
Drip Irrigation Depth Alters Root Morphology and Architecture and Cold Resistance of Alfalfa
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location and Plant Materials
2.2. Experimental Design and Treatments
2.3. Root Morphology and Architecture
2.4. Semi-Lethal Temperature
2.5. Root Physiological Indicators
2.6. Statistical Analyses
3. Results
3.1. Biomass
3.2. Root Morphological Indicators
3.3. Root System Architecture
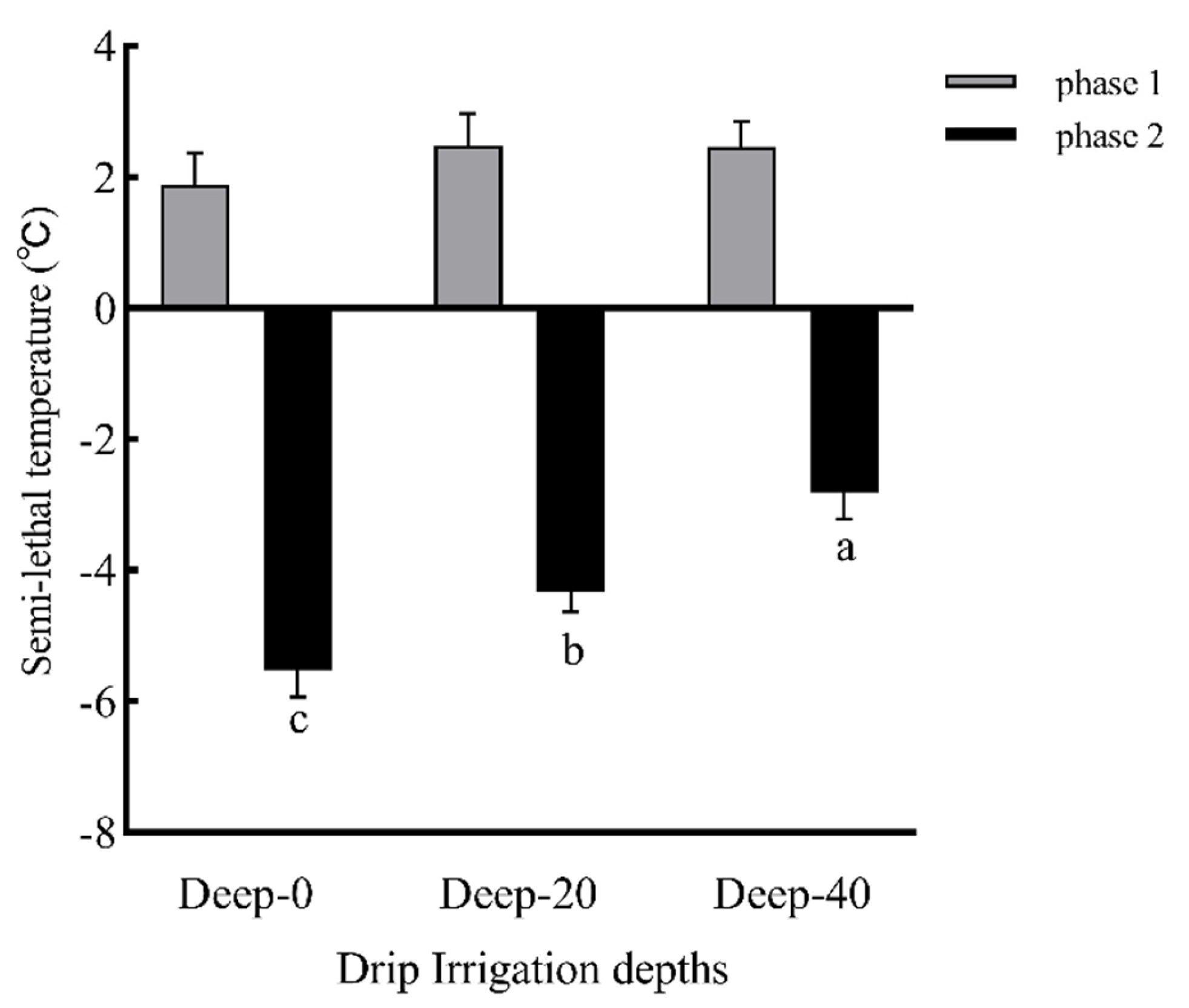
3.4. Semi-Lethal Temperature
3.5. Correlation Analysis between LT50 and Phenotypic Traits
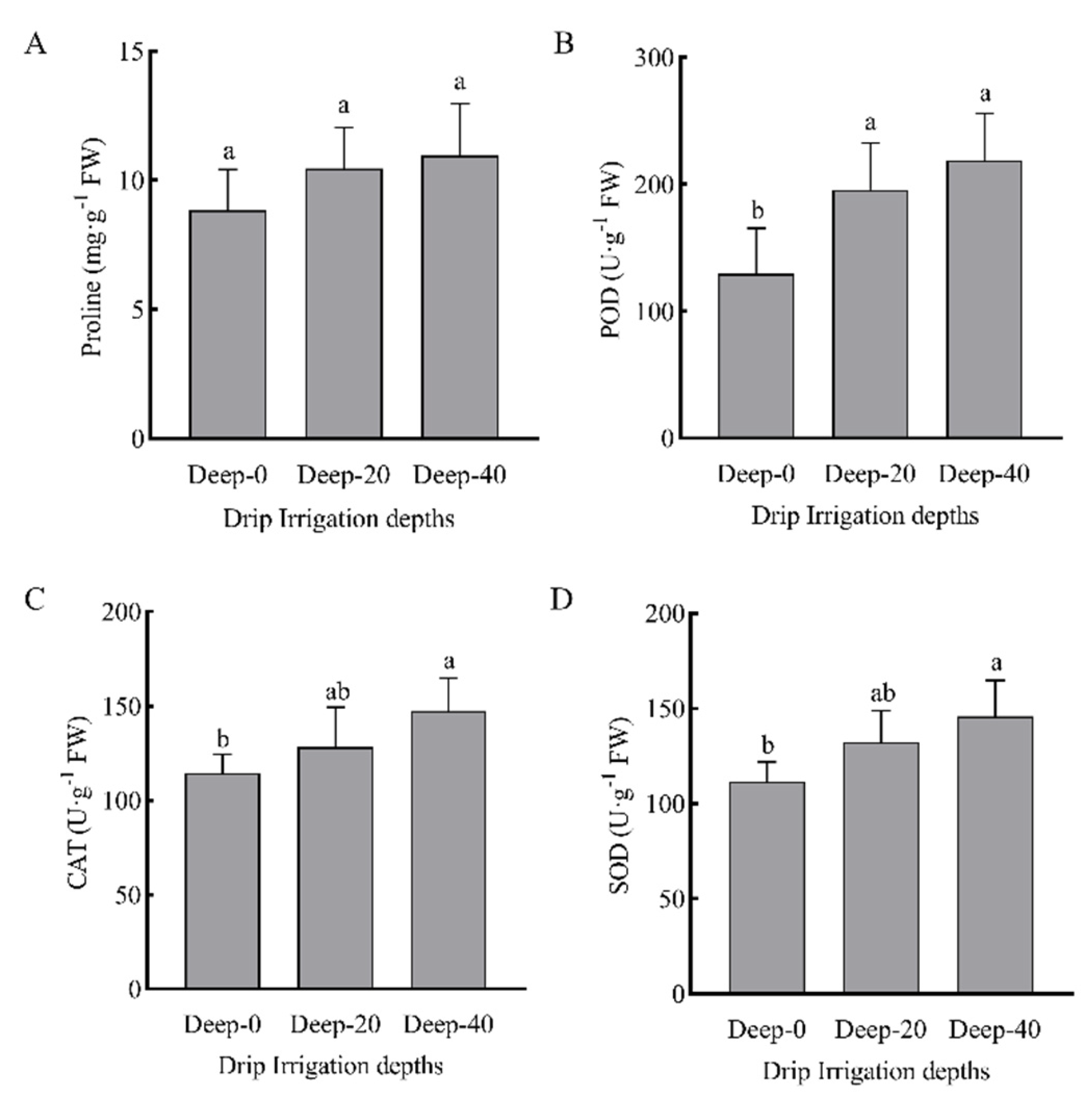
3.6. Reactive Oxygen Species and MDA
3.7. Antioxidant System Activity
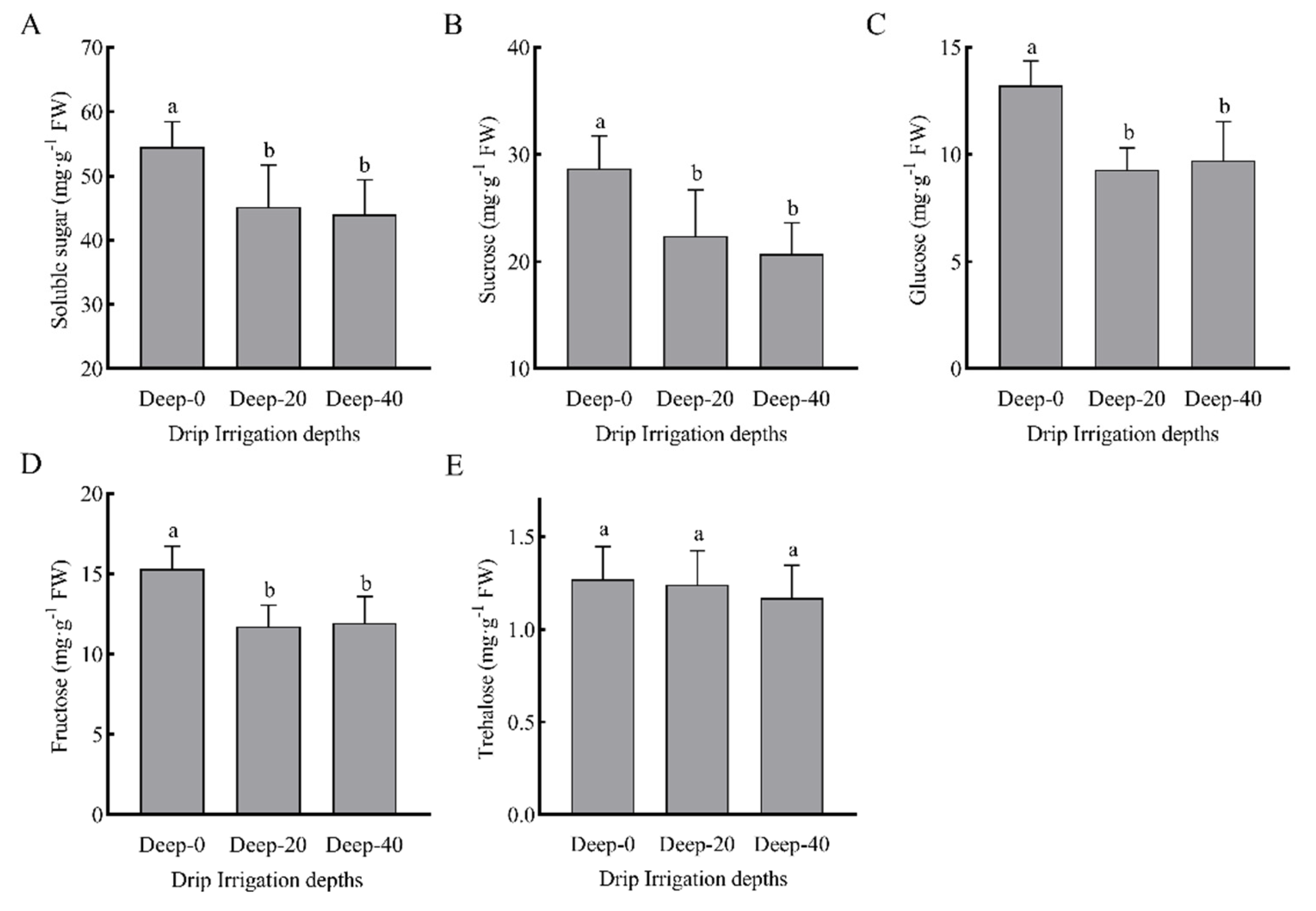
3.8. Non-Structural Carbohydrates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Russelle, M.P. Alfalfa. Am. Sci. 2001, 89, 252–261. [Google Scholar] [CrossRef]
- Carol, K.A. Reconstructing patterns of temperature, phenology, and frost damage over 124 years. Ecology 2013, 94, 41–50. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Li, X.L.; Li, F.; Tao, Y.; Liu, L.; Wang, Z.L.; Sun, Q.Z. Response of alfalfa root traits to fall dormancy and its effect on winter hardiness. Sci. Agric. Sin. 2015, 48, 1689–1701. [Google Scholar] [CrossRef]
- Schwinning, S.; Starr, B.I.; Ehleringer, J.R. Dominant cold desert plants do not partition warm season precipitation by event size. Oecologia 2003, 136, 252–260. [Google Scholar] [CrossRef]
- Schwinning, S.; Sala, O.E. Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia 2004, 141, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Be’langer, G.; Rochette, P.; Castonguay, Y.; Bootsma, A.; Mongrain, D.; Ryan, D.A.J. Climate change and winter survival of perennial forage crops in Eastern Canada. Agron. J. 2002, 94, 1120–1130. [Google Scholar] [CrossRef]
- Zhang, L.J.; Zhong, T.X.; Xu, L.X.; Han, L.B.; Zhang, X.Z. Water Deficit Irrigation Impacts on Antioxidant Metabolism Associated with Freezing Tolerance in Creeping Bentgrass. J. Am. Soc. Hortic. Sci. 2015, 140, 323–332. [Google Scholar] [CrossRef]
- Xu, H.Y.; Tong, Z.Y.; He, F.; Li, X.L. Response of Alfalfa (Medicago sativa L.) to Abrupt Chilling as Reflected by Changes in Freezing Tolerance and Soluble Sugars. Agronomy 2020, 10, 255–270. [Google Scholar] [CrossRef]
- Castonguay, Y.; Laberge, S.; Brummer, E.C.; Volenec, J.J. Alfalfa Winter Hardiness: A Research Retrospective and Integrated Perspective. Adv. Agronomy 2006, 90, 203–265. [Google Scholar]
- Li, S.; Wan, L.Q.; Nie, Z.N.; Li, X.L. Fractal and Topological Analyses and Antioxidant Defense Systems of Alfalfa (Medicago sativa L.) Root System under Drought and Rehydration Regimes. Agronomy 2020, 10, 805–826. [Google Scholar] [CrossRef]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, K.L.; Smith, D. Association of various morphological character and seed germination with the winter hardiness of alfalfa. Crop Sci. 1963, 3, 234–237. [Google Scholar] [CrossRef]
- Schwab, P.M.; Barnes, D.K.; Sheaffer, C.C.; Li, P.H. Factors Affecting a Laboratory Evaluation of Alfalfa Cold Tolerance. Crop Sci. 1996, 36, 318–324. [Google Scholar] [CrossRef]
- Smith, D. Root Branching of Alfalfa Varieties and Strains. Agron. J. 1951, 43, 573–575. [Google Scholar] [CrossRef]
- Xu, H.Y.; Li, Z.Y.; Tong, Z.Y.; He, F.; Li, X.L. Metabolomic analyses reveal substances that contribute to the increased freezing tolerance of alfalfa (Medicago sativa L.) after continuous water deficit. BMC Plant Biol. 2020, 20, 15. [Google Scholar] [CrossRef]
- Kong, R.; Henry, H. Interactions of plant growth responses to spring freezing and summer drought: A multispecies comparison. Am. J. Bot. 2019, 106, 531–539. [Google Scholar] [CrossRef]
- Kong, R.; Henry, H. Cross acclimation effects of spring freezing and summer drought on plant functional groups and ecosystem properties. Environ. Exp. Bot. 2019, 164, 52–57. [Google Scholar] [CrossRef]
- Gama, G.F.V.; Oliveira, R.M.d.; Pinheiro, D.T.; Silva, L.J.d.; Dias, D.C.F.d.S. Yield and physiological quality of wheat seeds produced under different irrigation depths and leaf Silicon. Semin. Ciências Agrárias 2021, 42, 2233–2252. [Google Scholar] [CrossRef]
- Bouma, T.J.; Nielsen, K.L.; Hal, J.V.; Koutstaal, B. Root system topology and diameter distribution of species from habitats differing in inundation frequency. Funct. Ecol. 2001, 15, 360–369. [Google Scholar] [CrossRef]
- Tanabe, M.; Kayama, A.; Kato, R.; Ito, Y. Estimation of the fractal dimension of freature surface patterns by box-counting method. Fractals 1999, 7, 335–340. [Google Scholar] [CrossRef]
- Anower, M.R.; Fennell, A.; Boe, A.; Mott, I.W.; Peel, M.D.; Wu, Y. Physiological and molecular characterisation of lucerne (Medicago sativa L.) germplasm with improved seedling freezing tolerance. Crop Pasture Sci. 2016, 67, 655–665. [Google Scholar] [CrossRef]
- Ma, L.-h.; Liu, X.-l.; Wang, Y.-k.; Wu, P.-t. Effects of drip irrigation on deep root distribution, rooting depth, and soil water profile of jujube in a semiarid region. Plant Soil 2013, 373, 995–1006. [Google Scholar] [CrossRef]
- Cullen, P.W.; Turner, A.K.; Wilson, J.H. The effect of irrigation depth on root growth of some pasture species. Plant Soil 1972, 37, 345–352. [Google Scholar] [CrossRef]
- Zhang, X.; Pei, D.; Chen, S. Root growth and soil water utilization of winter wheat in the North China Plain. Hydrol. Processes 2004, 18, 2275–2287. [Google Scholar] [CrossRef]
- Kume, T.; Sekiya, N.; Yano, K. Heterogeneity in spatial P-distribution and foraging capability by Zea mays: Effects of patch size and barriers to restrict root proliferation within a patch. Ann. Bot. 2006, 98, 1271–1277. [Google Scholar] [CrossRef]
- Tian, H.Y.; De Smet, I.; Ding, Z.J. Shaping a root system: Regulating lateral versus primary root growth. Trends Plant Sci. 2014, 19, 426–431. [Google Scholar] [CrossRef]
- Padilla, F.M.; Miranda, J.D.; Jorquera, M.J.; Pugnaire, F.I. Variability in amount and frequency of water supply affects roots but not growth of arid shrubs. Plant Ecol. 2009, 204, 261–270. [Google Scholar] [CrossRef]
- Withington, J.M.; Reich, P.B.; Oleksyn, J.; Eissenstat, D.M. Comparisons of structure and life span in roots and leaves among temperate trees. Ecol. Monogr. 2006, 76, 381–397. [Google Scholar] [CrossRef]
- Rewald, B.; Ephrath, J.E.; Rachmilevitch, S. A root is a root is a root? Water uptake rates of Citrus root orders. Plant Cell Environ. 2011, 34, 33–42. [Google Scholar] [CrossRef]
- Guo, D.L.; Mitchell, R.J.; Hendricks, J.J. Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest. Oecologia 2004, 140, 450–457. [Google Scholar] [CrossRef]
- Hassouni, K.; Alahmad, S.; Belkadi, B.; Filali-Maltouf, A.; Hickey, L.T.; Bassi, F.M. Root System Architecture and Its Association with Yield under Different Water Regimes in Durum Wheat. Crop Sci. 2018, 58, 2331–2346. [Google Scholar] [CrossRef] [Green Version]
- Bauhus, J.; Khanna, P.K.; Menden, N. Aboveground and belowground interactions in mixed plantations of Eucalyptus globulus and Acacia mearnsii. Can. J. For. Res. 2000, 30, 1886–1894. [Google Scholar] [CrossRef]
- Enquist, B.J.; Niklas, K.J. Global allocation rules for patterns of biomass partitioning in seed plants. Science 2002, 295, 1517–1520. [Google Scholar] [CrossRef] [PubMed]
- Guswa, A.J. Effect of plant uptake strategy on the water−optimal root depth. Water Resour. Res. 2010, 46. [Google Scholar] [CrossRef]
- Li, Z.; Wan, L.; Li, S.; Li, X.; He, F.; Tong, Z. Plastic response of Medicago sativa L. root system traits and cold resistance to simulated rainfall events. PeerJ 2021, 9, e11962. [Google Scholar] [CrossRef]
- Lynch, J.P. Root phenotypes for improved nutrient capture: An underexploited opportunity for global agriculture. New Phytol. 2019, 223, 548–564. [Google Scholar] [CrossRef]
- Yan, X.; Liao, H.; Nian, H. Principles and Applications of Root Biology; Science Press: Beijing, China, 2007. [Google Scholar]
- De Sme, I.; Signora, L.; Beeckman, T.; Inzé, D.; Foyer, C.H.; Zhang, H. An abscisic acid-sensitive checkpoint in lateral root development of Arabidopsis. Plant J. 2003, 33, 543–555. [Google Scholar] [CrossRef]
- Phene, C.J.; Detar, W.R.; Clark, D.A. Real-time irrigation scheduling of cotton with an automated pan evaporation system. Appl. Eng. Agric. 1992, 8, 787–793. [Google Scholar] [CrossRef]
- Green, S.R.; Clothier, B.E. Root water uptake by kiwifruit vines following partial wetting of the root zone. Plant Soil 1995, 173, 317–328. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Mahmud, J.A.; Suzuki, T.; Fujita, M. Insights into spermine-induced combined high temperature and drought tolerance in mung bean: Osmoregulation and roles of antioxidant and glyoxalase system. Protoplasma 2017, 254, 445–460. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puyang, X.; An, M.; Han, L.; Zhang, X. Protective effect of spermidine on salt stress induced oxidative damage in two Kentucky bluegrass (Poa pratensis L.) cultivars. Ecotoxicol. Environ. Saf. 2015, 117, 96–106. [Google Scholar] [CrossRef]
- Katuwal, K.B.; Schwartz, B.; Jespersen, D. Desiccation avoidance and drought tolerance strategies in bermudagrasses. Environ. Exp. Bot. 2020, 171, 103947. [Google Scholar] [CrossRef]
- Bertrand, A.; Bipfubusa, M.; Claessens, A.; Rocher, S.; Castonguay, Y. Effect of photoperiod prior to cold acclimation on freezing tolerance and carbohydrate metabolism in alfalfa (Medicago sativa L.). Plant Sci. 2017, 264, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Castonguay, Y.; Bertrand, A.; Michaud, R.; Laberge, S. Cold-Induced Biochemical and Molecular Changes in Alfalfa Populations Selectively Improved for Freezing Tolerance. Crop Sci. 2011, 51, 2132–2144. [Google Scholar] [CrossRef]
- Panjtandoust, M.; Wolyn, D.J.; Navabi, A. Asparagus cultivars with varying adaptation to southern Ontario differ for induction of freezing tolerance in the fall. Can. J. Plant Sci. 2016, 96, 252–264. [Google Scholar] [CrossRef]
- Panjtandoust, M.; Wolyn, D.J. Freezing Tolerance Attributes during Spring Deacclimation for Three Asparagus Cultivars with Varying Adaptation to Southern Ontario. J. Agron. Crop Sci. 2016, 141, 22–33. [Google Scholar] [CrossRef]
- Kalberer, S.R.; Wisniewski, M.; Arora, R. Deacclimation and reacclimation of cold-hardy plants: Current understanding and emerging concepts. Plant Sci. 2006, 171, 3–16. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Li, X.; He, F. Drip Irrigation Depth Alters Root Morphology and Architecture and Cold Resistance of Alfalfa. Agronomy 2022, 12, 2192. https://doi.org/10.3390/agronomy12092192
Li Z, Li X, He F. Drip Irrigation Depth Alters Root Morphology and Architecture and Cold Resistance of Alfalfa. Agronomy. 2022; 12(9):2192. https://doi.org/10.3390/agronomy12092192
Chicago/Turabian StyleLi, Zhensong, Xianglin Li, and Feng He. 2022. "Drip Irrigation Depth Alters Root Morphology and Architecture and Cold Resistance of Alfalfa" Agronomy 12, no. 9: 2192. https://doi.org/10.3390/agronomy12092192