1. Introduction
Ultraviolet radiation (UVR) is recognized as an important environmental abiotic stressor causing negative effects on plant development and adversely impacting crop yield. Initial comprehension of morphological damage to plants caused by UVR occurred almost a century ago [
1], and many authors recently and essentially concluded that UVR, particularly its UV-B component, generally causes a negative effect on the plant biomass of several important crops, including mung beans, corn, cotton, soybeans, sugarcane, rice and wheat [
2]. However, few solutions exist to protect plants of any species or cultivar against UVR, leaving large-scale agricultural crops without practical management tools to address it.
Solar ultraviolet radiation (UVR) reaches the planet stratosphere in three wavelength ranges: UV-A, UV-B and UV-C. As UV-C is completely absorbed by oxygen combined with the ozone layer and does not reach the Earth’s surface [
3,
4], most prior studies have covered UV-B due to its stronger level of damage; its photons are more able to break chemical bonds and rearrange molecular structures within plant cells [
5]. As for UV-A radiation, which is unaffected by stratospheric O3 depletion, there has been much less research on its effects on plants [
6].
Agricultural crops are exposed to direct sunlight, which they require for photosynthesis, and consequently they simultaneously receive UV radiation, which could be a major limiting factor in the photosynthetic efficiency of crop plants [
7]. Failure to protect from UV-B may result in a wide range of morphological, physiological and metabolic responses, including altered plant growth, reduced yield and damage to photosystem II (PSII) [
8]. In a review of 129 studies on 35 crop species published since 1975, the authors verified that UV-B exerts negative effects on plant biomass and its reproductive organs [
2]. Even the roots of a plant, although below ground, may negatively react to the plant’s leaves being exposed above ground to ambient levels of UV-B [
9]. As a result of UVR stress, an imbalance occurs in cellular homeostasis which is mainly caused by reduced photosynthesis rates and changes in plant metabolism [
10], and induces ROS (reactive oxygen species), which cause lipid peroxidation and consequent damage to cell membranes [
11]. Under stress conditions, plants adapt through physiological and metabolic responses regulated at the transcriptional level [
12,
13]. Ultraviolet radiation is an energetic driver of a diverse range of plant responses, and may be exploitable in the context of a sustainable contribution toward strengthening global crop production [
14].
Several major agricultural crops have been subject to assessments of UV radiation’s impact [
15]. A study exposing corn to four UV-B doses (0, 5, 10, and 15 kJ m
−2 day
−1) demonstrated that enhanced or current ambient levels of UV-B radiation can adversely affect corn growth [
16]. In another study with cotton, plants were exposed to three levels of UV-B (0, 7 and 14 kJ m
−2 day
−1) in a factorial with three day/night temperatures, evidencing an interaction between UV-B and temperature that negatively impacted yield [
17]. A two-year study using three cultivars of soybeans exposed to enhanced UV-B demonstrated its negative impact on yield components, with a decrease in number of pods and average yield reduction per plant of 43.7% [
18]. Another study with several soybean cultivars under the exclusion or presence of ambient UVR showed that UVR always caused a negative impact on biomass and yield over all cultivars [
19]. High accumulation of proline is a common physiological response in plants exposed to various abiotic stresses, including UV radiation [
20]. An important marker for oxidative stress in living organisms occurring in response to UV radiation is malondialdehyde (MDA), content of which may rise even at minor levels of exposure to UV-A radiation [
21]. To address this issue, some researchers propose that breeding programs develop more adapted cultivars to UV radiation [
14], particularly in response to ongoing climate change and with priority for tropical and subtropical zones.
Breeding programs require a considerable lead time to deliver new high-yielding varieties, which generally takes 10 years [
22]. Thus, practical near-term solutions to protect large-scale field crops against excessive UV damage through exogenous spray of a composition may be an alternative. One approach for mitigating UV stress in plants has been proposed through plant nutrition with soluble silicon (Si); it leads to Si accumulation in the form of biogenic silica (amorphous form) under the epidermis of the leaves, which then operationally reduces UVR transmission from the external environment to the inside of the leaf [
23,
24]. However, not all plant species or their varieties present the ability to absorb, translocate and accumulate Si, and later to form biogenic silica [
24,
25].
We hypothesized that mitigation of UV damage could be achieved by depositing a layer with fine particles of insoluble crystals of quartz silica sand on the external plant part over the leaves. Thus, we undertook a study in soybeans (Glycine max (L.) Merrill) in a greenhouse under three scenarios of UV radiation exposure (no UV, +UV-A or +UV-B), and for each situation, imposed foliar sprays both with and without a suspension with a composition principally based on this mineral. This was followed by assessment of the plants for (1) the presence of biochemical compounds related to abiotic stress and (2) phenotype and yield parameters to determine the effects of UV-derived stress, and the potential of the mineral composition in mitigating such stress.
2. Materials and Methods
2.1. Plant Cultivation Description
Seeds of soybean (
Glycine max (L.) Merrill) cultivar NA5909 RG were cultivated in pots under greenhouse conditions. The pots had a capacity of 8 L and were filled with cultivation substrate mix composed of organic material (product brand BIOMIX purchased at market and composed by crushed bark of pine/eucalyptus and powder of coconut fiber, pH 6.5, bulk density = 0.3 Mg m
−3) and sand in a proportion of 3:1, respectively. The seeds used are of a commercially registered and available cultivar and were obtained as a sample of a commercial seed lot from a private grower in the São Paulo state of Brazil. A total of 12 pots were sown with 15 seeds in each pot as of 22 September 2021, and after germination, plantlets were selected at the VC stage (when cotyledons are above ground and unifoliate leaves are unrolled) to thin them to the best two seedlings per pot. The greenhouse was covered with a translucid anti-UV plastic film (brand Suncover Diff, Code CT-C655, 150 micra, manufactured by GINEGAR), which allows 89% transmittance of PAR (photosynthetically active radiation) from sunlight and promotes diffusion of 60% of it in the chamber to reach the plants uniformly, while retaining 99.9% of UV frequencies (as measured with an INSTUTHERM MRU-201 reader). During the cultivation of the plants, the total sunlight radiation of UV+PAR on the region and over the greenhouse was an average of 11.5 MJ m
−2 day
−1 (data obtained from INMET—National Institute of Meteorology of Brazil). Plants were then irrigated once a day in the morning with 400 mL of water per pot and fertilized once a week with a solution containing key nutritive elements as suggested by EMBRAPA [
26]. All methods were carried out in accordance with relevant guidelines and legislation.
2.2. Treatment Description
The factorial design involves a three UV exposure regimen (no UV, +UV-A or +UV-B) using two mitigation treatments (sprayed or unsprayed) with or without the foliar application of a composition aiming to mitigate damages by UV exposure. The composition used is as described in the patent application WIPO/PCT WO2022/011441 (based on 45% w/v of inert, insoluble, crystal, micronized quartz sand, 12% w/v of zinc oxide and 17% w/v of manganese sulfate, presented as a concentrated suspension and purchased in the Brazilian market, branded as ACLIMAT), which, once sprayed over plant leaves, fundamentally acts as an external physical barrier to reduce the entrance of UV rays into the tissues. The product label claims to contain adjuvants to ensure optimal coverage and to limit wash-off; thus, no other product was added for spraying. Four replicates were used for each treatment, each replicate being represented by one plant; thus, two pots, each one having two plants, were used per treatment.
For the first objective of the study (UV impact), the greenhouse was divided into three compartments (chambers) using the same anti-UV plastic film to isolate radiation from each one. The first compartment remained without UV radiation, the second was exposed to artificial UV-A radiation and the third to artificial UV-B radiation. Irradiation for each UV type over the plants took place during 37 days from 29 DAE (days after emergence) to 66 DAE. The +UV-A chamber was irradiated with Philips TL-K 40 W/10-R lamps (manufactured by Koninklijke Philips N.V., purchased in São Paulo, Brazil), always placed 60 cm above the top of the plants, emitting 1080 µW cm−2 for 600 min per day (starting at 7:00 am); this is equivalent to 389 kJ m−2 day−1. The +UV-B chamber was irradiated with Philips TL 40 W/12 lamps (manufactured by Koninklijke Philips N.V., purchased in São Paulo, Brazil), always placed 90 cm above the top of the plants, emitting 330 µW cm−2, for 45 min per day (three sessions of 15 min spaced at 60 min intervals, around midday); this is equivalent to 9 kJ m−2 day−1. The radiation intensity was measured and monitored at the top 10 cm level of the plants. The pots were rotated once a week within each chamber to ensure uniform exposure growing conditions.
For the second objective, four plants (two pots) under each of the three UV regimens were subjected (sprayed) to a foliar spray with 2.6 mL per plant of a 0.5% water suspension with the composition, whereas the other four plants (two pots) serving as controls were sprayed only with pure water (unsprayed). The first foliar spray occurred at the V3 (third vegetative node with unfolded trifoliates) crop stage and was repeated once a week for five weeks.
2.3. Biochemical Analysis of Plant Leaves
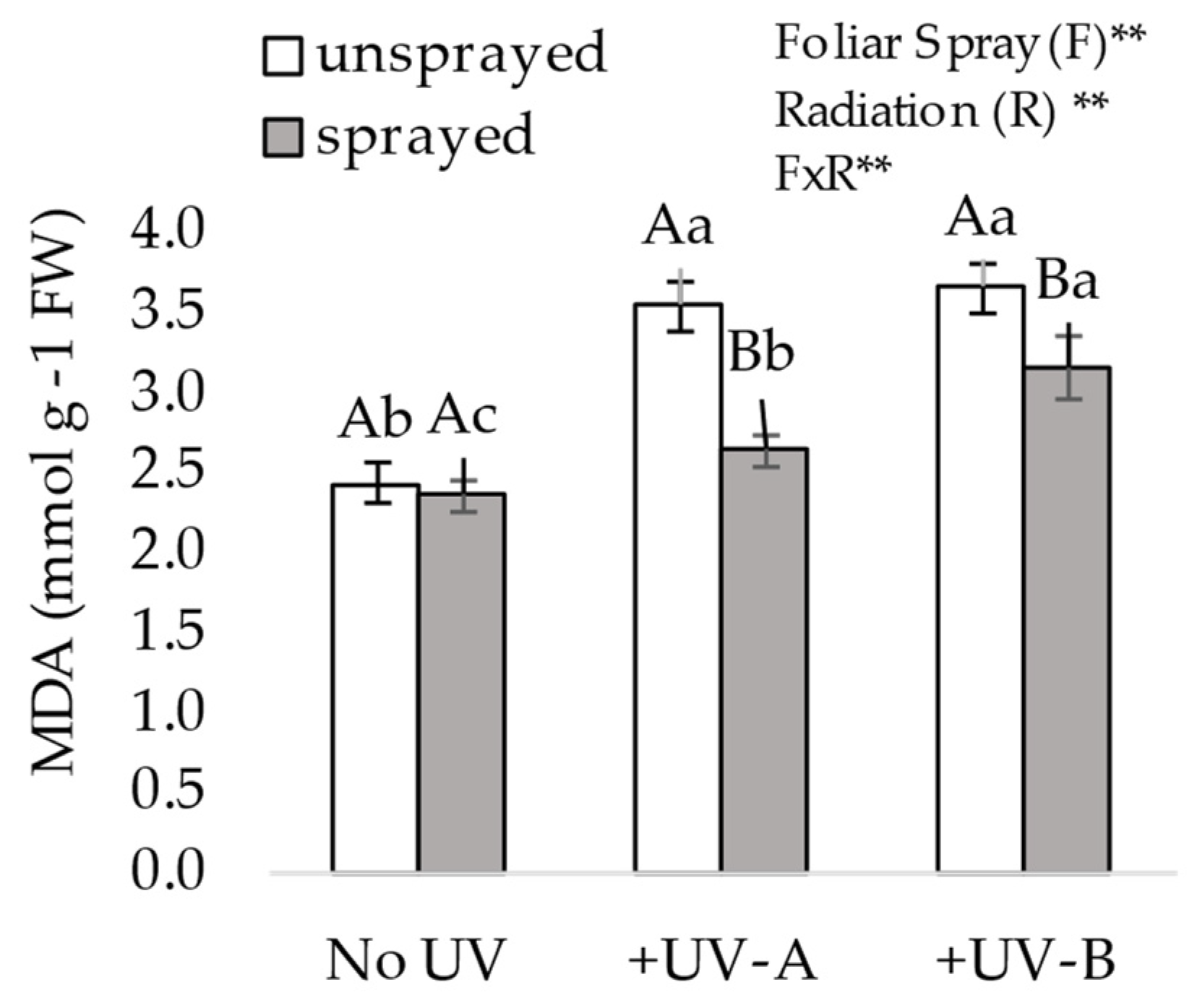
At 84 DAE, eighteen days after radiation over plants ended, one leaflet of each trifoliate of each plant serving as replicate was extracted without the petiole, forming a bulk per plant set in layers in the same sequence of its position along the stem. This was then promptly measured for its fresh weight and immediately thereafter packed in an aluminum ziplocked envelope and stored at −80 °C for further analysis. For each analysis, a piece of the bulk was extracted to represent the whole plant as much as possible. The primary aim of the study was to assess the effect of mitigating UVR impact on biomass, and the biochemical analyses presented here are those of key indicators that support the evidence of plant stress levels, i.e., malondialdehyde (MDA) and proline.
2.3.1. Malondialdehyde (MDA) Content
Thiobarbituric acid reactive substances’ (TBARS) content was used to estimate lipid peroxidation. Malondialdehyde (MDA) equivalent concentration was calculated using an extinction coefficient of 1.55 × 10
−5 mol
−1 cm
−1, with readings between 535 and 600 nm. The results were expressed in mmoL mg
−1 of fresh tissue. The content was determined as described by Heath et al. [
27] with adaptations by Gratão et al.
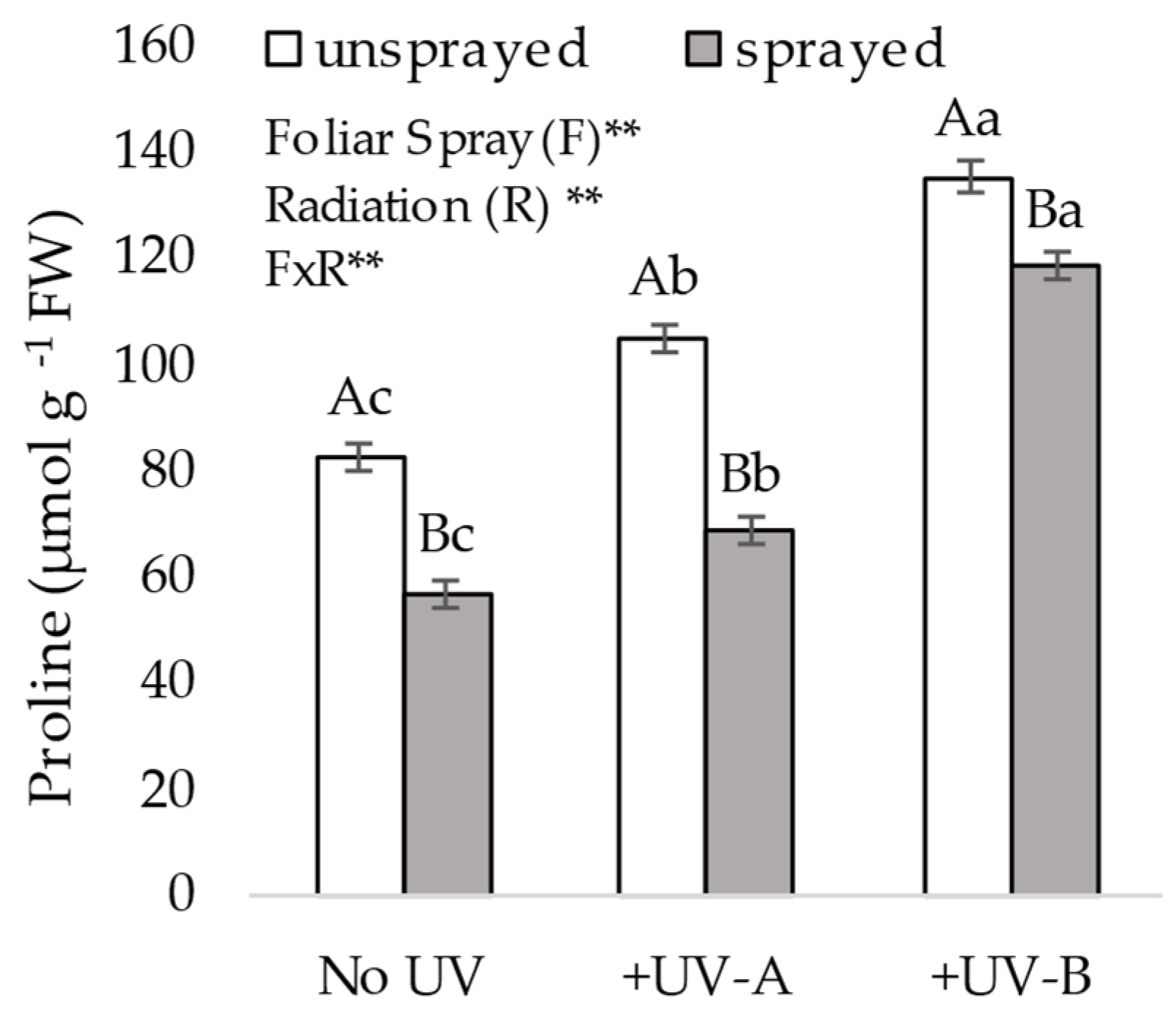
2.3.2. Proline Content
Fresh leaves were homogenized in 3% sulfosalicylic acid and filtered. The mixture filtrate was added to 1 mL each of acid ninhydrin and glacial acetic acid and was placed in boiling water for 1 h, and toluene (4 mL) was added to the mixture. Absorbance was measured spectrophotometrically at 520 nm and converted to µmol g
−1 fresh weight against standard proline. The proline content was determined as described by Bates [
28].
2.4. Biometric Assessments of Plants
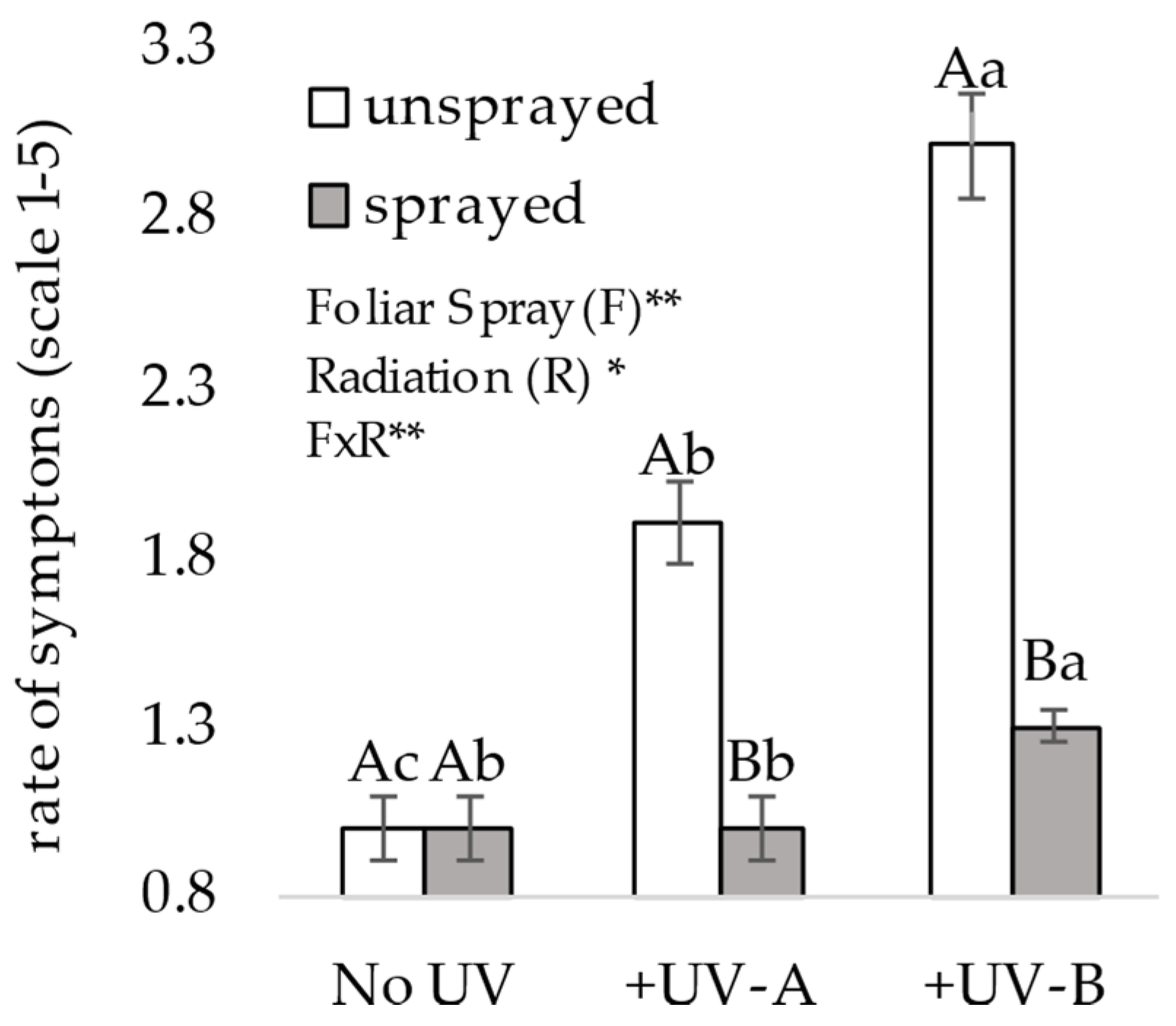
2.4.1. Description of UVR Impact Visual Assessment on Leaflet
Individual plants were assessed periodically to determine the visual effect on morphological symptoms from UV stress using a scale from 1 to 5, where 1 indicated no symptoms, 2 indicated a slight reduction in leaf size, 3 indicated some crispy wrinkle aspects on leaves, 4 indicated browning and 5 indicated high symptoms of necrosis. Assessments started one week after the initiation of UV radiation and were repeated once a week, with each assessment focusing on the trifoliate recently formed and after its exposure to UVR during the preceding five to ten days, thus allowing us to obtain an average rating during plant development.
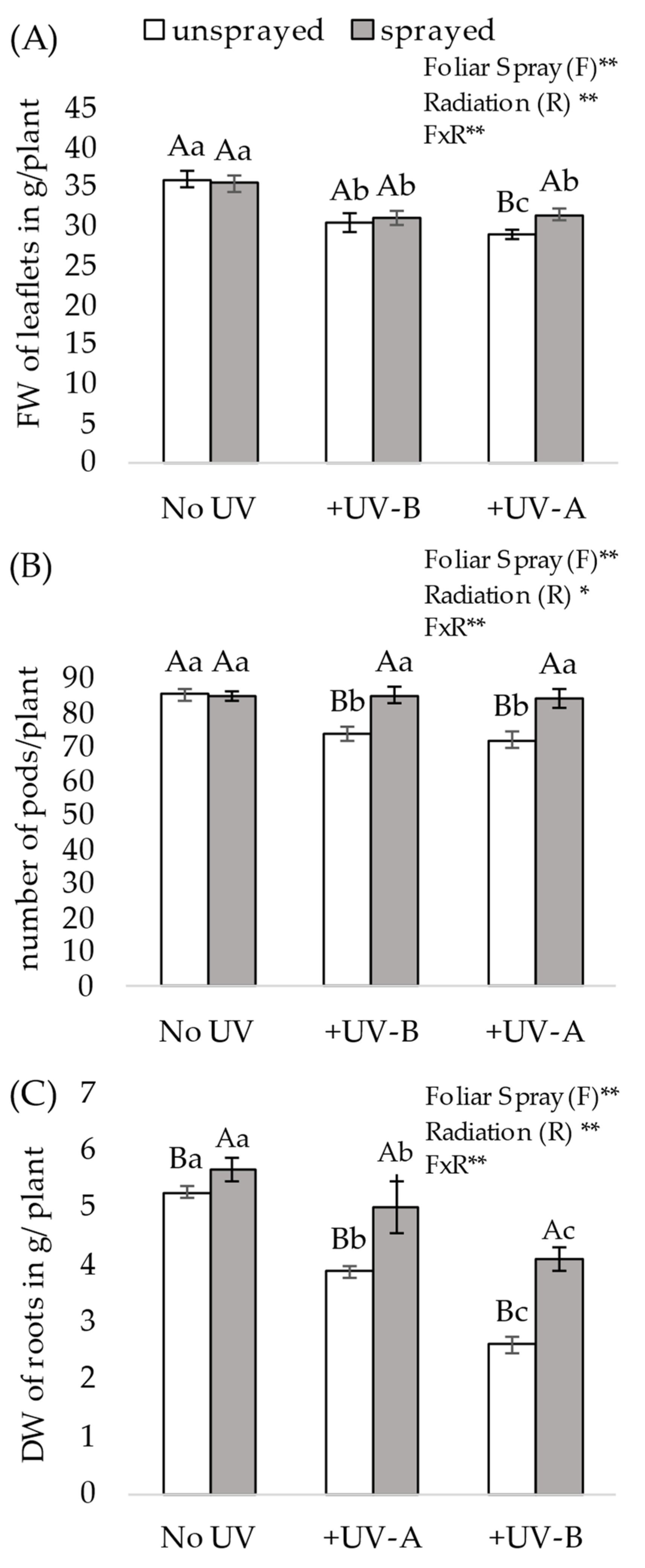
2.4.2. Description of UVR Impact Assessment on Aerial Biomass
Plants were harvested at 85 DAE when pods were formed and reached the R5:3 stage (grain being filled into the pod). The individual plants had their stems cut 15 cm from the soil surface, their fresh weight was measured for leaflets and the number of pods per plant was counted. Leaflets were removed without the petiole and joined in a bulk per plant, and their fresh weight was promptly measured.
2.4.3. Impact on the Number of Pods per Plant
All reproductive structures, ranging from R3 (pods just fertilized and size of 2–3 mm) to R5:3 (grain being filled into the pod), were extracted from plants and counted, with the recognition that they may not represent the final plant yield, since between the stage of pod removal and plant senescence, the younger pods at the R3 or R4 stage (a pod completely developed but without visible grain in it) may abort or fail to become sufficiently mature to be represented in the true final yield. The pods counted ranged from those greater than 1 cm (recently fertilized) up to pods in full grain filling, serving as an indicator of plant yield potential under stress and after mitigation.
2.4.4. Impact on Roots
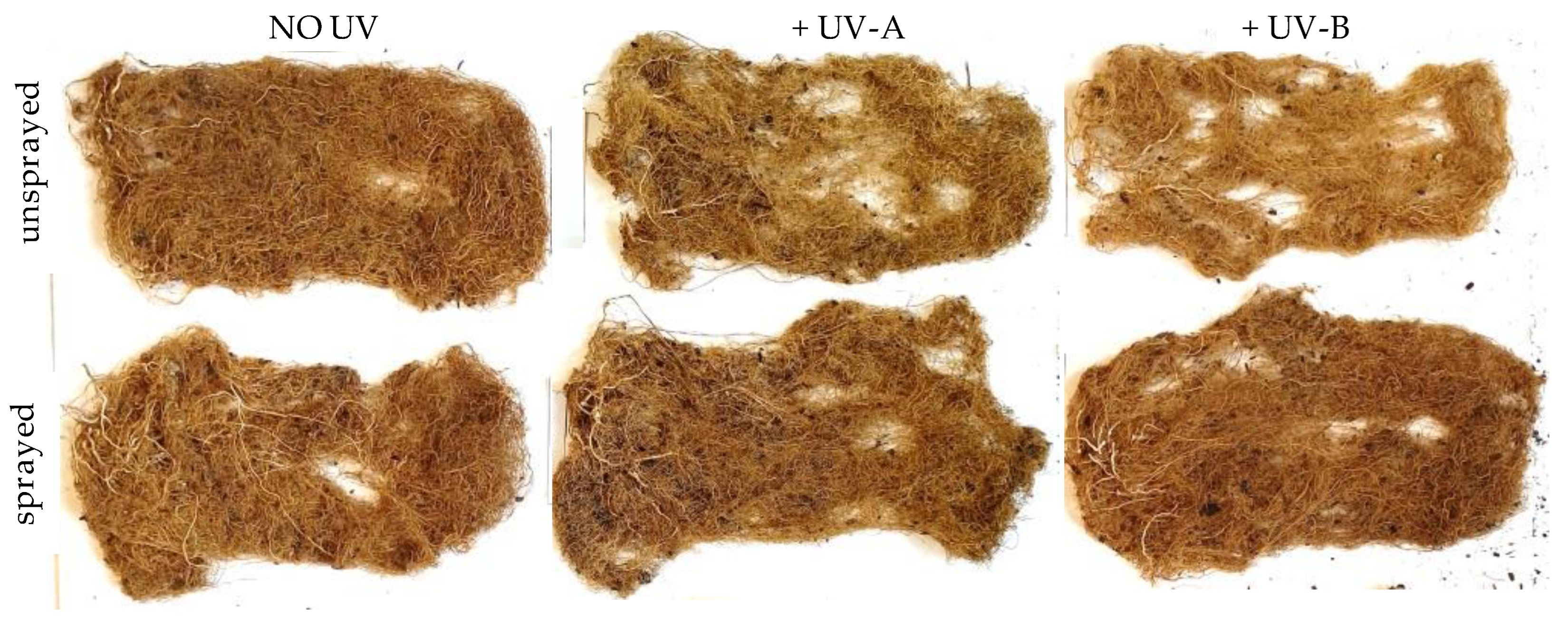
After the removal of the aerial structure of the plants, the pots containing a piece of remaining stem and root were carefully washed to remove the cultivation substrate and preserve as much root tissue as possible. To dry the cleaned roots, they were hung and exposed to the sun during the entire day. The remaining debris from the cultivation substrate still attached to the roots was then removed by gently shaking it. Once clean and dry, the remaining piece of the stem and of the hypocotyl of the pivot larger than 2–3 mm was removed with scissors to assess solely the dry weight of the true root component.
2.5. Statistical Analysis
Statistical analysis was performed by AgroStat software [
29]. Data were submitted to variance analysis using the F test, and any significant differences between treatments were compared using the Tukey test at 5% probability.
4. Discussion
Most studies concerning UVR impact in plants have focused on UV-B because of its higher energy aspect and erythemal injury, and those studies that cover UV-A effects are frequently performed in combination with UV-B, as both reach the earth surface simultaneously with sunlight. Indeed, plant responses to UV-A radiation have been less frequently studied than those to UV-B, leaving an important gap in our understanding of the response to solar UV radiation by plants [
30].
Whereas prior studies demonstrate that UV-B (280–315 nm) causes losses in the biomass and yield of most crop plants [
2], many of the effects on growth and development exerted by UV-A are distinct from those triggered by UV-B, and vary considerably in terms of the direction of the response [
31]. The present study separately analyzed exposure to the two wavelength ranges to better understand the effects of each wavelength range over soybeans.
Biochemical assays were performed for two key components responsive to abiotic stress, MDA and proline. The data presented in our study are in syntony with the plant morphology measurements, and show good correlation with visually observed impacts and mitigation treatments. In a similar study in soybean exposed to UV radiation, a higher MDA content and a reduction in the number of pods per plant were shown to be caused by such stress [
32].
In our present study, MDA content is higher in both UV regimens in comparison with no UV, and of very similar content in both +UV-A and +UV-B exposure for unsprayed plants; although, the damage observed in the biomass is not necessarily of equal magnitude. Similarly to what has been reported in a prior study on the responsiveness of MDA in living organisms to UV-A [
21], MDA seems to be equally and highly responsive to both types of UV stress; the data of this present study suggest that UV-A may be as harmful as UV-B to plant physiology, as MDA possibly indicates the higher accumulation of ROS in unsprayed plants for both UV types. Although complementary analysis to measure specific ROS products could further differentiate the damage caused by each UV wavelength, the MDA content here shows that both UV-A and UV-B cause damage. The accumulation of ROS causes increased levels of peroxidation of membrane lipids, and MDA content indicates this type of cell damage [
33]. In our study, the MDA content from leaflets sprayed with the composition is significantly lower than that in unsprayed leaflets, with greater mitigation shown under +UV-A exposure. This may indicate either that the composition is more effective in mitigating UV-A, or that UV-B is indeed a much more stressful wavelength, even though MDA reduction is significant in both exposure cases when plants are sprayed.
The measurements of proline in the present study following exposure to +UV-A and +UV-B are in line with the observations of other prior studies [
34] on the effects of UV-A and UV-A/UV-B on soybean seedlings, which found a 25% lower content of proline in plants isolated from exposure to both UV-A and UV-A/UV-B. In our study, we observed both a proline response to UV regimens and a different response following foliar treatment, showing mitigation of the UV effect. As opposed to MDA, proline content progressively increases as the observed damage on plant biomass is higher, and follows the respective levels of the effects of +UV-A and +UV-B. This is in accordance with well-established scientific documentation that indicates proline accumulates in response to environmental stresses, and can act as a signaling molecule to modulate mitochondrial functions, influence cell proliferation or cell death and trigger specific gene expression, which can be essential for plant recovery from stress [
35]. The accumulation of proline in response to UV-A, UV-B and UV-C occurs even in well-adapted desert plants acclimated under high levels of sunlight [
36]. An interesting finding in this study is the lower proline content in sprayed plants under all three UV regimen cases. For +UV-A and +UV-B cases, it is reasonable to infer that the composition applied over the top of plants formed a layer that prevented high levels of UV radiation from entering the leaves; thus, the organism was not required to increase proline for self-protection against this abiotic stress. For the no UV situation, we hypothesized that some other stress factor occurred simultaneously; it is possible that fluctuation in soil moisture due to watering only once a day led the leaves to experience variation in their turgor, and as the microparticles of quartz sand over the leaves preserve some leaf turgor (prior observation from authors, not published), such a change in a different concurring stress factor might have led to a lower content of proline in sprayed plants in the no UV situation. This could also explain the slightly higher root weight of the no UV plants. The proline assay suggests that the foliar treatment mitigated the stress caused by +UV-A and +UV-B.
A few prior studies measuring MDA and proline in connection with stress caused by UV radiation usually assayed the leaves during or immediately after exposure to UV [
23,
37,
38]. An interesting observation from this study is that leaves were collected 18 days after irradiation ended, and the variable content of these compounds was still measurable, possibly indicating that the effects of UV are long lasting in plant tissues.
In the present study, both UV-B and UV-A cause negative impacts on plant morphology, physiology and yield potential. The assessment of visual stress on leaflets of plants shows that UV-A presents a soft visual morphological effect when compared with UV-B. However, both UV-A and UV-B cause damage to root volume, foliar biomass and yield potential due to the number of pods per plant. Under exposure to +UV-A and +UV-B, the impact is negative and more severe on unsprayed plants, in contrast to sprayed plants, for which the mineral composition mitigates such deleterious effects.
The impact on root development caused by both wavelengths is of great importance, as it will limit yield potential or cause yield variations under common field situations in the event of insufficient or irregular rain distribution. UV-B causes roots to have less dry weight or deformity due to bending induced by UV-B radiation through a flavonoid-mediated phototropic response [
39], and it has been demonstrated that exposure to UV-B induces an inhibition of primary root elongation because of a decrease in cell proliferation in the meristematic zone of the primary roots [
40]. Here, the study assesses the effect of both frequencies of radiation (+UV-A and +UV-B) and verifies a reduction in root weight of 50% under +UV-B and 29% under +UV-A, which are very close to the observations of a prior study [
41] that found a reduction of 41% under UV-B and 51% under combined UV-A/UV-B, thereby also confirming the effect of UV-A. The present study also aligns with other findings [
42] that measured the root weight of soybean seedlings grown under exclusion of UV-B or at two levels of exposure (2.63 kJ m
−2 day
−1 or 6.17 kJ m
−2 day
−1) to elevated UV-B radiation and showed that both levels of exposure inhibited root growth by an average of 30% dry weight compared with the control. The data presented here shows that a foliar spray with the composition (sprayed) attenuates the impact of both UV-A and UV-B, since sprayed plants develop better roots; this result is in conformity with root measurements in a prior study that showed higher dry root weight when soybean plants were grown under the exclusion of UVR effects, using a UV filter shield over the plot to protect the plants [
41]. The importance of these findings is that protecting crops through reduction of UVR exposure through the common practice of foliar spray will allow improved root development, a critical feature for optimal water and nutrient extraction from the soil, thus contributing to higher and/or more stable yields that are impacted by exposure to ambient UV radiation, an important advancement in the face of climate change.
The present study shows that the fresh leaf biomass of unsprayed plants is also impacted by both +UV-A and +UV-B in the order of −16% and −19%, respectively, and sprayed plants show a slight mitigation of this negative effect, but not to the level observed in the no UV situation. The effect found remains significant and aligns with the findings of other studies [
18,
41].
Regarding yield, the present study confirms a previous report that UV-B causes a reduction in yield potential, notably due to a smaller number of pods per plant, with a reduction of 25% in the number of pods per plant with supplemental UV-B over ambient UV-B [
38]. Conversely, in a prior study applying the exclusion of UV-B to 8 soybean cultivars, an increase in the number of pods per plant of up to 43% was found [
19]. In a 2-year study of the effects of UV-B on soybean yield, it was determined that yield reduction was mainly attributable to a change in the number of pods per plant under UV-B radiation, which decreased the number of pods per plant of three soybean cultivars by 34.5% on average [
18]. In the present study, the yield potential represented by the number of pods per plant is impacted both by +UV-A and +UV-B, where unsprayed plants show a reduction of 14% and 16% in pod structures under +UV-A and +UV-B, respectively, in comparison with a no UV regimen. Interestingly and as hypothesized, sprayed plants in the same +UV-A and +UV-B stressing environments show stress mitigation and present no yield loss, whereas, no difference in the number of pods per plant occurs between sprayed and unsprayed plants in the absence of such UV stress.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





