
Molecular Basis of Resistance to Mesosulfuron-Methyl in a Black-Grass (Alopecurus myosuroides Huds.) Population from China
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Collection in the Fields
2.2. Whole-Plant Dose–Response Assay and the Effect of P450s Inhibitor Malathion on Mesosulfuron-Methyl Resistance
2.3. DNA Extraction and PCR Amplification
2.4. ALS Enzyme Activity Assay
2.5. Sensitivity to Other Herbicides with Various Modes of Action
2.6. Statistical Analyses
3. Results
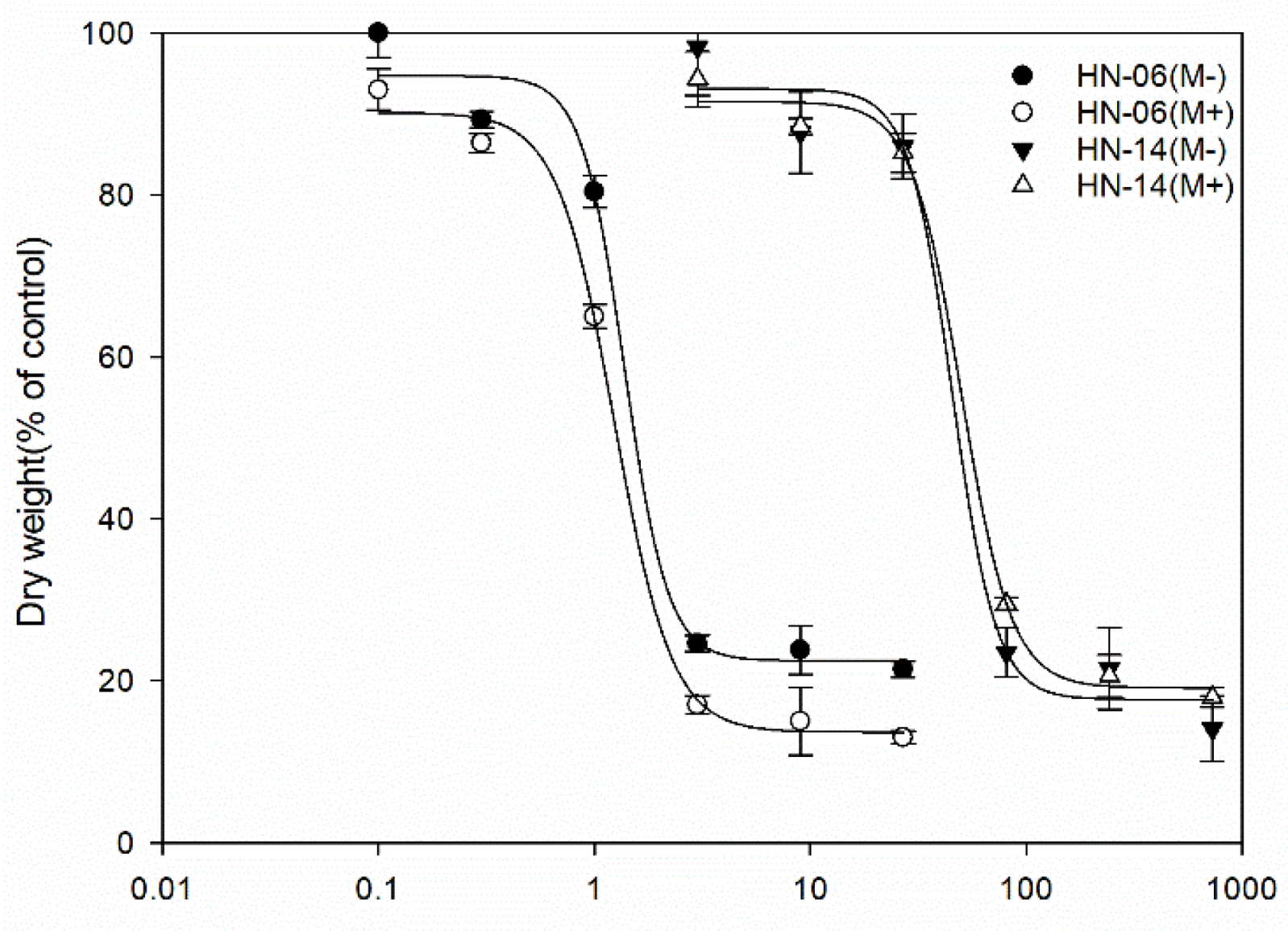
3.1. Sensitivity Bioassay to Mesosulfuron-Methyl
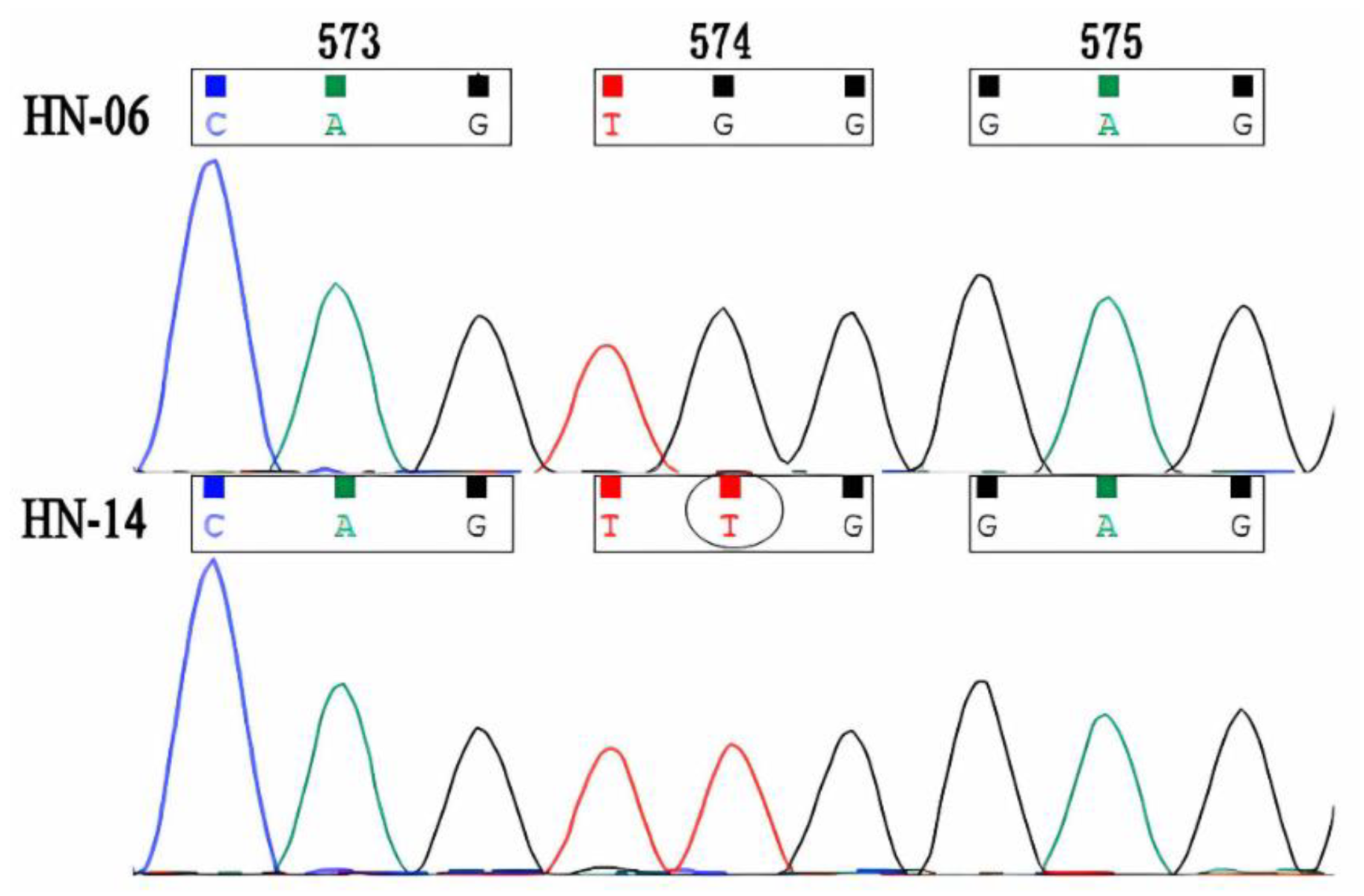
3.2. ALS Gene Sequencing and Sequences Analysis
3.3. In Vitro ALS Inhibition Assay
3.4. Cross Resistance to Other Herbicides
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cavan, G.; Cussans, J.; Moss, S.R. Modelling Different Cultivation and Herbicide Strategies for Their Effect on Herbicide Resistance in Alopecurus myosuroides. Weed Res. 2000, 40, 561–568. [Google Scholar] [CrossRef]
- Gao, X.; Sun, Z.; Li, M.; Fang, F.; Li, J.; Wu, J.; Lee, M. The Effect of Herbicide Application Timing and Dosage on Control of Alopecurus myosuroides in Winter Wheat. Acta Prataculturae Sin. 2016, 25, 172–179. [Google Scholar]
- Heap, J.; Knight, R. A Population of Ryegrass Tolerant to the Herbicide Diclofop-Methyl in South Australia. J. Aust. Inst. Agric. Sci. Aust. 1982, 48, 156–157. [Google Scholar]
- Heap International Survey of Herbicide Resistant Weeds. 2022. Available online: https://www.weedscience.org/ (accessed on 23 July 2022).
- McCourt, J.A.; Pang, S.S.; King-Scott, J.; Guddat, L.W.; Duggleby, R.G. Herbicide-Binding Sites Revealed in the Structure of Plant Acetohydroxyacid Synthase. Proc. Natl. Acad. Sci. USA 2006, 103, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Yuan, G.; Liu, W.; Bi, Y.; Du, L.; Zhang, C.; Li, Q.; Wang, J. Multiple Resistance to ACCase and AHAS-Inhibiting Herbicides in Shortawn Foxtail (Alopecurus aequalis Sobol.) from China. Pestic. Biochem. Physiol. 2015, 124, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yuan, G.; Du, L.; Guo, W.; Li, L.; Bi, Y.; Wang, J. A Novel Pro197Glu Substitution in Acetolactate Synthase (ALS) Confers Broad-Spectrum Resistance across ALS Inhibitors. Pestic. Biochem. Physiol. 2015, 117, 31–38. [Google Scholar] [CrossRef]
- Vandersteen, J.; Jaunard, D.; Mahy, G.; Bizoux, J.-P.; Monty, A.; Henriet, F.; Vancutsem, F.; Bodson, B. Dynamics of Black-Grass Populations Depending on the Sowing Time of Winter Wheat. In Proceedings of the Communications in Agricultural and Applied Biological Sciences, Ghent, Belgium, 24 May 2011; Volume 76. [Google Scholar]
- Lutman, P.J.W.; Moss, S.R.; Cook, S.; Welham, S.J. A Review of the Effects of Crop Agronomy on the Management of Alopecurus myosuroides. Weed Res. 2013, 53, 299–313. [Google Scholar] [CrossRef]
- Moss, S.R.; Tatnell, L.V.; Hull, R.; Clarke, J.H.; Wynn, S.; Marshall, R. Integrated Management of Herbicide Resistance. HGCA Proj. Rep. 2010, 466, xvii + 115. [Google Scholar]
- Chauvel, B.; Guillemin, J.-P.; Colbach, N. Evolution of a Herbicide-Resistant Population of Alopecurus myosuroides Huds. in a Long-Term Cropping System Experiment. Crop Prot. 2009, 28, 343–349. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in Action: Plants Resistant to Herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S. Metabolism-Based Herbicide Resistance and Cross-Resistance in Crop Weeds: A Threat to Herbicide Sustainability and Global Crop Production. Plant Physiol. 2014, 166, 1106–1118. [Google Scholar] [CrossRef]
- Délye, C.; Michel, S.; Bérard, A.; Chauvel, B.; Brunel, D.; Guillemin, J.; Dessaint, F.; Le Corre, V. Geographical Variation in Resistance to Acetyl-coenzyme A Carboxylase-inhibiting Herbicides across the Range of the Arable Weed Alopecurus myosuroides (Black-grass). New Phytol. 2010, 186, 1005–1017. [Google Scholar] [CrossRef]
- Kaundun, S.S.; Hutchings, S.-J.; Dale, R.P.; McIndoe, E. Role of a Novel I1781T Mutation and Other Mechanisms in Conferring Resistance to Acetyl-CoA Carboxylase Inhibiting Herbicides in a Black-Grass Population. PLoS ONE 2013, 8, e69568. [Google Scholar] [CrossRef] [Green Version]
- Petit, C.; Bay, G.; Pernin, F.; Délye, C. Prevalence of Cross- or Multiple Resistance to the Acetyl-Coenzyme A Carboxylase Inhibitors Fenoxaprop, Clodinafop and Pinoxaden in Black-Grass (Alopecurus myosuroides Huds.) in France. Pest Manag. Sci. 2010, 66, 168–177. [Google Scholar] [CrossRef]
- Adamczewski, K.; Kierzek, R.; Matysiak, K. Multiple Resistance to Acetolactate Synthase (ALS)- and Acetyl-Coenzyme A Carboxylase (ACCase)-Inhibiting Herbicides in Black-Grass (Alopecurus myosuroides Huds.) Populations from Poland. J. Plant Prot. Res. 2016, 56, 402–410. [Google Scholar] [CrossRef]
- Alwarnaidu Vijayarajan, V.B.; Forristal, P.D.; Cook, S.K.; Schilder, D.; Staples, J.; Hennessy, M.; Barth, S. First Detection and Characterization of Cross- and Multiple Resistance to Acetyl-CoA Carboxylase (ACCase)- and Acetolactate Synthase (ALS)-Inhibiting Herbicides in Black-Grass (Alopecurus myosuroides) and Italian Ryegrass (Lolium Multiflorum) Populations from Ireland. Agriculture 2021, 11, 1272. [Google Scholar] [CrossRef]
- Keshtkar, E.; Mathiassen, S.K.; Moss, S.R.; Kudsk, P. Resistance Profile of Herbicide-Resistant Alopecurus myosuroides (Black-Grass) Populations in Denmark. Crop Prot. 2015, 69, 83–89. [Google Scholar] [CrossRef]
- Bi, Y.; Liu, W.; Guo, W.; Li, L.; Yuan, G.; Du, L.; Wang, J. Molecular Basis of Multiple Resistance to ACCase- and ALS-Inhibiting Herbicides in Alopecurus japonicus from China. Pestic. Biochem. Physiol. 2016, 126, 22–27. [Google Scholar] [CrossRef]
- Guo, W.; Lv, L.; Zhang, L.; Li, Q.; Wu, C.; Lu, X.; Liu, W.; Wang, J. Herbicides Cross Resistance of a Multiple Resistant Short-Awn Foxtail (Alopecurus aequalis Sobol.) Population in Wheat Field. Chil. J. Agric. Res. 2016, 76, 163–169. [Google Scholar] [CrossRef]
- Lan, Y.; Li, W.; Wei, S.; Huang, H.; Liu, Z.; Huang, Z. Multiple Resistance to ACCase- and ALS-Inhibiting Herbicides in Black-Grass (Alopecurus myosuroides Huds.) in China. Pestic. Biochem. Physiol. 2022, 184, 105127. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Fang, F.; Xue, D.; Li, R.; Gao, X.; Li, M. A Novel Naturally Phe206Tyr Mutation Confers Tolerance to ALS-Inhibiting Herbicides in Alopecurus myosuroides. Pestic. Biochem. Physiol. 2022, 186, 105156. [Google Scholar] [CrossRef]
- Ge, L. Resistance of Black Grass to Fenoxaprop-P-ethyl and Mesosulfuron-methyl in Wheat Field; Shandong Agricultural University: Tai’an, China, 2019. (In Chinese) [Google Scholar]
- Liu, W.; Wu, C.; Guo, W.; Du, L.; Yuan, G.; Wang, J. Resistance Mechanisms to an Acetolactate Synthase (ALS) Inhibitor in Water Starwort (Myosoton aquaticum) Populations from China. Weed Sci. 2015, 63, 770–780. [Google Scholar] [CrossRef]
- Doyle, J.J. Isolation of Plant DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Yu, Q.; Shane Friesen, L.J.; Zhang, X.Q.; Powles, S.B. Tolerance to Acetolactate Synthase and Acetyl-Coenzyme A Carboxylase Inhibiting Herbicides in Vulpia Bromoides Is Conferred by Two Co-Existing Resistance Mechanisms. Pestic. Biochem. Physiol. 2004, 78, 21–30. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Moss, S.R.; Perryman, S.A.M.; Tatnell, L.V. Managing Herbicide-Resistant Blackgrass (Alopecurus myosuroides): Theory and Practice. Weed Technol. 2007, 21, 300–309. [Google Scholar] [CrossRef]
- Tranel, P.J.; Wright, T.R. Resistance of Weeds to ALS-Inhibiting Herbicides: What Have We Learned? Weed Sci. 2002, 50, 700–712. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S.B. Resistance to AHAS Inhibitor Herbicides: Current Understanding. Pest Manag. Sci. 2014, 70, 1340–1350. [Google Scholar] [CrossRef]
- Tranel, P.J.; Wright, T.R.; Heap, I.M. Mutations in Herbicide-Resistant Weeds to Inhibition of Acetolactate Synthase. 2022. Available online: https://www.weedscience.org/ (accessed on 23 July 2022).
- Beckie, H.J.; Tardif, F.J. Herbicide Cross Resistance in Weeds. Crop Prot. 2012, 35, 15–28. [Google Scholar] [CrossRef]
- Ashigh, J.; Tardif, F.J. ALS-Inhibitor Resistance in Populations of Eastern Black Nightshade (Solanum ptycanthum) from Ontario. Weed Technol. 2006, 20, 308–314. [Google Scholar] [CrossRef]
- Chen, J.; Huang, Z.; Zhang, C.; Huang, H.; Wei, S.; Chen, J.; Wang, X. Molecular Basis of Resistance to Imazethapyr in Redroot Pigweed (Amaranthus retroflexus L.) Populations from China. Pestic. Biochem. Physiol. 2015, 124, 43–47. [Google Scholar] [CrossRef]
- Wang, H.; Wang, H.; Zhao, N.; Zhu, B.; Sun, P.; Liu, W.; Wang, J. Multiple Resistance to PPO and ALS Inhibitors in Redroot Pigweed (Amaranthus retroflexus). Weed Sci. 2020, 68, 19–26. [Google Scholar] [CrossRef]
- Zhao, N.; Yan, Y.; Du, L.; Zhang, X.; Liu, W.; Wang, J. Unravelling the Effect of Two Herbicide Resistance Mutations on Acetolactate Synthase Kinetics and Growth Traits. J. Exp. Bot. 2020, 71, 3535–3542. [Google Scholar] [CrossRef]
- Kaundun, S.S. Resistance to Acetyl-CoA Carboxylase-Inhibiting Herbicides. Pest Manag. Sci. 2014, 70, 1405–1417. [Google Scholar] [CrossRef]
- Délye, C.; Menchari, Y.; Guillemin, J.-P.; Matéjicek, A.; Michel, S.; Camilleri, C.; Chauvel, B. Status of Black Grass (Alopecurus myosuroides) Resistance to Acetyl-Coenzyme A Carboxylase Inhibitors in France. Weed Res. 2007, 47, 95–105. [Google Scholar] [CrossRef]
- Yu, Q.; Han, H.; Vila-Aiub, M.M.; Powles, S.B. AHAS Herbicide Resistance Endowing Mutations: Effect on AHAS Functionality and Plant Growth. J. Exp. Bot. 2010, 61, 3925–3934. [Google Scholar] [CrossRef]
- Yu, Q.; Han, H.; Li, M.; Purba, E.; Walsh, M.J.; Powles, S.B. Resistance Evaluation for Herbicide Resistance–Endowing Acetolactate Synthase (ALS) Gene Mutations Using Raphanus Raphanistrum Populations Homozygous for Specific ALS Mutations. Weed Res. 2012, 52, 178–186. [Google Scholar] [CrossRef]
- Duggleby, R.G.; Pang, S.S.; Yu, H.; Guddat, L.W. Systematic Characterization of Mutations in Yeast Acetohydroxyacid Synthase. Eur. J. Biochem. 2003, 270, 2895–2904. [Google Scholar] [CrossRef]
- Le, D.T.; Yoon, M.-Y.; Tae Kim, Y.; Choi, J.-D. Two Consecutive Aspartic Acid Residues Conferring Herbicide Resistance in Tobacco Acetohydroxy Acid Synthase. Biochim. Biophys. Acta—Proteins Proteomics 2005, 1749, 103–112. [Google Scholar] [CrossRef]
- Zhao, N.; Yan, Y.; Ge, L.; Zhu, B.; Liu, W.; Wang, J. Target Site Mutations and Cytochrome P450s Confer Resistance to Fenoxaprop-P-Ethyl and Mesosulfuron-methyl in Alopecurus aequalis. Pest Manag. Sci. 2019, 75, 204–214. [Google Scholar] [CrossRef]
- Kaundun, S.S. An Aspartate to Glycine Change in the Carboxyl Transferase Domain of Acetyl CoA Carboxylase and Non-Target-Site Mechanism(s) Confer Resistance to ACCase Inhibitor Herbicides in a Lolium multiflorum Populatio. Pest Manag. Sci. 2010, 66, 1249–1256. [Google Scholar] [CrossRef]
- Délye, C.; Matéjicek, A.; Michel, S. Cross-Resistance Patterns to ACCase-Inhibiting Herbicides Conferred by Mutant ACCase Isoforms in Alopecurus myosuroides Huds. (Black-Grass), Re-Examined at the Recommended Herbicide Field Rate. Pest Manag. Sci. 2008, 64, 1179–1186. [Google Scholar] [CrossRef]
- Yu, Q.; Collavo, A.; Zheng, M.-Q.; Owen, M.; Sattin, M.; Powles, S.B. Diversity of Acetyl-Coenzyme A Carboxylase Mutations in Resistant Lolium Populations: Evaluation Using Clethodim. Plant Physiol. 2007, 145, 547–558. [Google Scholar] [CrossRef]
- Zhang, H.; Tweel, B.; Tong, L. Molecular Basis for the Inhibition of the Carboxyltransferase Domain of Acetyl-Coenzyme-A Carboxylase by Haloxyfop and Diclofop. Proc. Natl. Acad. Sci. USA 2004, 101, 5910–5915. [Google Scholar] [CrossRef] [Green Version]
- Délye, C.; Zhang, X.Q.; Michel, S. Matéjicek, A.; Powles, S.B. Molecular Bases for Sensitivity to Acetyl-Coenzyme A Carboxylase Inhibitors in Black-Grass. Plant Physiol. 2005, 137, 794–806. [Google Scholar] [CrossRef]
- Ahodo, K.; Oglethorpe, D.; Hicks, H.L.; Freckleton, R.P. Estimating the Farm-Level Economic Costs of Spring Cropping to Manage Alopecurus myosuroides (Black-Grass) in UK Agriculture. J. Agric. Sci. 2019, 157, 318–332. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | State | Location | Longitude/Latitude | Site | Time | Numbers of Herbicide Application | ||
|---|---|---|---|---|---|---|---|---|
| Village | Country | Province | ||||||
| HN-14 | R a | Luodian | Zhumadian | Henan | N33.056796 E114.121244 | Wheat field | 2017.05 | Fenoxaprop-P-ethyl ≥ 15 |
| Mesosulfuron-methyl ≥ 7 | ||||||||
| HN-06 | S b | Hexiao | Zhumadian | Henan | N32.707560 E114.299764 | Uncultivated land | 2017.05 | Never applied |
| Group | Herbicide | Rate (g a.i. ha−1) | Resistance Classification a |
|---|---|---|---|
| APP | Fenoxaprop-P-ethyl | 62.1 | RRR |
| Clodinafop-propargyl | 45 | RRR | |
| Diclofop-methyl | 972 | RR | |
| Fluazifop-P-butyl | 135 | RRR | |
| Haloxyfop-methyl | 40.5 | RRR | |
| Quizalofop-P-ethyl | 52.5 | S | |
| CHD | Sethoxydim | 150 | RR |
| Tralkoxydim | 390 | S | |
| DEN | Pinoxaden | 45 | R? |
| ALS | Mesosulfuron-methyl | 9 | RRR |
| Imazethapyr | 100 | S | |
| PSⅡ | Isoproturon | 900 | S |
| Herbicide | Population | C a | D b | B c | GR50 d | RI e |
|---|---|---|---|---|---|---|
| Mesosulfuron-methyl | HN-06 | 22.4664 (3.88) | 94.7370 (3.88) | −4.3322 (2.62) | 1.38 (0.3) | |
| HN-14 | 17.6951 (4.57) | 93.1030 (4.57) | −4.2756 (1.63) | 45.59 (9.92) | 33.0 | |
| Malathion + Mesosulfuron-methyl | HN-06 | 13.6476 (2.28) | 90.1691 (2.34) | −3.2643 (0.92) | 1.24 (0.10) | |
| HN-14 | 19.1257 (2.24) | 91.5331 (2.23) | −3.7303 (0.67) | 50.25 (5.01) | 40.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, X.; Yang, C.; Hu, M.; Duan, Y.; Zhang, N.; Wang, J.; Wang, H.; Liu, W. Molecular Basis of Resistance to Mesosulfuron-Methyl in a Black-Grass (Alopecurus myosuroides Huds.) Population from China. Agronomy 2022, 12, 2203. https://doi.org/10.3390/agronomy12092203
Qin X, Yang C, Hu M, Duan Y, Zhang N, Wang J, Wang H, Liu W. Molecular Basis of Resistance to Mesosulfuron-Methyl in a Black-Grass (Alopecurus myosuroides Huds.) Population from China. Agronomy. 2022; 12(9):2203. https://doi.org/10.3390/agronomy12092203
Chicago/Turabian StyleQin, Xiangyang, Cheng Yang, Mengmeng Hu, Yunxia Duan, Na Zhang, Jinxin Wang, Hengzhi Wang, and Weitang Liu. 2022. "Molecular Basis of Resistance to Mesosulfuron-Methyl in a Black-Grass (Alopecurus myosuroides Huds.) Population from China" Agronomy 12, no. 9: 2203. https://doi.org/10.3390/agronomy12092203