1. Introduction
Phelipanche aegyptiaca is an annual root parasitic weed that is widely distributed in temperate and subtropical regions of the Mediterranean, southern and northern Africa, eastern and southern Europe, and South Asia [
1,
2,
3]. This parasite of the Orobanchaceae family is one of the most harmful to agricultural crops worldwide. In China,
P. aegyptiaca has spread almost all over Xinjiang, severely affecting the production of the main crops, such as tomato, melon, and sunflower. It has given rise to a large area of production reduction, or even almost no harvest, causing a loss of more than CNY 500 million to the local area each year [
4,
5]. Hence, the prevention and control of
P. aegyptiaca is in an important position. As an obligate parasitic weed,
P. aegyptiaca parasitizes the roots of host crops to obtain water and nutrients through the haustoria, affecting the normal growth of crops and causing agricultural losses. Due to the huge seed bank of
P. aegyptiaca in the soil and its unique parasitic characteristics, traditional agricultural and chemical methods do not effectively control its hazards. Therefore, blocking the nutrient absorption channel and reducing the harm of
P. aegyptiaca to the host are of great scientific significance for its control.
The life cycle of
P. aegyptiaca is the same as the majority of the holoparasitic Orobanchaceae, including the seed phase, the autotrophic growth phase, and the heterotrophic growth phase. In the seed phase, the seeds of
P. aegyptiaca dormant in the soil to form a huge seed bank, which germinated after responding to allelochemicals, such as strigolactones (SLs) secreted by the host roots, and then enter in an autotrophic growth phase. Subsequently, the radicle of
P. aegyptiaca tends to grow toward the host root and forms terminal haustorium under the induction of HIFs when approaching the host root. Then
P. aegyptiaca invaded the host by means of the haustorium, established the parasitic relationship, and entered the heterotrophic growth phase [
1,
6]. Thus it can be seen that the haustorium is the special organ of parasitic plants, and its formation is a critical step for the transition from an autotrophic to a heterotrophic lifestyle [
7]. Obligate parasitic plant
P. aegyptiaca forms terminal haustoria to attach and penetrate the host root and obtain sugars, water, and other nutrients required for life by developing the vascular connection between the parasite and the host, thus causing damage to host growth and development [
8]. In addition, haustoria are also an important channel for material exchange and information communication between hosts and parasites, such as viruses, siRNAs, nucleic acids, and proteins, which are transferred between the parasite and host through the haustoria [
9,
10,
11,
12]. Therefore, the haustorium initiation and development are crucial steps in the lifecycle of parasitic plants [
1,
7,
13]. Nevertheless, haustorium initiation is not spontaneous during the growth of parasitic plants; only contact with host- or non-host-exudated chemical stimulation under appropriate temperatures and humidity conditions can trigger haustorium initiation and development. The chemicals that induce haustorium initiation in the Orobanchaceae are called haustorium-inducing factors (HIFs) [
7]. Searching for HIFs is helpful in understanding the haustorium formation of parasitic plants and in applying it to the control of
P. aegyptiaca. However, their synthetic pathways and specific sources are largely unknown. Numerous studies have shown that the root exudates of numerous plant species can induce prehaustoria [
7,
14,
15,
16]. The root exudates extracted from the highly susceptible
Brassica napus cultivar had a high induction activity for the haustorium formation of
Phelipanche ramosa, and parasite aggressiveness was significantly enhanced after treatment with root exudates [
14]. Root exudates and tissue extracts of rice and
Arabidopsis can induce
Phtheirospermum japonicum and
Striga hermonthica haustorium formation [
15], which confirmed that host root exudates contain HIFs of parasitic plants. In 2022, the planting area of
Cucumis melo L. in Xinjiang was 626,500 hectares, with a total output of 2,116,600 tons, as an important economic crop in Xinjiang,
C. melo has been seriously damaged by
P. aegyptiaca. Although it is one of the main hosts of
P. aegyptiaca, the effects of its root exudates on the germination and haustorium development of
P. aegyptiaca have not been reported. The interaction mechanism between
C. melo root exudates and
P. aegyptiaca haustorium is also unknown.
At present, an increasing number of allelochemicals have been found and extracted from plant root exudates, and the haustorial induction activity of individual allelochemicals has been determined. It has been reported that the types of parasitic plant HIFs in plant root exudates mainly include phenolic acids, quinones, flavonoids, lignin units, cytokinins, and cyclohexene oxides [
13], which show different haustorial induction activities for different parasitic plant species. The first HIF discovered, 2,6-dimethoxybenzoquinone (DMBQ), was isolated from sorghum roots and shown to trigger haustorium formation in
Triphysaria versicolor and
Striga hermonthica [
16,
17]. Compounds with structures similar to DMBQ, such as phenolic acids (including syringic acid, vanillic acid, and ferulic acid), aldehydes (including syringaldehyde), and flavonoids (including peonidin), have also been reported to induce haustoria in
Triphyasria versicolor,
Phtheirospermum japonicum, and
Striga hermontica [
15]. The mycotoxins sphaeropsidone and epi-sphaeropsidone, which are cyclohexene oxides isolated from
Diplodia cupressi, the causal agent for cypress (
Cupressus sempervirens L.) canker, have HIF activity in vitro toward
Orobanche crenata and
O. cumana [
18,
19]. Treatments with phytohormones also trigger haustorium formation. In addition, auxin efflux activity within the haustoria at later developmental stages is crucial for xylem bridge formation [
20]. Ethylene signaling in the parasite is crucial for the haustorium apex cells to differentiate into intrusive cells for host invasion. While the exogenous application of ethylene suppresses haustorium development, ethylene signaling is dispensable during early haustorium initiation, but is required for the timely termination of haustorium growth in the absence of a host [
21]. Nitrogen increases the levels of abscisic acid (ABA) in
P. japonicum and prevents the activation of hundreds of genes, including cell cycle and xylem development genes. Blocking ABA signaling overcomes nitrogen’s inhibitory effects, indicating that nitrogen represses haustoria formation by increasing ABA [
22]. Cytokinins constitutively exuded from host roots play a major role in haustorium formation and aggressiveness in
P. ramose [
14]. In addition, ROS and ROS-regulating enzymes are indispensable in the downstream signaling of HIFs for haustorium formation [
23]. In
P. japonicum, DMBQ initiates Ca
2+ signaling in the root and is important for the development of the haustorium [
24]. However, the regulation of these allelochemicals and pathways on
P. aegyptiaca haustorium has not been reported.
Here, based on the resistant and susceptible C. melo cultivars KR1326 and K1076, respectively, obtained by previous research, there was a significant difference in the induction effect of root exudates of KR1326 and K1076 on seed germination and haustorium formation. Using widely targeted metabolomics based on ultra-high performance liquid chromatography–tandem mass spectrometry (UPLC-MS/MS) technology, the metabolites in root exudates of P. aegyptiaca were analyzed and identified. Through multivariate statistical analysis methods, such as principal component analysis (PCA) and orthogonal partial least squares discriminant analysis (OPLS-DA), the differential metabolites in the root exudates of KR1326 and K1076 were screened, and the haustorium induction activity of allelochemicals that could be used as potential HIFs was tested. Thus, the allelochemicals in C. melo root exudates that can be used as HIFs of P. aegyptiaca were screened. This sheds new light on the control of P. aegyptiaca by regulating the haustorium development and provides clues for determining the mechanisms by which parasitic plants respond to host root-derived molecules.
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
In 2020, the seeds of
P. aegyptiaca were collected from tomato fields in Jimusar County, Xinjiang province, China. These seeds were stored at 25 °C in the dark before use. Cucumis melo L. cultivars KR1326 (resistant) and K1076 (susceptible) were provided by the Hami Melon Research Center, Xinjiang Academy of Agricultural Sciences.
Cucumis melo seeds were surface-sterilized using 2% (
v/
v) sodium hypochlorite and germinated in sterilized water for 2 days. Germinated seeds were planted in a 50-hole (5 × 5 × 5 cm) plastic cave dish containing vermiculite medium, grown in a growth chamber at 25 °C with a photoperiod of 16-h light and 8-h dark for 10 days, and regularly watered once every two days. Cucumis melo seedlings with one fully developed true leaf were transferred to a black hydroponic box [1200 mL, 15 × 20 × 7 cm (W × L × H)] filled with 1000 mL Hoagland nutrient solution (
Table S5). After rinsing the root with sterilized water at least three times, fresh nutrient solution was supplied every 3 days.
Phelipanche aegyptiaca seeds were sterilized with 75% (
v/
v) ethanol for 2 min and 2% (
v/
v) sodium hypochlorite for 15 min and then rinsed with sterilized water at least 5 times. Pre-conditioning, germination, and haustorium induction of
P. aegyptiaca seeds were performed in an incubator (A1000, CONVIRON, Winnipeg, MB, Canada) at 25 °C in the dark.
2.2. Collection and Treatment of Root Exudates
The root exudates were collected from C. melo seedlings at the flowering stage (33 days old) by means of hydroponic culture. To collect root exudates, six C. melo plants were moved to a clean hydroponic box filled with 300 mL sterilized water at 25 °C with a photoperiod of 16 h light and 8 h dark for 24 h. Three replicate samples were prepared for each experiment. The aqueous solution was collected from the hydroponic box and centrifuged (12,000 r/min, 4 °C) for 10 min. The supernatant was then immediately concentrated to 10 mL in a rotary evaporator (RV 10 D S96, IKA, Königswinter, Germany). Concentrated root exudates were used for P. aegyptiaca seed germination and haustorium induction after filtering with a microporous filter membrane (0.45 μm).
For metabolite extraction, 9 mL of each sample was placed in a 50-mL centrifuge tube after mixing, and the samples were immersed in liquid nitrogen. The samples were freeze-dried in a lyophilizer (SCIENTZ-12N/D, Zhejiang, China) after being completely frozen. Then, 300 µL of 70% methanol internal standard extract was added and scrolled for 15 min. The samples were then centrifuged (12,000 r/min, 4 °C) for 3 min. The supernatant was filtered with a microporous filter membrane (0.22 μm) and stored in a sample flask for UPLC-MS/MS.
2.3. Quantification of Seed Germination and Haustorium Induction
For pre-conditioning, 100
P. aegyptiaca seeds were evenly placed on a Petri dish with two pieces of moistened filter paper after surface sterilization and incubated at 25 °C in the dark for 5 days. For seed germination,
P. aegyptiaca seeds were treated with 1 mL of root exudates for 6 days after pre-conditioning. GR24 has been widely used in the seed germination of
Orobance [
25,
26] and thus was used as a positive control.
For haustorium induction, after rac-GR24 (1 × 10
−7 mol/L) treatment for 3 days, germinated
P. aegyptiaca seedlings were induced with
C. melo root exudates and each chemical compound (
Table S1) for 7 days before quantifying haustoria directly with a stereomicroscope (ZEISS AxioCam ICc 5, Baden-Württemberg, Germany). One
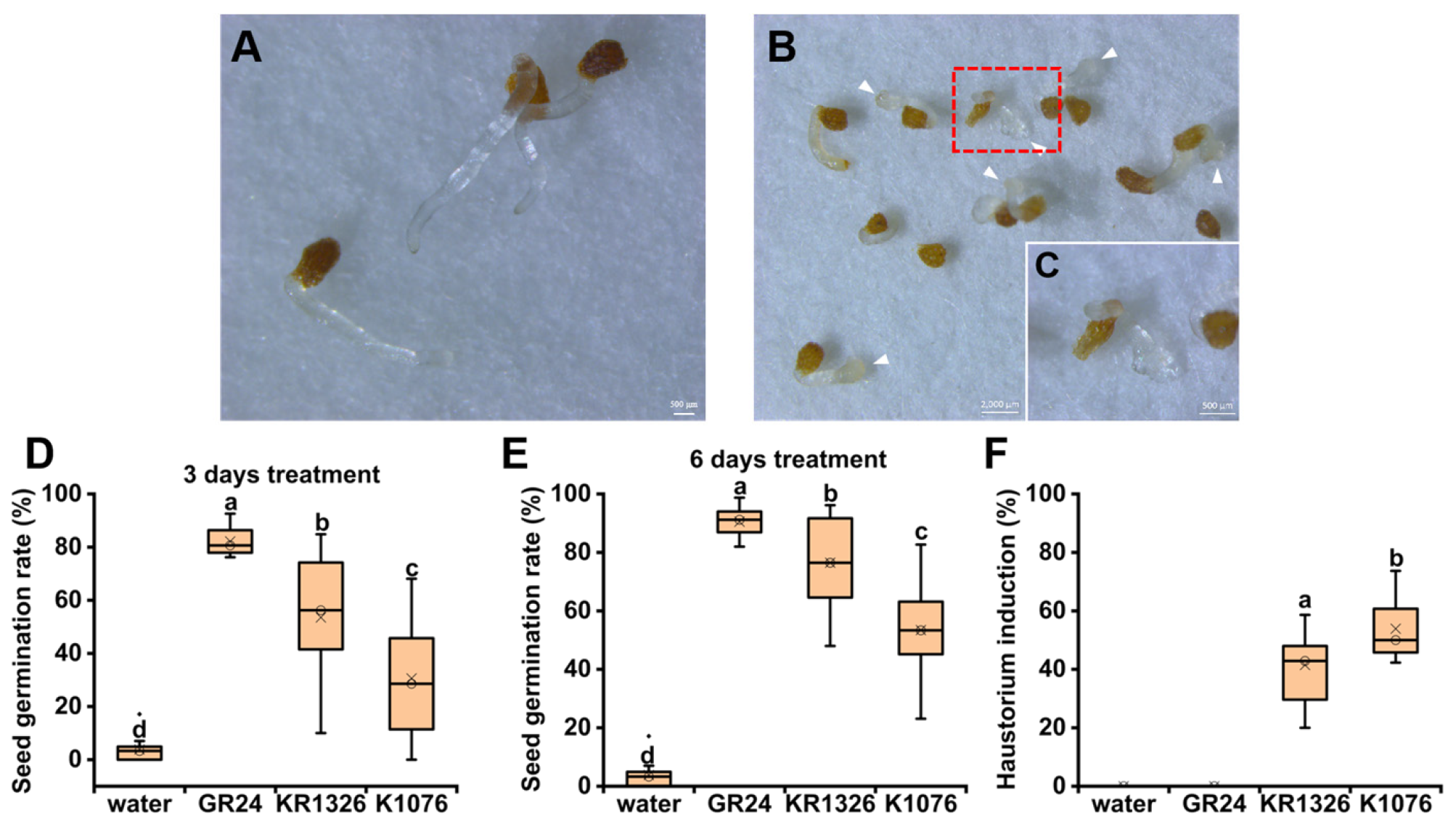
P. aegyptiaca seed can form only one terminal haustorium, developing a swollen and multicellular structure in the root apex (
Figure 1C). Therefore, we used the number of seeds that formed an obvious haustorium structure characteristic to measure the haustorium induction rate of each chemical by calculating the percentage of haustorium formation in
P. aegyptiaca seeds after treatment with different concentrations of each compound (
Table S1).
Seven metabolites were selected from the differential metabolites to evaluate their haustorium-inducing activity. According to previous studies, host-mediated cytokinins, ethylene signal, and auxin efflux activity play an important role in the regulation of haustorium formation [
25,
26]; therefore, the cytokinins auxin and ethephon were also applied for the haustorium induction of
P. aegyptiaca. DMBQ has been widely used as a HIF in hemi-parasitic plants and thus was tested for haustorium induction activity in
P. aegyptiaca. Root exudates were confirmed to have haustorium-inducing activity in this study, so the mixed root exudates of the two cultivars were used as positive controls to evaluate the haustorium activity of different chemical compounds. GR24 and sterilized water were treated separately as negative controls. Each chemical treatment was accompanied by the corresponding solvent as a control to evaluate whether the solvent had a negative effect on
P. aegyptiaca development. At least three biological replicates per treatment and 30
P. aegyptiaca seeds were measured per replicate.
2.4. Conditions for the Analysis of UPLC-MS/MS
Metabolite analysis was sequentially processed at Wuhan Metware Biotechnology Co., Ltd. (
www.metware.cn, accessed on 6 February 2022). The sample extracts were analyzed using an UPLC-ESI-MS/MS system (UPLC, SHIMADZU Nexera X2; MS, Applied Biosystems 4500 Q TRAP, Waltham, MA, USA).
The UPLC analytical conditions were as follows: UPLC: column, Agilent SB-C18 (1.8 µm, 2.1 mm × 100 mm); the mobile phase consisted of solvent A, pure water with 0.1% formic acid, and solvent B was acetonitrile with 0.1% formic acid. Sample measurements were performed with a gradient program that employed the starting conditions of 95% A and 5% B. Within 9 min, a linear gradient to 5% A and 95% B was programmed, and a composition of 5% A and 95% B was maintained for 1 min. Subsequently, a composition of 95% A and 5.0% B was adjusted within 1.1 min and kept for 2.9 min. The flow velocity was set as 0.35 mL per min; the column oven was set to 40 °C; the injection volume was 4 μL. The effluent was alternatively connected to an ESI-triple quadrupole-linear ion trap (QTRAP)-MS.
The ESI-Q TRAP-MS/MS analytical conditions were as follows: LIT and triple quadrupole (QQQ) scans were acquired on a triple quadrupole-linear ion trap mass spectrometer (Q TRAP), AB4500 Q TRAP UPLC/MS/MS System, equipped with an ESI Turbo Ion-Spray interface, operating in positive and negative ion mode and controlled by Analyst 1.6.3 software (AB Sciex, Framingham, MA, USA). The ESI source operation parameters were as follows: ion source, turbo spray; source temperature 550 °C; ion spray voltage (IS) 5500 V (positive ion mode)/−4500 V (negative ion mode); ion source gas I (GSI), gas II (GSII), and curtain gas (CUR) were set at 50, 60, and 25.0 psi, respectively; collision-activated dissociation (CAD) was high. Instrument tuning and mass calibration were performed with 10 and 100 μmol/L polypropylene glycol solutions in the QQQ and LIT modes, respectively. QQQ scans were acquired as MRM experiments with collision gas (nitrogen) set to medium. Declustering potential (DP) and collision energy (CE) for individual MRM transitions were performed with further DP and CE optimization. A specific set of MRM transitions was monitored for each period, according to the metabolites eluted within this period.
2.5. Qualitative and Quantitative Determination of Metabolites
Based on the self-built database MWDB (Metware Biotechnology Co., Ltd., Wuhan, China) and the public database of metabolite information, secondary mass spectrometry data were subjected to qualitative analysis. To avoid interference, isotope signals, duplicate signals of K+, Na+, and NH4+ ions, and duplicate signals of fragment ions derived from other relatively large molecules were excluded.
Metabolite quantification was performed using the MRM mode of the QQQ mass spectrometer. In the MRM mode, the quadrupole first screened the precursor ions (parent ions) of the target substances and removed the corresponding ions of other substances with different molecular weights to eliminate interference preliminarily. The precursor ions were fragmented via induced ionization in the collision chamber to form many fragment ions, which were then filtered through QQQ to select single-fragment ions with the desired characteristics while eliminating interference from non-target ions, leading to the increased precision and repeatability of the quantification results. After the metabolite mass spectrometry data were obtained for the different samples, all mass spectrum peaks were subjected to area integration, and the mass spectra peaks of the same metabolite in the different samples were integrated and corrected using MultiaQuant software (Sciex, Framingham, MA, USA). The area of each peak represented the relative content of the corresponding substance.
2.6. Data Analysis
Multivariate statistical analysis methods, namely, principal component analysis (PCA) and orthogonal partial least squares-discriminant analysis (OPLS-DA), were used to process the metabolic data. PCA was first carried out on all samples, including quality control samples (QC), to elucidate the total metabolic differences and degree of variation among the samples. Fold change (FC) and variable importance in projection (VIP) values of the orthogonal partial least squares discriminant analysis (OPLS-DA) model were used to screen differential metabolites. The accumulation pattern of metabolites among different samples was analyzed by cluster analysis with R software version 2.8.0, (
www.r-project.org/, accessed on 25 February 2022). The Kyoto Encyclopedia of Genes and Genomes (KEGG) database was used to annotate and enrich differential metabolites.
The obtained experimental data were subjected to a one-way analysis of variance (ANOVA) using IBM SPSS Statistics V22. The mean values of the tested treatments were compared using the least significant range (L.S.R.) according to Duncan’s multiple range test at a p = 0.05 level of probability. Figures were drawn using Origin 2021 and Rstudio Version 1.3.1093 (Boston, MA, USA).
4. Discussion
Phelipanche aegyptiaca is one of the most destructive root parasitic plants of Orobanchaceae, which has significant impacts on crop yields worldwide [
27]. However, as an obligate parasite,
P. aegyptiaca has brought many difficulties to the control work. Effective control methods have become the subject of constant exploration.
Reducing the soil seed bank by inducing suicidal germination of the parasite seeds is an effective method to control the parasite, which has prompted many researchers to study chemicals regulating the germination of parasites. Strigolactones (SLs) are the most potent stimulants and are distributed widely in the plant kingdom [
28]. MP3, MP16, and Nijmegen-1, three types of SL analog, the germination induction effect on
Striga hermonthica is comparable to the standard SL analog GR24, which can be used to improve suicide germination technology [
29].
Brassica napus exudes only small amounts of SLs, while another active stimulant 2-phenylethyl isothiocyanate (ITC), secreted from root exudates of oilseed rape induced the germination of
P. ramosa [
30]. Besides, the haustorium formation is another key step in the life cycle of parasites after seed germination. It is also significant to study the haustorium formation for understanding the interaction mechanism between the parasite and the host and applying it in the control of the parasite.
At present, research on haustoria mostly focuses on semi-parasitic plants, and the haustorium-inducing factors in holoparasitic plants and the mechanisms of parasitic plants responding to host roots or host root-derived molecules are largely unknown.
Cucumis melo is a major host of
P. aegyptiaca. The resistant cultivar KR1326 and susceptible cultivar K1076 were selected to collect root exudates for seed germination and haustorium induction of
P. aegyptiaca. Obligate parasite
P. aegyptiaca can parasitize the whole life cycle of the
C. melo, including seedling, flowering, and fruiting, and even after the death of the host,
P. aegyptiaca can still survive for a short time [
1]. In contrast, melon at the flowering stage will absorb more nutrients for fruiting, they would grow vigorously and secrete more root exudates. Therefore,
C. melo at the flowering stage was selected to collect root exudates. In the present study,
C. melo root exudate extracts stably induced
P. aegyptiaca seed germination and haustorium formation in vitro (
Figure 1), corresponding to previous studies where seeds of parasitic plants from the Orobanchaceae family responded to host root extracts [
14,
15]. The swelling of the root radicle tip, shorter roots, and outgrowth of epidermal cells were observed in
P. aegyptiaca after treatment with root exudates or specific chemicals, which were typical haustorium features, which is similar to the description of the characteristics of
Phelipanche spp. haustoria in previous studies [
7,
14].
Haustorium development in Orobanchaceae is not a spontaneous process; it must be stimulated by chemicals secreted by the host roots to initiate the haustorium [
1,
9]. Almost 50% of
P. aegyptiaca seeds had induced haustoria after treatment with root exudates, and no haustoria were observed in the control (
Figure 4). The results showed that there are special allelochemicals in
C. melo root exudates that can trigger cell differentiation at the apex of the
P. aegyptiaca radicle and then form a multicellular early haustorial structure.
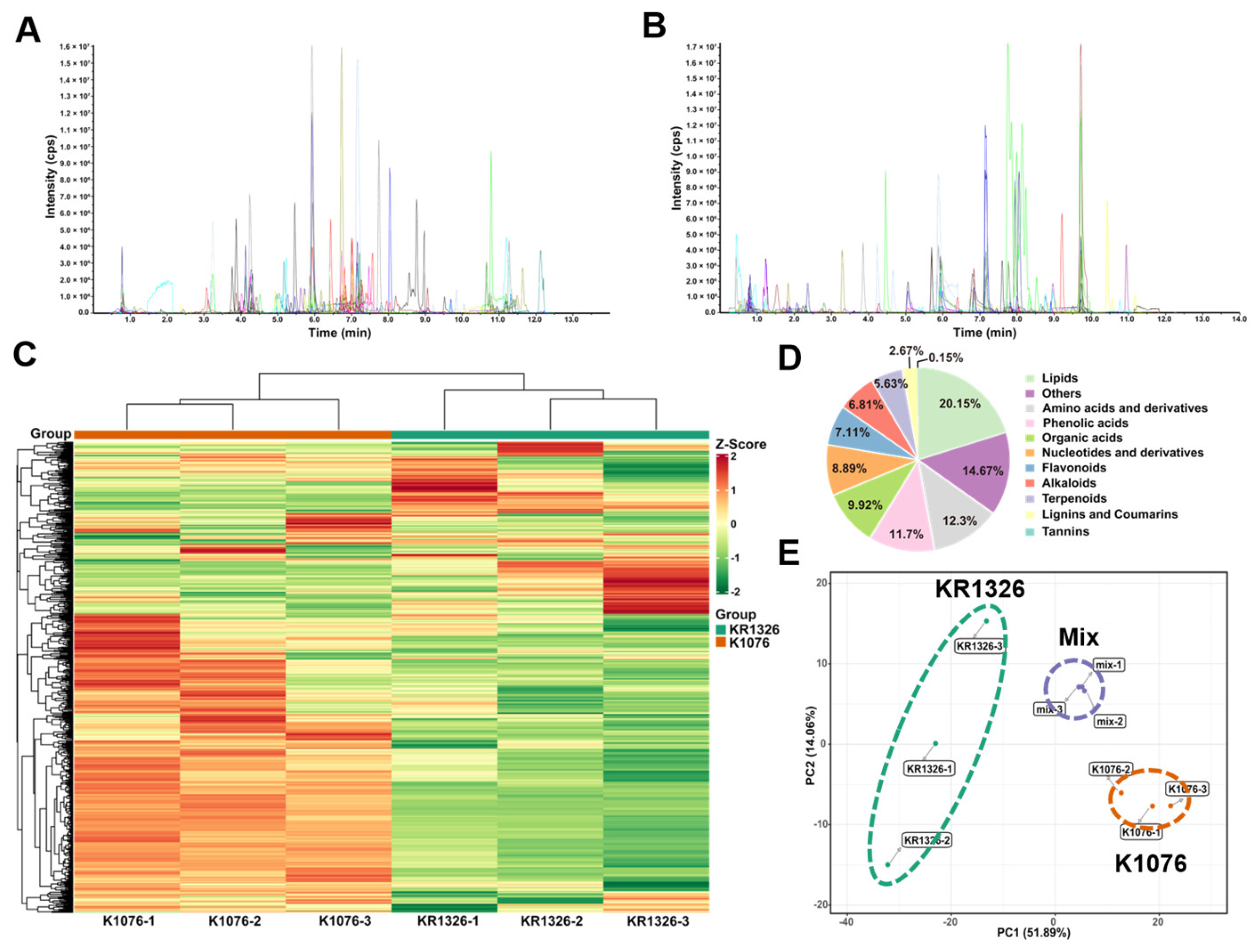
To search for allelochemicals related to haustorium induction in root exudates, we compared the induction activity of resistant and susceptible
C. melo root exudates. Through comparison, we found differences in induction activity between the resistant and susceptible cultivars (
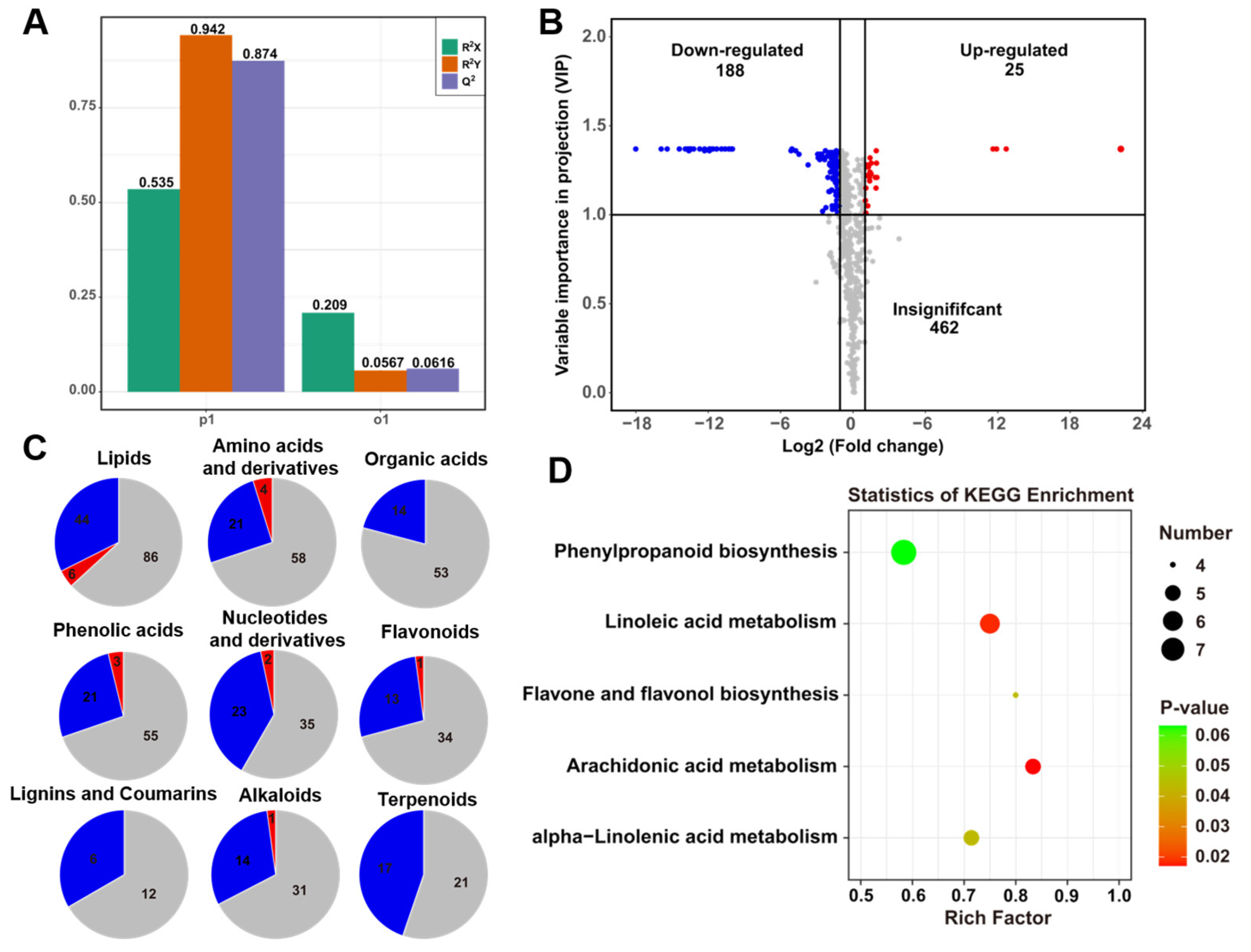
Figure 2C,E). Therefore, widely targeted metabonomics based on UPLC MS/MS technology were used to analyze and identify metabolites in root exudates. Notably, 88% of differential metabolites differentially accumulated in K1076 rather than in KR1326 root exudates (
Figure 3B), which was closely related to the physiological differences between the two cultivars. The developed size of the root system and plant height of K1076 were significantly higher than KR1326; thus, the K1076 roots secreted more metabolites (
Figure S1). The KEGG enrichment analysis confirmed this result, differential metabolites were significantly enriched in arachidonic acid metabolism, linoleic acid metabolism, and alpha-linolenic acid metabolism pathways (
Figure 3D); the metabolites in these pathways are mainly important energy storage substances in plants [
31], which were all higher in K1076 (
Figure S3). In addition, p-coumaric acid, p-coumaraldehyde, p-coumaric alcohol, and cinnamic acid were significantly enriched in the phenylpropanoid biosynthesis pathway. They are all precursors of lignin biosynthesis, and lignin has been reported as an important source of HIFs in semi-parasitic plants [
15]. Therefore, we speculated that the phenylpropanoid biosynthesis pathway is closely related to the HIFs of the obligate parasitic plant
P. aegyptiaca. A previous study indicated that quercetin could inhibit the radicle growth of
P. ramosa and that 0.05–0.1 mM quercetin and its derivatives had haustorium induction activity [
32]. In the present study, 3-O-methyl quercetin was significantly enriched in the flavone and flavonol biosynthesis pathways. Therefore, there are potential HIFs of
P. aegyptiaca in the flavone and flavonol biosynthesis pathways. These results suggest that these differential metabolites contain HIFs and that different types and contents of HIFs in host exudates result in differences in haustorial induction activity.
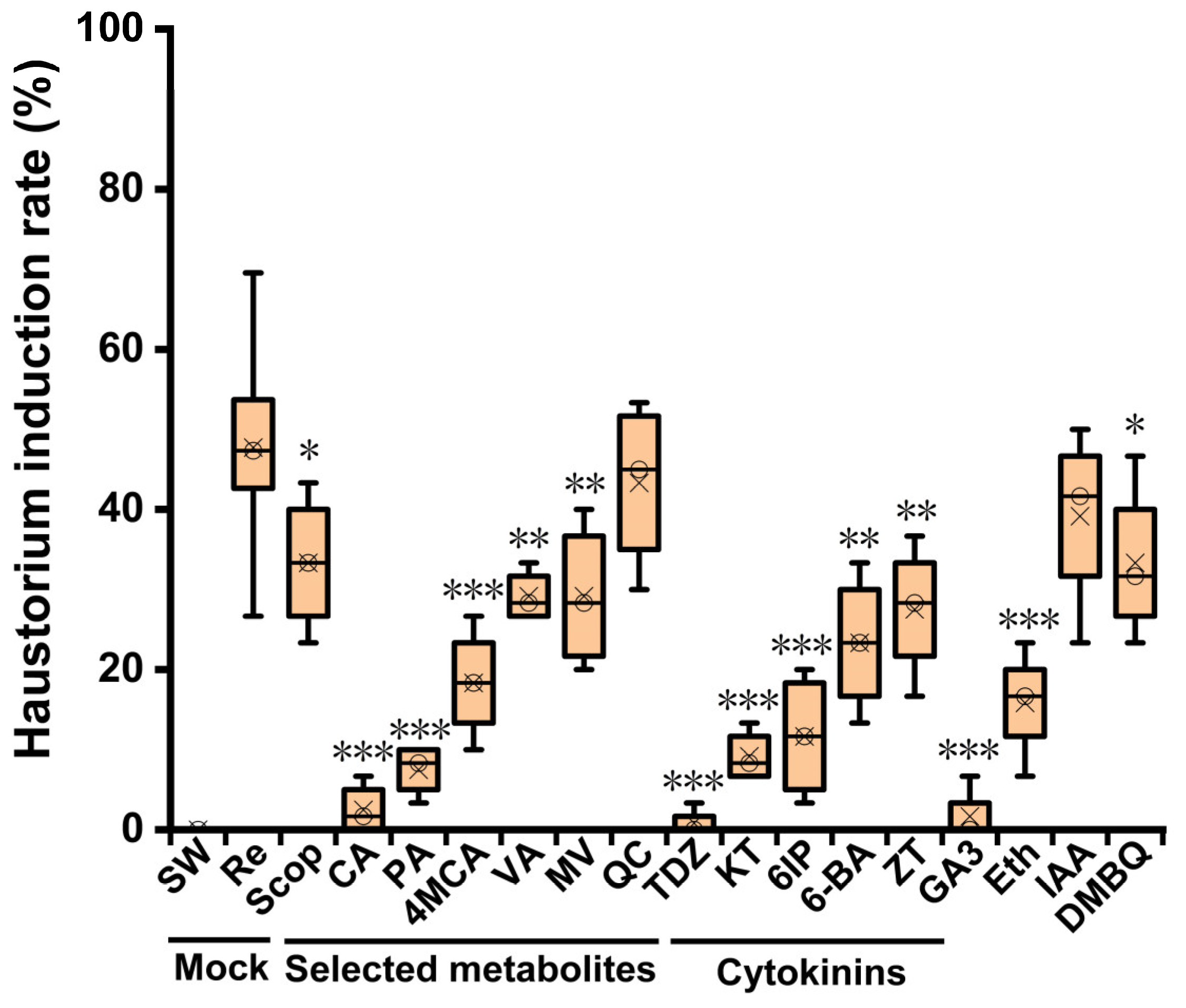
Among the different compounds tested in this study, QC and IAA induced haustorium formation with maximum activity similar to
C. melo root exudate treatment (
Figure 4). Quercetin is a flavonoid compound that widely exists in plants, which can adjust polar auxin transport and regulate the activity of various antioxidant enzymes [
33]. A recent study showed that quercetin can reduce
P. ramosa radicle growth and induce haustoria at a concentration range of 0.1–0.05 mM [
32]. This is consistent with the results that haustorium formation was observed under QC treatment at 10
−3–10
−6 mol/L in the present study. 3-O-Methylquercetin, a derivative of quercetin, was significantly enriched in K1076 root exudates. This may be associated with its higher haustorium induction rate than with KR1326. Auxin signaling and transport genes are upregulated in the haustorial tissues of some parasitic plants [
34,
35,
36]. Exogenous treatment with IAA has been shown to trigger haustorial hair formation in
T. versicolor [
37]. This evidence demonstrates that auxin plays an important role in regulating haustorium development in parasitic plants. Similarly, in the present study, exogenous application of 10
−4 mol/L IAA treatment displayed relatively high haustorium induction activity, whereas low haustorium induction rates were shown at lower or higher concentrations. A transcriptomic analysis of
P. aegyptiaca showed that the levels of plant hormone GA3 and ethylene increased after germination stimulation [
27]. However, we noticed that the exogenous application of ethylene and GA3 almost did not induce the haustorium formation (
Figure S2O,P), This does not mean that GA3 and ethylene do not participate in the regulation of the formation of
P. aegyptiaca. On the contrary, GA3 and ethylene, and other plant hormones may change endogenously to regulate the growth of
P. aegyptiaca before parasitism, which is worthy of further study.
Scop and DMBQ also showed relatively high haustorial induction activity. DMBQ, the first HIF identified directly from host root extracts, is a common product of lignin biosynthesis and degradation released by host cells upon host peroxidase activity triggered by hydrogen peroxide production at the tip of
Striga radicles [
9]. At present, it has been widely used in the haustorium induction of semi-parasitic plants. However, the effect of haustoria on obligate parasitic plants has long been uncertain. The results in the present study indicated that
P. aegyptiaca haustoria can be induced by DMBQ under appropriate conditions, and there is haustorium induction activity at a concentration of 10
−4–10
−6 mol/L (
Figure S2M). Other previous studies have confirmed this result. BQ and DMBQ can induce haustorium formation in
Orobanche cumana,
Orobanche minor, and
Phelipanche ramose [
32,
38]. Those results demonstrated that DMBQ is also an HIF of obligate-parasitic plants, but the haustorium induction activity is far lower than that of semi-parasitic plants; thus, higher haustorium induction requires further optimization of conditions.
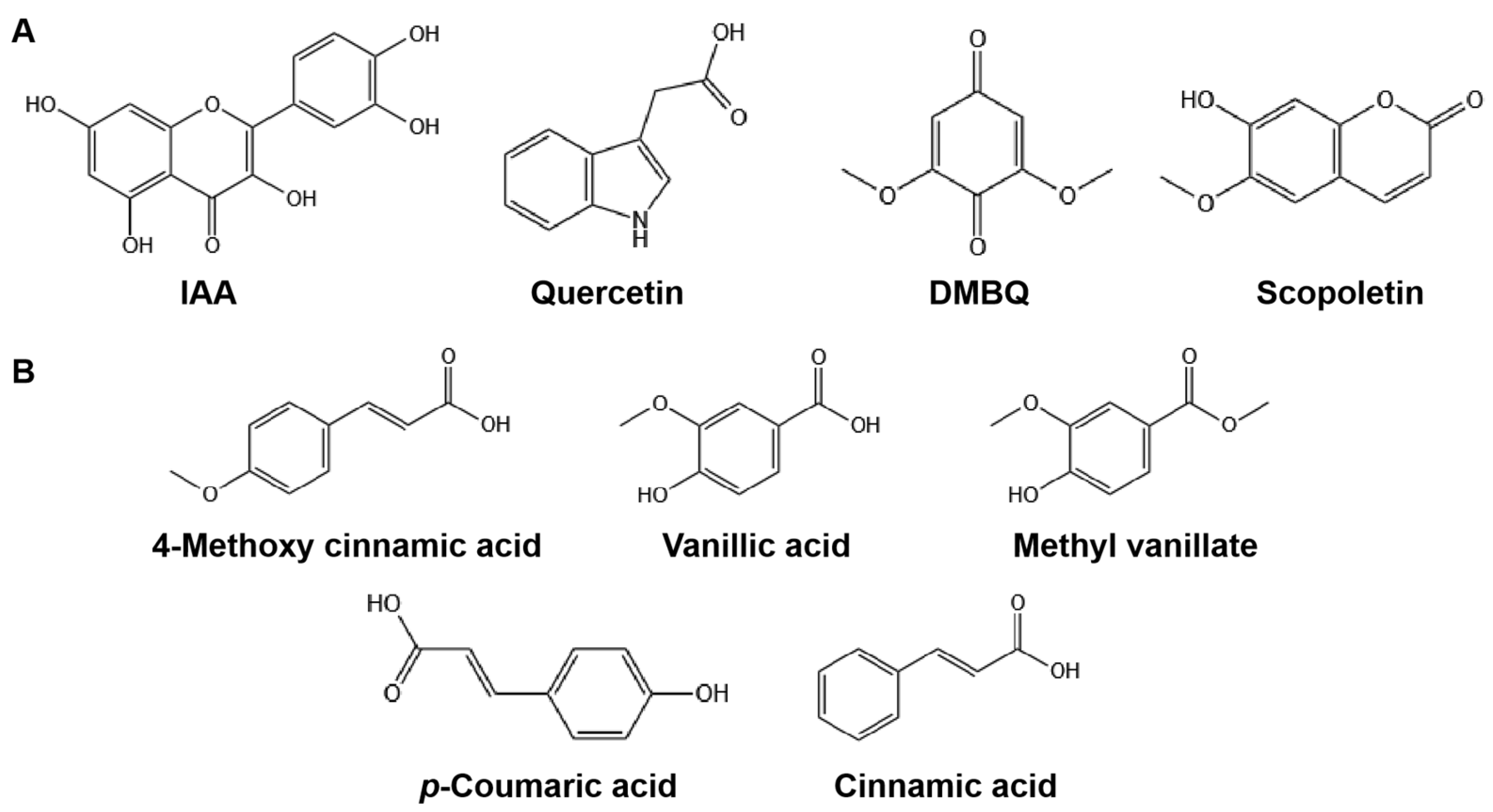
A previous study showed that plant lignin is an important source of HIFs in parasitic plants and that at least one methoxyl group adjacent to the 4-hydroxyl group in phenolics or the 1-oxo group in quinone is an important structural feature of lignin-associated HIFs [
15]. PA and CA are important intermediates in lignin biosynthesis, and 4MCA, VA, and MV are their structural analogs [
39,
40], which are all differential metabolites identified from
C. melo root exudates and differentially accumulated in K1076 (
Table S3). Interestingly, among these metabolites, 4MCA, VA, and MV displayed certain haustorium induction activity, and PA and CA were almost inactive. Compared with material structures, 4MCA, VA, and MV have at least one methoxyl group on the benzene ring, whereas PA and CA do not (
Figure 5). These results confirm previous results and suggest that lignin-related compounds are also an important source of HIFs for obligate parasitic plants
P. aegyptiaca, in which the methoxyl group is an important factor affecting its haustorium induction activity.
In addition, the allelochemicals secreted by the host are necessary for the germination and the formation of the haustorium of parasitic plants. While some hosts with pre-attachment resistance can make themselves “invisible” to parasitic plants by regulating metabolism and changing the type and content of allelochemicals secreted to the outside, or even directly secrete toxic chemicals to inhibit the germination and haustorium formation of parasitic plants, thus avoiding being parasitized [
41]. Therefore, studying the metabolic differences between resistant and susceptible hosts is helpful to reveal the mechanism of host resistance and control of parasitic plants.
At present, no haustorium inhibitors were found in this study, while previous studies have shown that exogenous treatment with the auxin activity inhibitor p-chlorophenoxyisobutyric acid (PCIB) or the auxin transport inhibitor 1-N-naphthylphthalamic acid (NPA) dramatically reduces the frequency of infection of Arabidopsis thaliana by
P. aegyptiaca [
42]. Tetrafluorobenzoquinone (TFBQ) and Cyclopropyl-p-benzoquinone (CPBQ), the structural analogs of DMBQ, were applied to treat the
Triphysaria versicolor seeds and reduce the formation of haustorium in the presence of the DMBQ [
43]. The results in these studies indicate that the haustorium inhibitors of the parasitic plant
P. aegyptiaca may exist, which requires testing more compounds.
In the present study, quercetin, IAA, and some phenolic acids were screened from the root exudates of C. melo and played important roles in inducing haustorium formation in P. aegyptiaca. This sheds new light on the mechanism, whereby the host plant first reduces the biosynthesis and secretion of HIFs by applying exogenous inhibitors related to HIFs, and then reduces haustorium formation to avoid the parasitism of P. aegyptiaca, which provides effective control methods against parasitic weeds.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




