
Functional Analysis of ScABI3 from Syntrichia caninervis Mitt. in Medicago sativa L.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Construction of Plant Expression Vector and Transformation
2.3. PCR Authentication and qRT-PCR Analysis
2.4. Determination of Endogenous ABA Content
2.5. Measurement of Leaf Area
2.6. Measurement of the Leaf-Stem Ratio
2.7. Determination of Photosynthetic Indicators
2.8. Determination of Crude Protein and Fat Contents
2.9. Statistical Analysis
3. Results
3.1. Regeneration of Transgenic Plants
3.2. PCR and qRT-PCR Analysis of the Transgenics
3.3. Endogenous ABA Content in Transgenic Alfalfa
3.4. Phenotype Analysis
3.5. Leaf Gas Exchange Parameters of Transgenic Alfalfa
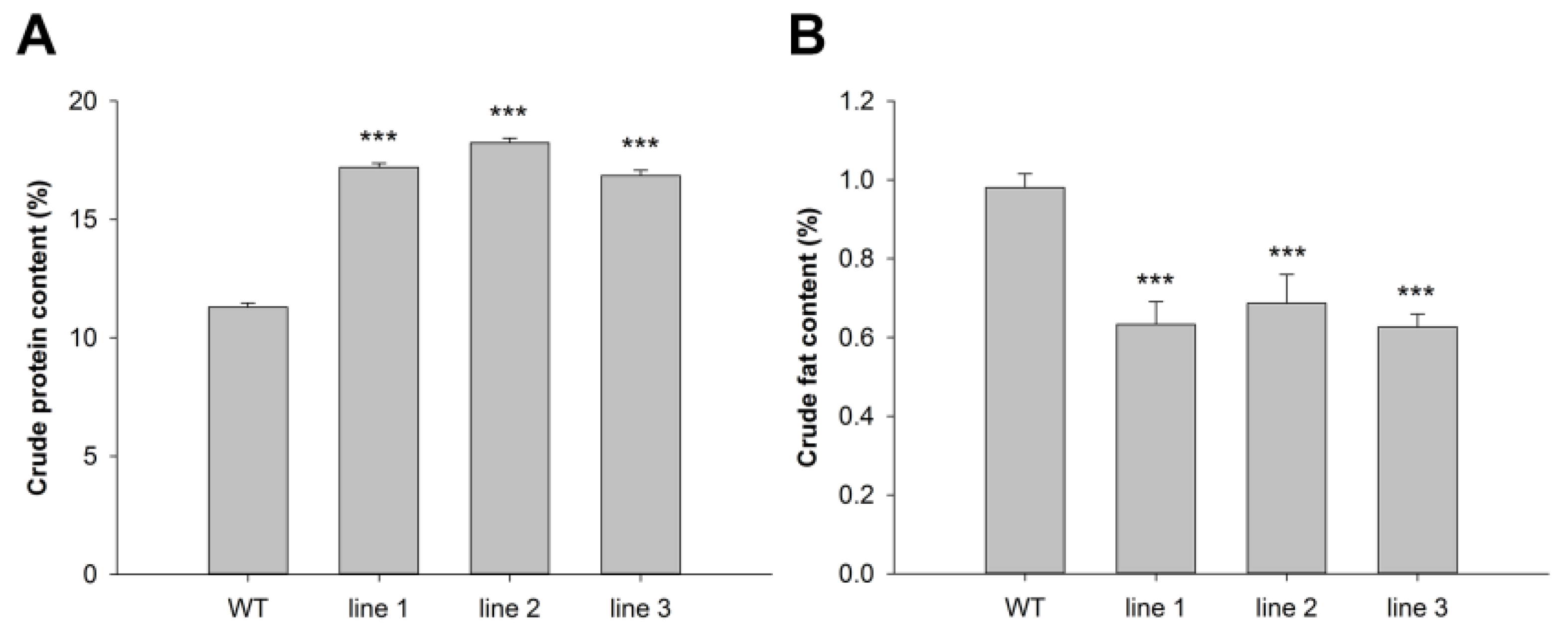
3.6. Crude Protein and Fat Contents of Transgenic Alfalfa
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Radović, J.; Sokolović, D.; Marković, J. Alfalfa-most important perennial forage legume in animal husbandry. Biotechnol. Anim. Husb. 2009, 25, 465–475. [Google Scholar] [CrossRef]
- Chen, H.; Zeng, Y.; Yang, Y.; Huang, L.; Tang, B.; Zhang, H.; Hao, F.; Liu, W.; Li, Y.; Liu, Y.; et al. Allele-aware chromosome-level genome assembly and efficient transgene-free genome editing for the autotetraploid cultivated alfalfa. Nat. Commun. 2020, 11, 2494. [Google Scholar] [CrossRef] [PubMed]
- Wolabu, T.W.; Cong, L.; Park, J.-J.; Bao, Q.; Chen, M.; Sun, J.; Xu, B.; Ge, Y.; Chai, M.; Liu, Z.; et al. Development of a highly efficient multiplex genome editing system in outcrossing tetraploid alfalfa (Medicago sativa). Front. Plant Sci. 2020, 11, 1063. [Google Scholar] [CrossRef]
- Li, X.; Brummer, E.C. Applied genetics and genomics in alfalfa breeding. Agronomy 2012, 2, 40–61. [Google Scholar] [CrossRef]
- Matthews, C.; Arshad, M.; Hannoufa, A. Alfalfa response to heat stress is modulated by microRNA. Physiol. Plant. 2019, 165, 830–842. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Austin, R.S.; Amyot, L.; Hannoufa, A. Comparative transcriptome investigation of global gene expression changes caused by miR156 overexpression in Medicago sativa. BMC Genom. 2016, 17, 658. [Google Scholar] [CrossRef]
- Song, Y.; Lv, J.; Ma, Z.; Dong, W. The mechanism of alfalfa (Medicago sativa L.) response to abiotic stress. Plant Growth Regul. 2019, 89, 239–249. [Google Scholar] [CrossRef]
- Zhang, J.; Duan, Z.; Zhang, D.; Zhang, J.; Di, H.; Wu, F.; Wang, Y. Co-transforming bar and CsLEA enhanced tolerance to drought and salt stress in transgenic alfalfa (Medicago sativa L.). Biochem. Biophys. Res. Commun. 2016, 472, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.; Bao, A.-K.; Kumar, T.; Pan, Y.-Q.; Bao, Z.; Wang, F.W.a.S.-M. Assessment of stress tolerance, productivity, and forage quality in T1 transgenic alfalfa co-overexpressing ZxNHX and ZxVP1-1 from Zygophyllum xanthoxylum. Front. Plant. Sci. 2016, 7, 1598. [Google Scholar] [CrossRef]
- Deak, M.; Kiss, G.B.; Koncz, C.; Dudits, D. Transformation of Medicago by Agrobacterium mediated gene transfer. Plant Cell Rep. 1986, 5, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Delmas, F.; Sankaranarayanan, S.; Deb, S.; Widdup, E.; Bournonville, C.; Bollier, N.; Northey, J.G.B.; McCourt, P.; Samuel, M.A. ABI3 controls embryo degreening through Mendel’s I locus. Proc. Natl. Acad. Sci. USA 2013, 110, 3888–3894. [Google Scholar] [CrossRef] [PubMed]
- Bradyz, S.M.; Sarkarz, S.F.; Bonettay, D.; McCourt, P. The ABSCISIC ACID INSENSITIVE3(ABI3) gene is modulated by farnesylation and is involved in auxin signaling and lateral root development in Arabidopsis. Plant J. 2003, 34, 67–75. [Google Scholar] [CrossRef]
- Tian, R.; Wang, F.; Zheng, Q.; Niza, V.M.A.G.E.; Downie, A.B.; Perry, S.E. Direct and indirect targets of the arabidopsis seed transcription factor ABSCISIC ACID INSENSITIVE3. Plant J. 2020, 103, 1679–1694. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, A.; Cho, S.H.; Marella, H.; Sakata, Y.; Perroud, P.-F.; Pan, A.; Quatrano, R.S. Role of ABA and ABI3 in Desiccation Tolerance. Science 2010, 327, 546. [Google Scholar] [CrossRef]
- Tamminen, I.; Makela, P.; Heino, P.; Palva, E.T. Ectopic expression of ABI3 gene enhances freezing tolerance in response to abscisic acid and low temperature in Arabidopsis thaliana. Plant J. 2001, 25, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Gampala, S.S.L.; Ritchie, G.L.; Payton, P.; Burke, J.J.; Rock, C.D. Related to ABA-Insensitive3 (ABI3)/Viviparous1 and AtABI5 transcription factor coexpression in cotton enhances drought stress adaptation. Plant Biotechnol. J. 2014, 12, 578–589. [Google Scholar] [CrossRef]
- Rohde, A.; Prinsen, E.; Rycke, R.D.; Engler, G.; Montagu, M.V.; Boerjan, W. PtABI3 impinges on the growth and differentiation of embryonic leaves during bud set in poplar. Plant Cell 2002, 14, 1885–1901. [Google Scholar] [CrossRef]
- Wang, S.; Guo, T.; Shen, Y.; Wang, Z.; Kang, J.; Zhang, J.; Yi, F.; Yang, Q.; Long, R. Overexpression of MtRAV3 enhances osmotic and salt tolerance and inhibits growth of Medicago truncatula. Plant Physiol. Biochem. 2021, 163, 154–165. [Google Scholar] [CrossRef]
- Pan, Z.; Pitt, W.G.; Zhang, Y.; Wu, N.; Tao, Y.; Truscott, T.T. The upside-down water collection system of Syntrichia caninervis. Nat. Plants 2016, 2, 16076. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiujin, L.; Kechuang, Z.; Daoyuan, Z.; Kaiyun, G. An ABSCISIC ACID INSENSITIVE3-like gene from the desert moss Syntrichia caninervis confers abiotic stress tolerance and reduces ABA sensitivity. Plant Cell Tissue Org. 2018, 133, 417–435. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Mutailifu, A.; Zhang, D. Heterologous Overexpression of Desiccation-tolerance Moss ScABI3 Gene Changes Stomatal Phenotype and Improves Drought Resistance in Transgenic Arabidopsis. Chin. Bull. Bot. 2021, 56, 414–421. [Google Scholar] [CrossRef]
- Zhang, Y.; Mutailifu, A.; Zhang, Y.; Yang, H.; Zhang, D. Detection of abscisic acid and relative transcript abundance in Syntrichia caninervis Mitt. J. Bryol. 2021, 43, 376–383. [Google Scholar] [CrossRef]
- Wang, B.; Ma, M.; Lu, H.; Meng, Q.; Li, G.; Yang, X. Photosynthesis, sucrose metabolism, and starch accumulation in two NILs of winter wheat. Photosynth. Res. 2015, 126, 363–373. [Google Scholar] [CrossRef]
- Stedt, K.; Trigo, J.P.; Steinhagen, S.; Nylund, G.M.; Forghani, B.; Pavia, H.; Undeland, I. Cultivation of seaweeds in food production process waters: Evaluation of growth and crude protein content. Algal Res. 2022, 63, 102647. [Google Scholar] [CrossRef]
- Wong, K.H.; Cheung, P.C.K. Nutritional evaluation of some subtropical red and green seaweeds. Food Chem. 2000, 71, 475–482. [Google Scholar] [CrossRef]
- Hefferon, K.L. Nutritionally enhanced food crops; Progress and perspectives. Int. J. Mol. Sci. 2015, 16, 3895–3914. [Google Scholar] [CrossRef]
- Bao, A.-K.; Du, B.-Q.; Touil, L.; Kang, P.; Wang, Q.-L.; Wang, S.-M. Co-expression of tonoplast Cation/H+antiporter and H+-pyrophosphatase from xerophyte Zygophyllum xanthoxylum improves alfalfa plant growth under salinity, drought and field conditions. Plant Biotechnol. J. 2016, 14, 964–975. [Google Scholar] [CrossRef]
- Aung, B.; Gruber, M.Y.; Amyot, L.; Omari, K.; Bertrand, A.; Hannoufa, A. MicroRNA156 as a promising tool for alfalfa improvement. Plant Biotechnol. J. 2015, 13, 779–790. [Google Scholar] [CrossRef]
- Aung, B.; Gao, R.; Gruber, M.Y.; Yuan, Z.-C.; Sumarah, M.; Hannoufa, A. MsmiR156 affects global gene expression and promotes root regenerative capacity and nitrogen fixation activity in alfalfa. Transgenic Res. 2017, 26, 541–557. [Google Scholar] [CrossRef]
- Cen, H.; Wang, T.; Liu, H.; Wang, H.; Tian, D.; Li, X.; Cui, X.; Guan, C.; Zang, H.; Li, M.; et al. Overexpression of MsASMT1 promotes plant growth and decreases flavonoids biosynthesis in transgenic alfalfa (Medicago sativa L.). Front. Plant Sci. 2020, 11, 489. [Google Scholar] [CrossRef]
- Confalonieri, M.; Carelli, M.; Tava, A.; Borrelli, L. Overexpression of MtTdp2α (tyrosyl-DNA phosphodiesterase 2) gene confers salt tolerance in transgenic Medicago truncatula. Plant Cell Tissue Org. 2019, 137, 157–172. [Google Scholar] [CrossRef]
- Shen, C.; Du, H.; Chen, Z.; Lu, H.; Zhu, F.; Chen, H.; Meng, X.; Liu, Q.; Liu, P.; Zheng, L.; et al. The chromosome-level genome sequence of the autotetraploid alfalfa and resequencing of core germplasms provide genomic resources for alfalfa research. Mol. Plant 2020, 13, 1250–1261. [Google Scholar] [CrossRef]
- Li, X.-P.; Xu, Q.-Q.; Liao, W.-B.; Ma, Z.-J.; Xu, X.-T.; Wang, M.; Ren, P.-J.; Niu, L.J.; Jin, X.; Zhu, Y.-C. Hydrogen peroxide is involved in abscisic acid-induced adventitious rooting in cucumber (Cucumis sativus L.) under drought stress. J. Plant Biol. 2016, 59, 536–548. [Google Scholar] [CrossRef]
- Truong, H.A.; Lee, S.; Trinh, C.S.; Lee, W.J.; Chung, E.-H.; Hong, S.-W.; Lee, H. Overexpression of the HDA15 gene confers resistance to salt stress by the induction of NCED3, an ABA biosynthesis enzyme. Front. Plant Sci. 2021, 12, 631. [Google Scholar] [CrossRef]
- Qiu, D.; Hu, W.; Zhou, Y.; Xiao, J.; Hu, R.; Wei, Q.; Zhang, Y.; Feng, J.; Sun, F.; Sun, J.; et al. TaASR1-D confers abiotic stress resistance by affecting ROS accumulation and ABA signalling in transgenic wheat. Plant Biotechnol. J. 2021, 19, 1588. [Google Scholar] [CrossRef]
- Ma, H.; Liu, C.; Li, Z.; Ran, Q.; Xie, G.; Wang, B.; Fang, S.; Chu, J.; Zhang, J. ZmbZIP4 contributes to stress resistance in maize by regulating ABA synthesis and root development. Plant Physiol. 2018, 178, 753–770. [Google Scholar] [CrossRef]
- Ijaz, R.; Ejaz, J.; Gao, S.; Liu, T.; Imtiaz, M.; Ye, Z.; Wang, T. Overexpression of annexin gene AnnSp2, enhances drought and salt tolerance through modulation of ABA synthesis and scavenging ROS in tomato. Sci Rep. 2017, 7, 12087. [Google Scholar] [CrossRef]
- Wasilewska, A.; Vlad, F.; Sirichandra, C.; Redko, Y.; Jammes, F.; Valon, C.; Frey, N.F.d.; Leung, J. An update on abscisic acid signaling in plants and more…. Mol. Plant 2008, 1, 198–217. [Google Scholar] [CrossRef]
- Lopez-Molina, L.; Mongrand, S.; McLachlin, D.T.; Chait, B.T.; Chua, N.-H. ABI5 acts downstream of ABI3 to execute an ABA-dependent growth arrest during germination. Plant J. 2002, 32, 317–328. [Google Scholar] [CrossRef]
- Park, J.; Lee, N.; Kim, W.; Lim, S.; Choi, G. ABI3 and PIL5 collaboratively activate the expression of SOMNUS by directly binding to its promoter in imbibed Arabidopsis seeds. Plant Cell 2011, 23, 1404–1415. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Peng, C.; Liu, H.; Tang, M.; Yang, H.; Li, X.; Liu, J.; Sun, X.; Wang, X.; Xu, J.; et al. Genome-wide association study reveals candidate genes for control of plant height, branch initiation height and branch number in rapeseed (Brassica napus L.). Front. Plant Sci. 2017, 8, 1246. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xue, Q.; Zhang, Z.; Du, J.; Yu, D.; Huang, F. GmMYB181, a soybean R2R3-MYB protein, increases branch number in transgenic Arabidopsis. Front. Plant Sci. 2018, 9, 1027. [Google Scholar] [CrossRef]
- Veronesi, F.; Brummer, E.C.; Huyghe, C. Fodder Crops and Amenity Grasses; Springer: New York, NY, USA, 2010; pp. 395–437. [Google Scholar]
- Wang, K.; Liu, Y.; Teng, F.; Cen, H.; Yan, J.; Lin, S.; Li, D.; Zhang, W. Heterogeneous expression of Osa-MIR156bc increases abiotic stress resistance and forage quality of alfalfa. Crop J. 2021, 9, 1135–1144. [Google Scholar] [CrossRef]
- Wang, B.; Li, Z.; Ran, Q.; Li, P.; Peng, Z.; Zhang, J. ZmNF-YB16 overexpression improves drought resistance and yield by enhancing photosynthesis and the antioxidant capacity of maize plants. Front. Plant Sci. 2018, 9, 709. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yarra, R.; Yang, Y.; Liu, Y.; Yang, M.; Cao, H. The oil palm R2R3-MYB subfamily genes EgMYB111 and EgMYB157 improve multiple abiotic stress tolerance in transgenic Arabidopsis plants. Plant Cell Rep. 2022, 41, 377–393. [Google Scholar] [CrossRef]
- Strbanovic, R.; Stanisavljevic, R.; Dukanovic, L.; Postic, D.; Markovic, J.; Gavrilovic, V.; Dolovac, N. Variability and correlation of yield and forage quality in alfalfa varieties of different origin. J. Agric. Sci. 2017, 23, 128–137. Available online: https://doi.org/dergipark.org.tr/en/pub/ankutbd/issue/56551/786556 (accessed on 1 January 2022).
- Musco, N.; Koura, I.B.; Tudisco, R.; Awadjihè, G.; Adjolohoun, S.; Cutrignelli, M.I.; Mollica, M.P.; Houinato, M.; Infascelli, F.; Calabrò, S. Nutritional characteristics of forage grown in south of Benin. Asian-Australasian J. Anim. Sci. 2016, 29, 51. [Google Scholar] [CrossRef]
- Jia, B.; Sun, M.; Sun, X.; Li, R.; Wang, Z.; Wu, J.; Wei, Z.; DuanMu, H.; Xiao, J.; Zhu, Y. Overexpression of GsGSTU13 and SCMRP in Medicago sativa confers increased salt–alkaline tolerance and methionine content. Physiol. Plant. 2016, 156, 176–189. [Google Scholar] [CrossRef]
- Avraham, T.; Badani, H.; Galili, S.; Amir, R. Enhanced levels of methionine and cysteine in transgenic alfalfa ( Medicago sativa L.) plants over-expressing the Arabidopsis cystathionine γ -synthase gene. Plant Biotechnol. J. 2005, 3, 71–79. [Google Scholar] [CrossRef]
- Wang, S.; Guo, T.; Wang, Z.; Kang, J.; Yang, Q.; Shen, Y.; Long, R. Expression of three related to ABI3/VP1 genes in Medicago truncatula caused increased stress resistance and branch increase in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 611. [Google Scholar] [CrossRef]
- Gugała, M.; Sikorska, A.; Zarzecka, K.; Findura, P.; Malaga-Toboła, U. Chemical composition of winter rape seeds depending on the biostimulators used. Agronomy 2019, 9, 716. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wang, C.; Huang, M.; Zhang, Y. Functional Analysis of ScABI3 from Syntrichia caninervis Mitt. in Medicago sativa L. Agronomy 2022, 12, 2238. https://doi.org/10.3390/agronomy12092238
Zhang Y, Wang C, Huang M, Zhang Y. Functional Analysis of ScABI3 from Syntrichia caninervis Mitt. in Medicago sativa L. Agronomy. 2022; 12(9):2238. https://doi.org/10.3390/agronomy12092238
Chicago/Turabian StyleZhang, Yi, Chun Wang, Mingqi Huang, and Yigong Zhang. 2022. "Functional Analysis of ScABI3 from Syntrichia caninervis Mitt. in Medicago sativa L." Agronomy 12, no. 9: 2238. https://doi.org/10.3390/agronomy12092238