G.S.; Sukuru, V.S.P.; et al. Earliness in wheat: A key to adaptation under terminal and continual high temperature stress in South Asia. Field Crop. Res. 2013, 151, 19–26. [Google Scholar] [CrossRef]
- Barakat, M.N.; Al-Doss, A.A.; Moustafa, K.A.; Motawei, M.I.; Al-Ashkar, I.M.; Al-Otayk, S.M.; Alamri, M.S.; Mergoum, M. Mapping Wheat Qtls for Grain Yield Related Traits under High Temperature Stress. Genetika 2020, 52, 1107–1125. [Google Scholar] [CrossRef]
- Talukder, S.K.; Babar, M.A.; Vijayalakshmi, K.; Poland, J.; Prasad, P.V.; Bowden, R.; Fritz, A. Mapping QTL for the traits associated with heat tolerance in wheat (Triticum aestivum L.). BMC Genet. 2014, 15, 97. [Google Scholar] [CrossRef] [Green Version]
- Blum, A.; Sinmena, B.; Mayer, J.; Golan, G.; Shpiler, L. Stem Reserve Mobilization Supports Wheat-Grain Filling under Heat-Stress. Aust. J. Plant Physiol. 1994, 21, 771–781. [Google Scholar] [CrossRef]
- Pinto, R.S.; Reynolds, M.P.; Mathews, K.L.; McIntyre, C.L.; Olivares-Villegas, J.J.; Chapman, S.C. Heat and drought adaptive QTL in a wheat population designed to minimize confounding agronomic effects. Theor. Appl. Genet. 2010, 121, 1001–1021. [Google Scholar] [CrossRef] [Green Version]
- Pinto, R.S.; Reynolds, M.P. Common genetic basis for canopy temperature depression under heat and drought stress associated with optimized root distribution in bread wheat. Theor. Appl. Genet. 2015, 128, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Kumari, M.; Pudake, R.N.; Singh, V.P.; Joshi, A.K. Association of staygreen trait with canopy temperature depression and yield traits under terminal heat stress in wheat (Triticum aestivum L.). Euphytica 2013, 190, 87–97. [Google Scholar] [CrossRef]
- Pinto, R.S.; Lopes, M.S.; Collins, N.C.; Reynolds, M.P. Modelling and genetic dissection of staygreen under heat stress. Theor. Appl. Genet. 2016, 129, 2055–2074. [Google Scholar] [CrossRef] [Green Version]
- Asseng, S.; Ewert, F.; Martre, P.; Rotter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising temperatures reduce global wheat production. Nat. Clim. Chang. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Kumar, S.; Kumari, P.; Kumar, U.; Grover, M.; Singh, A.K.; Singh, R.; Sengar, R.S. Molecular approaches for designing heat tolerant wheat. J. Plant Biochem. Biot. 2013, 22, 359–371. [Google Scholar] [CrossRef]
- Abdolshahi, R.; Nazari, M.; Safarian, A.; Sadathossini, T.; Salarpour, M.; Amiri, H. Integrated selection criteria for drought tolerance in wheat (Triticum aestivum L.) breeding programs using discriminant analysis. Field Crop Res. 2015, 174, 20–29. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Alderfasi, A.; El-Hendawy, S.; Al-Suhaibani, N.; El-Kafafi, S.; Seleiman, M.F. Detecting Salt Tolerance in Doubled Haploid Wheat Lines. Agronomy 2019, 9, 211. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, K.; Mondal, S.; Ray, S.; Samal, P.; Pradhan, B.; Chattopadhyay, K.; Kar, M.K.; Swain, P.; Sarkar, R.K. Tissue tolerance coupled with ionic discrimination can potentially minimize the energy cost of salinity tolerance in rice. Front. Plant Sci. 2020, 11, 265. [Google Scholar] [CrossRef]
- Dadshani, S.; Sharma, R.C.; Baum, M.; Ogbonnaya, F.C.; Léon, J.; Ballvora, A. Multi-dimensional evaluation of response to salt stress in wheat. PLoS ONE 2019, 14, e0222659. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Romdhane, W.B.; El-Said, R.A.; Ghazy, A.; Attia, K.; Al-Doss, A. Agro-Physiologic Responses and Stress-Related Gene Expression of Four Doubled Haploid Wheat Lines under Salinity Stress Conditions. Biology 2021, 10, 56. [Google Scholar] [CrossRef]
- El-Hendawy, S.; Al-Suhaibani, N.; Al-Ashkar, I.; Alotaibi, M.; Tahir, M.U.; Solieman, T.; Hassan, W.M. Combining Genetic Analysis and Multivariate Modeling to Evaluate Spectral Reflectance Indices as Indirect Selection Tools in Wheat Breeding under Water Deficit Stress Conditions. Remote Sens. 2020, 12, 1480. [Google Scholar] [CrossRef]
- Grzesiak, S.; Hordyńska, N.; Szczyrek, P.; Grzesiak, M.T.; Noga, A.; Szechyńska-Hebda, M. Variation among wheat (Triticum easativum L.) genotypes in response to the drought stress: I–selection approaches. J. Plant Interact. 2019, 14, 30–44. [Google Scholar] [CrossRef] [Green Version]
- De Leon, T.B.; Linscombe, S.; Gregorio, G.; Subudhi, P.K. Genetic variation in Southern USA rice genotypes for seedling salinity tolerance. Front. Plant Sci. 2015, 6, 374. [Google Scholar] [CrossRef] [Green Version]
- El-Hennawy, M.; Abdalla, A.; Shafey, S.A.; Al-Ashkar, I. Production of doubled haploid wheat lines (Triticum aestivum L.) using anther culture technique. Ann. Agric. Sci. 2011, 56, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.M.; McCaig, T.N. Excised-leaf water retention capability as an indicator of drought resistance of Triticum genotypes. Can. J. Plant Sci. 1982, 62, 571–578. [Google Scholar] [CrossRef]
- Barrs, H. Determination of water deficits in plant tissues. In Water Deficits and Plant Growth; Academic Press: New York, NY, USA, 1968; Volume 1, pp. 235–368. [Google Scholar]
- Wang, X.; Wang, L.; Shangguan, Z. Leaf gas exchange and fluorescence of two winter wheat varieties in response to drought stress and nitrogen supply. PLoS ONE 2016, 11, e0165733. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of Catalases and Peroxidases. Method Enzymol. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Duckworth, H.W.; Coleman, J.E. Physicochemical and kinetic properties of mushroom tyrosinase. J. Biol. Chem. 1970, 245, 1613–1625. [Google Scholar] [CrossRef]
- Fehr, W. Principles of Cultivar Development: Theory and Technique; Macmillian Publishing Company: New York, NY, USA, 1991. [Google Scholar]
- Zadeh, L.A. Fuzzy sets as a basis for a theory of possibility. J. Fuzzy Sets Syst. 1978, 1, 3–28. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Alderfasi, A.; Ben Romdhane, W.; Seleiman, M.F.; El-Said, R.A.; Al-Doss, A. Morphological and genetic diversity within salt tolerance detection in eighteen wheat genotypes. Plants 2020, 9, 287. [Google Scholar] [CrossRef] [Green Version]
- Fischer, K. Breeding Rice for Drought-Prone Environments; International Rice Research Institute: Los Baños, Philippines, 2003; Volume 1. [Google Scholar]
- Bustos-Korts, D.; Boer, M.P.; Malosetti, M.; Chapman, S.; Chenu, K.; Zheng, B.; van Eeuwijk, F.A. Combining Crop Growth Modeling and Statistical Genetic Modeling to Evaluate Phenotyping Strategies. Front. Plant Sci. 2019, 10, 1491. [Google Scholar] [CrossRef] [Green Version]
- Falconer, D.; Mackay, T.; Bulmer, M. Introduction to Quantitative Genetics. Genet. Res. 1996, 68, 183. [Google Scholar]
- Obala, J.; Saxena, R.; Singh, V.; Vechalapu, S.; Das, R.R.; Rathore, A.; Kumar, C.S.; Saxena, K.; Tongoona, P.; Sibiya, J.; et al. Genetic variation and relationships of total seed protein content with some agronomic traits in pigeonpea (Cajanus cajan (L.) Millsp.). Aust. J. Crop Sci. 2018, 12, 1859–1865. [Google Scholar] [CrossRef]
- Barbin, D. Planejamento e Análise de Experimentos Agronômicos; Midas: Arapongas, Brazil, 2003. [Google Scholar]
- Wartha, C.A.; Cargnelutti Filho, A.; Lucio, A.D.; Follmann, D.N.; Kleinpaul, J.A.; Simoes, F.M. Sample sizes to estimate mean values for tassel traits in maize genotypes. Genet. Mol. Res. 2016, 15, gmr15049151. [Google Scholar] [CrossRef]
- Yu, R.; Wang, G.; Yu, X.; Li, L.; Li, C.; Song, Y.; Xu, Z.; Zhang, J.; Guan, C. Assessing alfalfa (Medicago sativa L.) tolerance to salinity at seedling stage and screening of the salinity tolerance traits. Plant Biol. 2021, 23, 664–674. [Google Scholar] [CrossRef]
- El-Hendawy, S.; Elshafei, A.; Al-Suhaibani, N.; Alotabi, M.; Hassan, W.; Dewir, Y.H.; Abdella, K. Assessment of the salt tolerance of wheat genotypes during the germination stage based on germination ability parameters and associated SSR markers. J. Plant Interact. 2019, 14, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, D.; Cornacchione, M.V.; Ferreira, J.F.; Suarez, D.L. Variable salinity responses of 12 alfalfa genotypes and comparative expression analyses of salt-response genes. Sci. Rep. 2017, 7, 42958. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Ma, X.; Cui, D.; Wang, Y.; Geng, L.; Cao, G.; Zhang, H.; Han, L. Comprehensive Evaluation and Analysis of the Mechanism of Cold Tolerance Based on the Transcriptome of Weedy Rice Seedlings. Rice 2020, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Crop breeding for salt tolerance in the era of molecular markers and marker-assisted selection. Plant Breed. 2013, 132, 10–20. [Google Scholar] [CrossRef]
- Khan, A.S.; Ashfaq, M.; Asad, M.A. A correlation and path coefficient analysis for some yield components in bread wheat. Asian J. Plant Sci. 2003, 2, 582–584. [Google Scholar]
- Del Moral, L.; Rharrabti, Y.; Villegas, D.; Royo, C. Evaluation of grain yield and its components in durum wheat under Mediterranean conditions. Agron. J. 2003, 95, 266–274. [Google Scholar] [CrossRef]
- Bojarian, M.; Asadi-Gharneh, H.A.; Golabadi, M. Factor analysis, stepwise regression and path coefficient analyses of yield, yield-associated traits, and fruit quality in tomato. Int. J. Veg. Sci. 2019, 25, 542–553. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Rattey, A.R.; Farquhar, G.D.; Richards, R.A.; Condon, A.T.G. Genomic regions for canopy temperature and their genetic association with stomatal conductance and grain yield in wheat. Funct. Plant Biol. 2013, 40, 14–33. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Pierre, C.S.; Saad, A.S.; Vargas, M.; Condon, A.G. Evaluating potential genetic gains in wheat associated with stress-adaptive trait expression in elite genetic resources under drought and heat stress. Crop Sci. 2007, 47, S-172–S-189. [Google Scholar] [CrossRef]
- Gautam, A.; Prasad, S.S.; Jajoo, A.; Ambati, D. Canopy temperature as a selection parameter for grain yield and its components in durum wheat under terminal heat stress in late sown conditions. Agric. Res. 2015, 4, 238–244. [Google Scholar] [CrossRef]
- Bahar, B.; Yildirim, M.; Barutcular, C.; Ibrahim, G. Effect of canopy temperature depression on grain yield and yield components in bread and durum wheat. Not. Bot. Horti Agrobot. 2008, 36, 34–37. [Google Scholar]
- Mason, R.E.; Singh, R.P. Considerations when deploying canopy temperature to select high yielding wheat breeding lines under drought and heat stress. Agronomy 2014, 4, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, M.; Manes, Y.; Izanloo, A.; Langridge, P. Phenotyping approaches for physiological breeding and gene discovery in wheat. Ann. Appl. Biol. 2009, 155, 309–320. [Google Scholar] [CrossRef]
- Reynolds, M.; Pask, A.; Mullan, D. Physiological Breeding I: Interdisciplinary Approaches to Improve Crop Adaptation; CIMMYT: Mexico City, Mexico, 2012. [Google Scholar]
- Zeng, L.; Shannon, M.C.; Grieve, C.M. Evaluation of salt tolerance in rice genotypes by multiple agronomic parameters. Euphytica 2002, 127, 235–245. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Al-Suhaibani, N.; Abdella, K.; Sallam, M.; Alotaibi, M.; Seleiman, M.F. Combining Genetic and Multidimensional Analyses to Identify Interpretive Traits Related to Water Shortage Tolerance as an Indirect Selection Tool for Detecting Genotypes of Drought Tolerance in Wheat Breeding. Plants 2021, 10, 931. [Google Scholar] [CrossRef] [PubMed]
- Platten, J.D.; Egdane, J.A.; Ismail, A.M. Salinity tolerance, Na+ exclusion and allele mining of HKT1;5 in Oryza sativa and O. glaberrima: Many sources, many genes, one mechanism? BMC Plant Biol. 2013, 13, 32. [Google Scholar] [CrossRef]
- Truxillo, C. Multivariate Statistical Methods: Practical Research Applications: Course Notes; SAS Institute: Cary, NC, USA, 2003; ISBN 159994586X. [Google Scholar]
- Singamsetti, A.; Shahi, J.P.; Zaidi, P.H.; Seetharam, K.; Vinayan, M.T.; Kumar, M.; Singla, S.; Shikha, K.; Madankar, K. Genotype × environment interaction and selection of maize (Zea mays L.) hybrids across moisture regimes. Field Crops Res. 2021, 270, 108224. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D.; da Silva, J.A.; Sari, B.G.; Diel, M.I. Mean performance and stability in multi-environment trials II: Selection based on multiple traits. Agron. J. 2019, 111, 2961–2969. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D.; da Silva, J.A.; Marchioro, V.S.; de Souza, V.Q.; Jost, E. Mean performance and stability in multi-environment trials I: Combining features of AMMI and BLUP techniques. Agron. J. 2019, 111, 2949–2960. [Google Scholar] [CrossRef]
- Olivoto, T.; Nardino, M. MGIDI: Toward an effective multivariate selection in biological experiments. Bioinformatics 2021, 37, 1383–1389. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variations | df | CT | LWC | RWC | Pn | Gs | Ci | E | Ls | POD | PPO | CAT | GLN | FLA | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season 1 | Replications | 2 | 0.0023 | 0.0018 | 0.0017 | 0.0018 | 0.0012 | 0.0012 | 0.0013 | 0.0031 | 0.0030 | 0.0014 | 0.0028 | 0.0015 | 0.0016 |
| Genotypes (G) | 19 | 0.0104 | 0.0135 | 0.0092 | 0.0307 | 0.0797 | 0.0171 | 0.0449 | 0.4274 | 1.6312 | 0.5980 | 1.9533 | 0.0154 | 0.0103 | |
| Error | 38 | 0.0015 | 0.0014 | 0.0013 | 0.0013 | 0.0008 | 0.0010 | 0.0009 | 0.0025 | 0.0019 | 0.0012 | 0.0023 | 0.0013 | 0.0012 | |
| Season 2 | Replications | 2 | 0.0028 | 0.0017 | 0.0017 | 0.0017 | 0.0014 | 0.0013 | 0.0013 | 0.0031 | 0.0029 | 0.0014 | 0.0027 | 0.0017 | 0.0014 |
| Genotypes (G) | 19 | 0.0346 | 0.0080 | 0.0099 | 0.0424 | 0.1201 | 0.0212 | 0.0689 | 0.2249 | 1.6030 | 0.5882 | 1.9203 | 0.0445 | 0.0368 | |
| Error | 38 | 0.0026 | 0.0013 | 0.0012 | 0.0014 | 0.0015 | 0.0010 | 0.0010 | 0.0023 | 0.0019 | 0.0012 | 0.0023 | 0.0014 | 0.0011 | |
| Combined | Seasons (S) | 1 | 0.1535 | 0.0533 | 0.0319 | 0.0218 | 0.0355 | 0.0041 | 0.0028 | 0.0490 | 0.0012 | 0.0010 | 0.0010 | 0.0908 | 0.0618 |
| Replications (Sea.) | 4 | 0.0026 | 0.0018 | 0.0017 | 0.0018 | 0.0013 | 0.0013 | 0.0013 | 0.0031 | 0.0029 | 0.0014 | 0.0028 | 0.0016 | 0.0015 | |
| Genotypes (G) | 19 | 0.0368 | 0.0178 | 0.0140 | 0.0660 | 0.1915 | 0.0363 | 0.1036 | 0.6125 | 0.8210 | 0.9820 | 0.8670 | 0.0340 | 0.0369 | |
| S ×G | 19 | 0.0085 | 0.0041 | 0.0052 | 0.0072 | 0.0080 | 0.0018 | 0.0100 | 0.0410 | 0.0002 | 0.0001 | 0.0001 | 0.0261 | 0.0102 | |
| Error | 76 | 0.0020 | 0.0014 | 0.0013 | 0.0013 | 0.0012 | 0.0010 | 0.0009 | 0.0024 | 0.0019 | 0.0012 | 0.0023 | 0.0013 | 0.0012 | |
| Heritability (h2 %) | 64.53 | 60.43 | 60.61 | 80.10 | 89.51 | 77.94 | 84.43 | 86.66 | 62.05 | 83.66 | 85.61 | 27.50 | 61.88 | ||
| Genetic gain (GG %) | 14.95 | 10.95 | 10.87 | 22.05 | 43.21 | 17.22 | 31.12 | 54.25 | 50.44 | 37.60 | 44.10 | 5.76 | 13.78 | ||
| G.C.V. % | 9.03 | 6.83 | 6.78 | 11.96 | 22.17 | 9.47 | 16.44 | 28.29 | 31.08 | 19.96 | 23.14 | 5.33 | 8.50 | ||
| Ph.C.V. % | 11.24 | 8.79 | 8.71 | 13.36 | 23.43 | 10.73 | 17.89 | 30.39 | 39.46 | 21.82 | 25.00 | 10.17 | 10.81 | ||
| Source of variations | df | GLA | LAI | DH | MD | GFD | NS | PH | SL | NSS | NKS | HW | GY | ||
| Season 1 | Replications | 2 | 0.0016 | 0.0005 | 0.0018 | 0.0017 | 0.0016 | 0.0012 | 0.0015 | 0.0015 | 0.0013 | 0.0016 | 0.0013 | 0.0012 | |
| Genotypes (G) | 19 | 0.0283 | 0.0441 | 0.0083 | 0.0062 | 0.0101 | 0.0299 | 0.0062 | 0.0240 | 0.0227 | 0.0139 | 0.0070 | 0.0260 | ||
| Error | 38 | 0.0013 | 0.0004 | 0.0022 | 0.0013 | 0.0017 | 0.0009 | 0.0013 | 0.0013 | 0.0012 | 0.0012 | 0.0010 | 0.0013 | ||
| Season 2 | Replications | 2 | 0.0011 | 0.0008 | 0.0017 | 0.0015 | 0.0013 | 0.0011 | 0.0016 | 0.0034 | 0.0018 | 0.0016 | 0.0014 | 0.0008 | |
| Genotypes (G) | 19 | 0.0593 | 0.0381 | 0.0081 | 0.0113 | 0.0438 | 0.0311 | 0.0177 | 0.1135 | 0.0135 | 0.0231 | 0.0090 | 0.0834 | ||
| Error | 38 | 0.0010 | 0.0007 | 0.0014 | 0.0012 | 0.0010 | 0.0008 | 0.0013 | 0.0023 | 0.0012 | 0.0014 | 0.0011 | 0.0008 | ||
| Combined | Seasons (S) | 1 | 0.4502 | 0.1920 | 0.0001 | 0.0423 | 0.002 | 0.0214 | 0.0030 | 0.5210 | 0.0337 | 0.0636 | 0.0183 | 0.0560 | |
| Replications (Sea.) | 4 | 0.0013 | 0.0007 | 0.0017 | 0.0016 | 0.0014 | 0.0011 | 0.0016 | 0.0024 | 0.0015 | 0.0016 | 0.0013 | 0.0010 | ||
| Genotypes (G) | 19 | 0.0582 | 0.0520 | 0.0154 | 0.0164 | 0.046 | 0.0414 | 0.0139 | 0.0501 | 0.0182 | 0.0244 | 0.0112 | 0.0816 | ||
| S ×G | 19 | 0.0293 | 0.0304 | 0.0009 | 0.0001 | 0.0070 | 0.0196 | 0.0100 | 0.0873 | 0.0183 | 0.0127 | 0.0047 | 0.0281 | ||
| Error | 76 | 0.0011 | 0.0005 | 0.0018 | 0.0013 | 0.0013 | 0.0009 | 0.0013 | 0.0018 | 0.0012 | 0.0013 | 0.0011 | 0.0011 | ||
| Heritability (h2 %) | 49.07 | 38.65 | 55.22 | 60.47 | 69.29 | 47.44 | 45.30 | 69.45 | 19.34 | 33.73 | 56.69 | 80.73 | |||
| Genetic gain (GG %) | 13.67 | 15.06 | 10.15 | 10.35 | 20.05 | 12.20 | 6.13 | 14.75 | 2.40 | 5.85 | 10.36 | 20.78 | |||
| G.C.V. % | 9.47 | 11.76 | 6.63 | 6.46 | 11.69 | 8.59 | 4.42 | 8.59 | 2.65 | 4.89 | 6.68 | 11.23 | |||
| Ph.C.V. % | 13.52 | 18.91 | 8.92 | 8.31 | 14.05 | 12.48 | 6.57 | 10.31 | 6.03 | 8.42 | 8.87 | 12.50 | |||
| Indices | S1 | S2 | Combined Data | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean ± SD | Min | Max | Mean ± SD | Min | Max | Mean ± SD | CV | |
| CT | 0.998 | 1.081 | 0.989 ± 0.059 | 0.950 | 1.131 | 0.988 ± 0.107 | 0.980 | 1.213 | 0.972 ± 0.045 | 4.60 |
| LWC | 0.804 | 0.974 | 0.955 ± 0.067 | 0.831 | 0.910 | 0.891 ± 0.051 | 0.826 | 0.962 | 0.913 ± 0.037 | 4.10 |
| RWC | 0.807 | 0.950 | 0.922 ± 0.055 | 0.769 | 0.891 | 0.889 ± 0.057 | 0.829 | 0.935 | 0.906 ± 0.036 | 3.98 |
| Pn | 0.700 | 0.987 | 0.895 ± 0.101 | 0.741 | 0.996 | 0.921 ± 0.118 | 0.721 | 1.050 | 0.908 ± 0.036 | 3.97 |
| Gs | 0.377 | 0.871 | 0.704 ± 0.163 | 0.386 | 0.881 | 0.738 ± 0.200 | 0.382 | 0.995 | 0.722 ± 0.035 | 4.80 |
| Ci | 0.657 | 0.917 | 0.795 ± 0.076 | 0.654 | 0.904 | 0.808 ± 0.084 | 0.656 | 0.877 | 0.802 ± 0.032 | 3.94 |
| E | 0.465 | 0.830 | 0.754 ± 0.122 | 0.433 | 0.850 | 0.764 ± 0.151 | 0.449 | 0.946 | 0.760 ± 0.030 | 3.95 |
| Ls | 0.745 | 0.981 | 0.932 ± 0.377 | 0.745 | 0.970 | 0.901 ± 0.273 | 0.745 | 0.959 | 0.903 ± 0.049 | 5.43 |
| POD | 0.072 | 1.146 | 0.832 ± 0.737 | 0.071 | 1.145 | 0.825 ± 0.731 | 0.072 | 1.154 | 0.829 ± 0.044 | 5.26 |
| PPO | 0.085 | 1.164 | 0.768 ± 0.446 | 0.084 | 1.162 | 0.762 ± 0.442 | 0.085 | 1.068 | 0.865 ± 0.035 | 4.00 |
| CAT | 0.103 | 1.153 | 0.915 ± 0.807 | 0.102 | 1.197 | 0.909 ± 0.800 | 0.103 | 1.627 | 0.912 ± 0.048 | 5.26 |
| GLN | 0.693 | 0.920 | 0.889 ± 0.072 | 0.705 | 0.930 | 0.904 ± 0.121 | 0.760 | 0.994 | 0.902 ± 0.036 | 4.00 |
| FLA | 0.687 | 0.928 | 0.884 ± 0.059 | 0.615 | 0.918 | 0.838 ± 0.110 | 0.703 | 0.920 | 0.861 ± 0.035 | 4.02 |
| GLA | 0.612 | 0.991 | 0.888 ± 0.097 | 0.449 | 0.970 | 0.796 ± 0.140 | 0.610 | 0.913 | 0.826 ± 0.033 | 4.02 |
| LAI | 0.329 | 0.712 | 0.471 ± 0.121 | 0.328 | 0.661 | 0.551 ± 0.112 | 0.345 | 0.671 | 0.511 ± 0.022 | 4.37 |
| DH | 0.838 | 0.948 | 0.835 ± 0.053 | 0.858 | 0.945 | 0.934 ± 0.052 | 0.848 | 0.978 | 0.870 ± 0.042 | 4.87 |
| MD | 0.848 | 0.911 | 0.812 ± 0.045 | 0.787 | 0.933 | 0.837 ± 0.061 | 0.817 | 0.952 | 0.845 ± 0.036 | 4.27 |
| GFD | 0.693 | 0.926 | 0.779 ± 0.058 | 0.575 | 0.958 | 0.769 ± 0.120 | 0.694 | 0.983 | 0.712 ± 0.036 | 5.06 |
| NS | 0.583 | 0.794 | 0.752 ± 0.100 | 0.559 | 0.809 | 0.726 ± 0.101 | 0.596 | 0.813 | 0.739 ± 0.030 | 4.06 |
| PH | 0.793 | 0.873 | 0.855 ± 0.046 | 0.736 | 0.854 | 0.804 ± 0.076 | 0.824 | 0.928 | 0.889 ± 0.036 | 4.05 |
| SL | 0.751 | 0.959 | 0.911 ± 0.089 | 0.770 | 0.947 | 0.907 ± 0.194 | 0.803 | 0.959 | 0.912 ± 0.042 | 4.65 |
| NSS | 0.664 | 0.892 | 0.851 ± 0.087 | 0.754 | 0.913 | 0.884 ± 0.067 | 0.760 | 0.881 | 0.857 ± 0.035 | 4.04 |
| NKS | 0.764 | 0.979 | 0.883 ± 0.068 | 0.756 | 0.980 | 0.927 ± 0.087 | 0.760 | 0.981 | 0.905 ± 0.036 | 3.98 |
| HW | 0.711 | 0.817 | 0.803 ± 0.048 | 0.720 | 0.845 | 0.828 ± 0.054 | 0.742 | 0.852 | 0.816 ± 0.033 | 4.06 |
| GY | 0.444 | 0.894 | 0.776 ± 0.092 | 0.436 | 0.851 | 0.704 ± 0.166 | 0.440 | 0.899 | 0.740 ± 0.033 | 4.48 |
| PCI1 | PCI2 | PCI3 | PCI4 | PCI5 | PCI6 | PCI7 | |
|---|---|---|---|---|---|---|---|
| Eigenvalue | 9.221 | 4.157 | 2.829 | 2.053 | 1.352 | 1.138 | 1.114 |
| Variability (%) | 35.465 | 15.989 | 10.883 | 7.896 | 5.199 | 4.378 | 4.283 |
| Cumulative % | 35.465 | 51.454 | 62.336 | 70.232 | 75.431 | 79.809 | 84.092 |
| Eigenvectors: | |||||||
| CT | −0.210 | 0.200 | 0.162 | 0.158 | 0.056 | 0.012 | −0.305 |
| LWC | 0.138 | 0.171 | 0.217 | 0.355 | 0.177 | 0.101 | 0.237 |
| RWC | 0.310 | 0.060 | 0.001 | 0.016 | 0.095 | −0.169 | −0.070 |
| Pn | 0.128 | −0.195 | −0.320 | 0.274 | −0.038 | 0.148 | 0.114 |
| Gs | −0.040 | 0.371 | −0.235 | −0.111 | −0.277 | −0.125 | 0.083 |
| Ci | −0.028 | 0.427 | −0.151 | 0.023 | 0.214 | 0.075 | −0.090 |
| E | −0.012 | 0.390 | −0.225 | −0.179 | −0.160 | −0.187 | 0.134 |
| Ls | 0.248 | −0.040 | 0.165 | −0.102 | 0.108 | −0.071 | 0.033 |
| POD | 0.102 | 0.070 | 0.085 | −0.314 | −0.391 | 0.517 | 0.132 |
| PPO | 0.181 | 0.086 | −0.236 | −0.327 | 0.024 | −0.122 | 0.324 |
| CAT | 0.225 | −0.070 | −0.256 | 0.294 | −0.089 | 0.167 | 0.014 |
| GLN | 0.246 | 0.053 | −0.019 | −0.311 | 0.174 | 0.150 | −0.318 |
| FLA | 0.233 | 0.175 | 0.131 | 0.065 | 0.065 | 0.171 | 0.278 |
| GLA | 0.145 | 0.098 | −0.169 | −0.001 | 0.559 | 0.371 | 0.143 |
| LAI | 0.193 | −0.081 | 0.290 | −0.096 | −0.244 | 0.151 | 0.009 |
| DH | 0.239 | 0.061 | 0.145 | −0.136 | 0.028 | −0.272 | −0.352 |
| MD | 0.288 | 0.059 | 0.131 | 0.065 | 0.093 | −0.079 | −0.138 |
| GFD | 0.186 | −0.192 | −0.261 | 0.168 | 0.031 | −0.194 | −0.138 |
| NS | 0.233 | 0.116 | 0.077 | −0.011 | −0.084 | 0.157 | −0.270 |
| PH | 0.276 | −0.044 | −0.133 | −0.127 | 0.082 | −0.119 | −0.034 |
| SL | 0.225 | 0.155 | 0.171 | 0.044 | −0.160 | 0.091 | −0.111 |
| NSS | 0.055 | 0.010 | 0.392 | −0.039 | 0.125 | −0.273 | 0.442 |
| NKS | 0.056 | 0.228 | 0.158 | 0.438 | −0.266 | 0.035 | −0.026 |
| HW | 0.257 | 0.100 | −0.035 | 0.168 | −0.155 | −0.337 | 0.149 |
| GY | 0.235 | −0.142 | −0.237 | 0.108 | −0.254 | 0.033 | −0.023 |
| Genotypes | The Values of Comprehensive Index (PCIi) | Membership Function Value | D | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PCI1 | PCI2 | PCI3 | PCI4 | PCI5 | PCI6 | PCI7 | P μ(x1) | P μ(x2) | P μ(x3) | P μ(x4) | P μ(x5) | P μ(x6) | P μ(x7) | Value | |
| DHL12 | −0.449 | 1.653 | 0.442 | 1.106 | 0.901 | 0.023 | −0.370 | 0.508 | 0.629 | 0.694 | 0.708 | 0.754 | 0.318 | 0.381 | 0.510 |
| DHL02 | −0.474 | 4.236 | 0.920 | 2.481 | −1.287 | −0.491 | 1.226 | 0.506 | 1.000 | 0.780 | 1.000 | 0.256 | 0.195 | 0.766 | 0.638 |
| DHL25 | −0.100 | −1.192 | 0.085 | 0.607 | 0.077 | −0.143 | 2.194 | 0.538 | 0.220 | 0.630 | 0.602 | 0.567 | 0.278 | 1.000 | 0.458 |
| DHL07 | −0.455 | 0.579 | −0.132 | −0.576 | −0.699 | −0.814 | 1.068 | 0.508 | 0.475 | 0.591 | 0.350 | 0.390 | 0.117 | 0.728 | 0.451 |
| DHL26 | −4.260 | −2.042 | −1.307 | −2.225 | −0.509 | −0.405 | 0.760 | 0.178 | 0.098 | 0.379 | 0.000 | 0.433 | 0.215 | 0.654 | 0.176 |
| Gemmeiza-9 | −1.151 | −0.275 | 0.927 | 1.222 | 1.670 | 0.183 | 0.635 | 0.447 | 0.352 | 0.782 | 0.733 | 0.930 | 0.356 | 0.623 | 0.458 |
| DHL11 | −3.398 | 0.644 | 2.012 | 0.148 | −1.776 | −0.577 | −1.813 | 0.253 | 0.484 | 0.977 | 0.504 | 0.145 | 0.174 | 0.032 | 0.374 |
| KSU106 | −3.055 | −2.727 | 0.150 | 1.117 | 1.979 | 2.086 | −0.195 | 0.282 | 0.000 | 0.642 | 0.710 | 1.000 | 0.811 | 0.423 | 0.293 |
| Gemmeiza-12 | −3.858 | 1.622 | 2.139 | −1.332 | 1.873 | −0.452 | −0.286 | 0.213 | 0.625 | 1.000 | 0.190 | 0.976 | 0.204 | 0.401 | 0.377 |
| DHL01 | −6.311 | −0.143 | −2.197 | 1.552 | −0.591 | −0.665 | −0.435 | 0.000 | 0.371 | 0.219 | 0.803 | 0.415 | 0.153 | 0.365 | 0.193 |
| DHL14 | −0.906 | −0.395 | −0.419 | −1.227 | −2.410 | 2.878 | 0.532 | 0.468 | 0.335 | 0.539 | 0.212 | 0.000 | 1.000 | 0.598 | 0.381 |
| DHL29 | 2.112 | 2.032 | −2.439 | −1.936 | 1.441 | −0.849 | 0.989 | 0.730 | 0.683 | 0.175 | 0.062 | 0.877 | 0.109 | 0.709 | 0.503 |
| DHL15 | 2.674 | 0.093 | −3.411 | 0.491 | −0.425 | −0.787 | −0.282 | 0.779 | 0.405 | 0.000 | 0.577 | 0.452 | 0.124 | 0.402 | 0.480 |
| DHL06 | 1.781 | −1.032 | −2.367 | −0.815 | 0.173 | 0.159 | −1.258 | 0.701 | 0.244 | 0.188 | 0.300 | 0.589 | 0.350 | 0.166 | 0.404 |
| Misr1 | 1.812 | −2.579 | 1.851 | −0.721 | −0.101 | −0.588 | −0.118 | 0.704 | 0.021 | 0.948 | 0.320 | 0.526 | 0.171 | 0.441 | 0.476 |
| DHL05 | 2.589 | 2.069 | −1.560 | 1.179 | 0.565 | 1.023 | −1.944 | 0.771 | 0.689 | 0.333 | 0.723 | 0.678 | 0.557 | 0.000 | 0.569 |
| DHL23 | 3.730 | 3.361 | 1.646 | −1.906 | 0.079 | 1.464 | 0.305 | 0.870 | 0.874 | 0.911 | 0.068 | 0.567 | 0.662 | 0.544 | 0.686 |
| Sakha-93 | 1.116 | −0.655 | 1.634 | −1.723 | −0.290 | −1.305 | −1.433 | 0.644 | 0.298 | 0.909 | 0.107 | 0.483 | 0.000 | 0.124 | 0.462 |
| Pavone-76 | 3.377 | −2.693 | 1.080 | 0.542 | −0.323 | −0.456 | 0.345 | 0.840 | 0.005 | 0.809 | 0.588 | 0.476 | 0.203 | 0.553 | 0.543 |
| DHL08 | 5.228 | −2.556 | 0.946 | 2.017 | −0.346 | −0.282 | 0.080 | 1.000 | 0.025 | 0.785 | 0.901 | 0.470 | 0.245 | 0.489 | 0.638 |
| Source | Stepwise Regression | Path Coefficient | ||||||
|---|---|---|---|---|---|---|---|---|
| Partitioning the Correlation | R2 | |||||||
| Regression Coefficient | p-Value | R2 Par. | R2 Com. | Direct Effect | Indirect Effect | Correlation Value | Direct Effect | |
| Intercept | 0.670 | 0.033 | ||||||
| GFD | 0.504 | 0.007 | 0.529 | 0.529 | 0.460 | 0.345 | 0.805 | 0.211 |
| SL | 0.334 | 0.049 | 0.074 | 0.603 | 0.276 | 0.222 | 0.498 | 0.076 |
| CT | −0.466 | 0.009 | 0.136 | 0.739 | −0.437 | −0.241 | −0.677 | 0.191 |
| Indirect effect | 0.261 | |||||||
| Total R2 | 0.739 | 0.739 | ||||||
| Residual | 0.511 | 0.511 | ||||||
| Genotypes | Dependent Indices | GY | Regression “GY” Value | Predicted Error Value | Relative Error Value | Evaluation Accuracy (%) | ||
|---|---|---|---|---|---|---|---|---|
| GFD | SL | CT | ||||||
| DHL12 | 0.694 | 0.845 | 1.315 | 0.769 | 0.689 | 0.081 | 0.105 | 89.49 |
| DHL02 | 0.695 | 0.896 | 1.286 | 0.736 | 0.720 | 0.016 | 0.021 | 97.86 |
| DHL25 | 0.798 | 0.907 | 1.078 | 0.840 | 0.873 | −0.032 | −0.038 | 96.16 |
| DHL07 | 0.783 | 0.751 | 1.007 | 0.811 | 0.846 | −0.035 | −0.043 | 95.72 |
| DHL26 | 0.694 | 0.709 | 1.171 | 0.773 | 0.711 | 0.062 | 0.080 | 92.03 |
| Gemmeiza-9 | 0.758 | 0.752 | 1.155 | 0.775 | 0.764 | 0.011 | 0.014 | 98.57 |
| DHL11 | 0.606 | 0.870 | 1.293 | 0.648 | 0.663 | −0.015 | −0.023 | 97.73 |
| KSU106 | 0.667 | 0.707 | 1.199 | 0.662 | 0.683 | −0.021 | −0.032 | 96.85 |
| Gemmeiza-12 | 0.709 | 0.794 | 1.300 | 0.545 | 0.687 | −0.142 | −0.261 | 73.92 |
| DHL01 | 0.725 | 0.640 | 1.250 | 0.729 | 0.666 | 0.063 | 0.086 | 91.39 |
| DHL14 | 0.751 | 0.863 | 1.137 | 0.846 | 0.807 | 0.039 | 0.046 | 95.36 |
| DHL29 | 0.783 | 0.805 | 1.083 | 0.736 | 0.829 | −0.093 | −0.126 | 87.37 |
| DHL15 | 0.975 | 0.859 | 1.086 | 0.929 | 0.942 | −0.013 | −0.014 | 98.57 |
| DHL06 | 0.817 | 0.851 | 1.029 | 0.919 | 0.886 | 0.033 | 0.036 | 96.41 |
| Misr1 | 0.783 | 0.850 | 1.082 | 0.820 | 0.844 | −0.024 | −0.029 | 97.08 |
| DHL05 | 0.852 | 0.929 | 1.198 | 0.920 | 0.851 | 0.069 | 0.075 | 92.53 |
| DHL23 | 0.715 | 0.857 | 1.099 | 0.777 | 0.805 | −0.028 | −0.036 | 96.40 |
| Sakha-93 | 0.757 | 0.863 | 1.185 | 0.814 | 0.787 | 0.027 | 0.033 | 96.69 |
| Pavone-76 | 0.898 | 0.903 | 1.090 | 0.869 | 0.916 | −0.047 | −0.054 | 94.55 |
| DHL08 | 0.929 | 0.929 | 1.022 | 0.951 | 0.972 | −0.022 | −0.023 | 97.73 |
| Average | 94.12 | |||||||
| Statistic | GFD | SL | CT | GY |
|---|---|---|---|---|
| Multicollinearity statistics: | ||||
| Tolerance | 0.321 | 0.696 | 0.465 | 0.278 |
| VIF | 3.114 | 1.436 | 2.151 | 3.598 |
| Unidimensional test of equality of the means of the classes: | ||||
| Lambda | 0.377 | 0.220 | 0.216 | 0.404 |
| F | 6.195 | 13.319 | 13.584 | 5.536 |
| DF1 | 4 | 4 | 4 | 4 |
| DF2 | 15 | 15 | 15 | 15 |
| p-value | 0.004 | <0.0001 | <0.0001 | 0.006 |
| Parameters | Can1 | Can2 | Can3 |
|---|---|---|---|
| Eigenvalue | 5.551 | 4.433 | 0.200 |
| Discrimination (%) | 54.325 | 43.385 | 1.959 |
| Cumulative % | 54.325 | 97.710 | 99.669 |
| Bartlett’s statistic | 54.926 | 27.670 | 3.128 |
| p-value | 0.000 | 0.001 | 0.537 |
| Canonical correlations | 0.921 | 0.903 | 0.408 |
| Variables/Factors correlations: | |||
| GFD | −0.497 | 0.668 | 0.543 |
| SL | 0.425 | 0.875 | 0.062 |
| CT | 0.732 | −0.629 | 0.181 |
| GY | −0.505 | 0.673 | 0.083 |
| Heat Group | |||
| HS | −1.659 | −3.080 | −0.044 |
| HT | −0.017 | 2.056 | 0.312 |
| I | −2.940 | 0.682 | 0.667 |
| S | 3.681 | −0.949 | 0.173 |
| T | −0.357 | 1.088 | −0.516 |
| Genotypes | Classification | Cross-Validation | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Prior | Posterior | Membership Probabilities | Posterior | Membership Probabilities | |||||||||
| Pr(HS) | Pr(HT) | Pr(I) | Pr(S) | Pr(T) | HS | HT | I | S | T | ||||
| DHL12 | S | S | 0.000 | 0.000 | 0.000 | 0.999 | 0.001 | S | 0.001 | 0.004 | 0.000 | 0.972 | 0.023 |
| DHL02 | S | S | 0.000 | 0.000 | 0.000 | 1.000 | 0.000 | S | 0.000 | 0.000 | 0.000 | 0.999 | 0.001 |
| DHL25 | T | HT | 0.000 | 0.520 | 0.000 | 0.000 | 0.480 | HT | 0.000 | 0.703 | 0.000 | 0.000 | 0.297 |
| DHL07 | I | I | 0.008 | 0.001 | 0.955 | 0.000 | 0.036 | HS | 0.637 | 0.000 | 0.000 | 0.000 | 0.363 |
| DHL26 | HS | HS | 1.000 | 0.000 | 0.000 | 0.000 | 0.000 | HS | 0.999 | 0.000 | 0.001 | 0.000 | 0.001 |
| Gemmeiza-9 | HS | HS | 0.928 | 0.001 | 0.029 | 0.000 | 0.042 | HS | 0.865 | 0.001 | 0.062 | 0.000 | 0.072 |
| DHL11 | S | S | 0.000 | 0.000 | 0.000 | 1.000 | 0.000 | S | 0.000 | 0.000 | 0.000 | 1.000 | 0.000 |
| KSU106 | HS | HS | 0.999 | 0.000 | 0.000 | 0.000 | 0.001 | HS | 0.993 | 0.000 | 0.000 | 0.000 | 0.006 |
| Gemmeiza-12 | S | S | 0.000 | 0.000 | 0.000 | 1.000 | 0.000 | S | 0.001 | 0.000 | 0.000 | 0.999 | 0.000 |
| DHL01 | HS | HS | 1.000 | 0.000 | 0.000 | 0.000 | 0.000 | HS | 1.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| DHL14 | T | T | 0.000 | 0.169 | 0.001 | 0.000 | 0.830 | T | 0.000 | 0.215 | 0.001 | 0.000 | 0.783 |
| DHL29 | T | T | 0.001 | 0.112 | 0.043 | 0.000 | 0.844 | T | 0.003 | 0.177 | 0.129 | 0.000 | 0.691 |
| DHL15 | I | I | 0.000 | 0.105 | 0.845 | 0.000 | 0.050 | HT | 0.000 | 0.894 | 0.000 | 0.000 | 0.106 |
| DHL06 | T | T | 0.000 | 0.114 | 0.226 | 0.000 | 0.660 | I | 0.000 | 0.100 | 0.742 | 0.000 | 0.158 |
| Misr1 | T | T | 0.000 | 0.188 | 0.008 | 0.000 | 0.804 | T | 0.000 | 0.204 | 0.010 | 0.000 | 0.787 |
| DHL05 | HT | HT | 0.000 | 0.755 | 0.000 | 0.003 | 0.242 | T | 0.000 | 0.273 | 0.000 | 0.035 | 0.692 |
| DHL23 | HT | T | 0.000 | 0.133 | 0.000 | 0.000 | 0.867 | T | 0.000 | 0.003 | 0.000 | 0.001 | 0.996 |
| Sakha-93 | T | T | 0.000 | 0.231 | 0.000 | 0.020 | 0.749 | T | 0.001 | 0.418 | 0.000 | 0.025 | 0.556 |
| Pavone-76 | HT | HT | 0.000 | 0.766 | 0.005 | 0.000 | 0.230 | HT | 0.000 | 0.671 | 0.009 | 0.000 | 0.320 |
| DHL08 | HT | HT | 0.000 | 0.813 | 0.018 | 0.000 | 0.170 | T | 0.000 | 0.395 | 0.181 | 0.000 | 0.424 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Ashkar, I.; Sallam, M.; Ghazy, A.; Ibrahim, A.; Alotaibi, M.; Ullah, N.; Al-Doss, A. Agro-Physiological Indices and Multidimensional Analyses for Detecting Heat Tolerance in Wheat Genotypes. Agronomy 2023, 13, 154. https://doi.org/10.3390/agronomy13010154
Al-Ashkar I, Sallam M, Ghazy A, Ibrahim A, Alotaibi M, Ullah N, Al-Doss A. Agro-Physiological Indices and Multidimensional Analyses for Detecting Heat Tolerance in Wheat Genotypes. Agronomy. 2023; 13(1):154. https://doi.org/10.3390/agronomy13010154
Chicago/Turabian StyleAl-Ashkar, Ibrahim, Mohammed Sallam, Abdelhalim Ghazy, Abdullah Ibrahim, Majed Alotaibi, Najeeb Ullah, and Abdullah Al-Doss. 2023. "Agro-Physiological Indices and Multidimensional Analyses for Detecting Heat Tolerance in Wheat Genotypes" Agronomy 13, no. 1: 154. https://doi.org/10.3390/agronomy13010154