
Effects of Digested Pig Slurry on Photosynthesis, Carbohydrate Metabolism and Yield of Tomato (Solanum lycopersicum L.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Fertilizer and Substrate Treatments
2.3. Sampling and Measurements
2.3.1. Growth Measurements
2.3.2. Leaf Photosynthesis Measurements
2.3.3. Leaf Carbohydrate Contents and Metabolism Enzymes Measurements
2.3.4. Fruit Carbohydrate Contents and Metabolism Enzymes Measurements
2.4. Statistical Analysis
3. Results
3.1. Tomato Growth and Yield
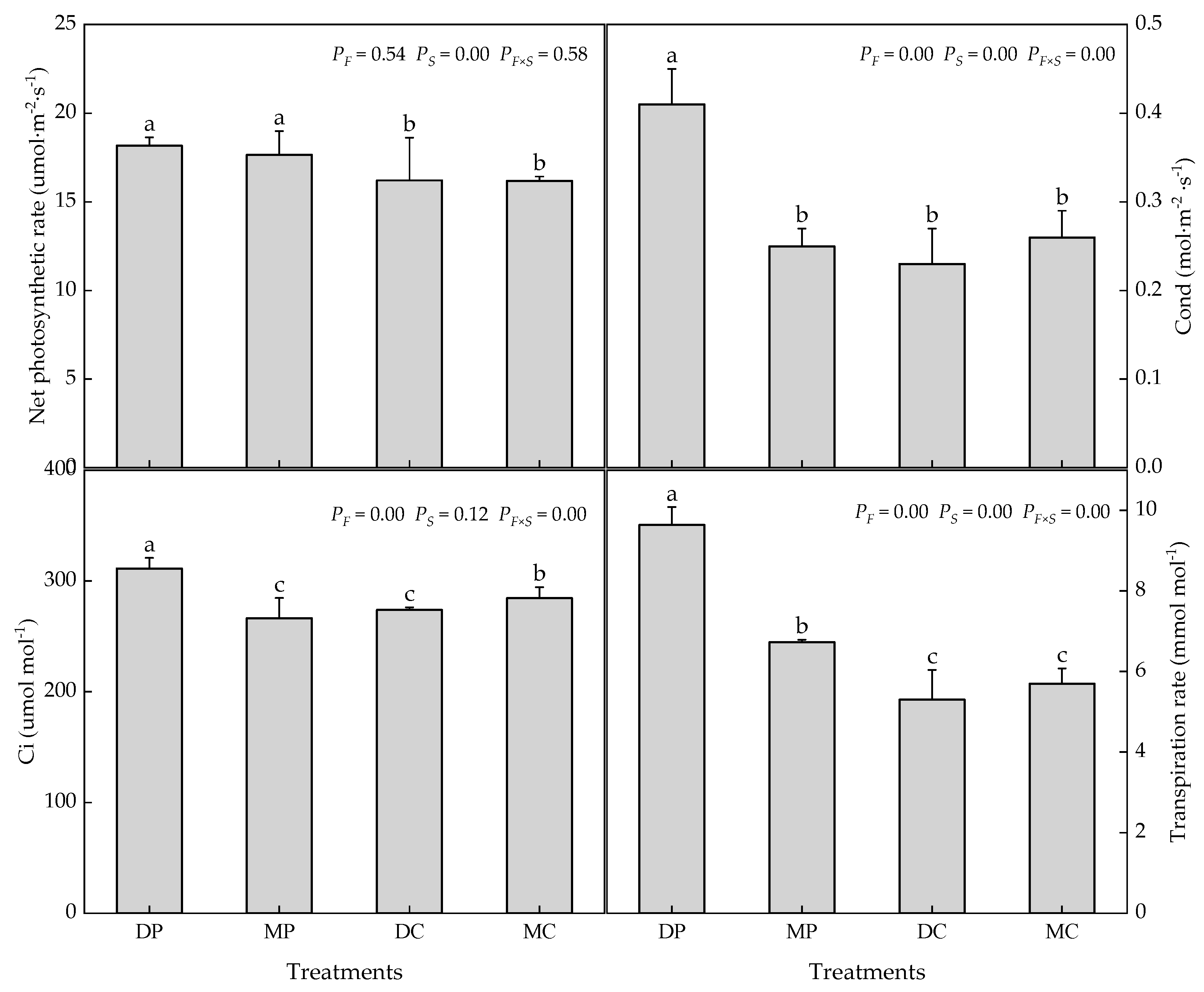
3.2. Photosynthetic Parameters and Carbohydrate Metabolism of Tomato Leaves
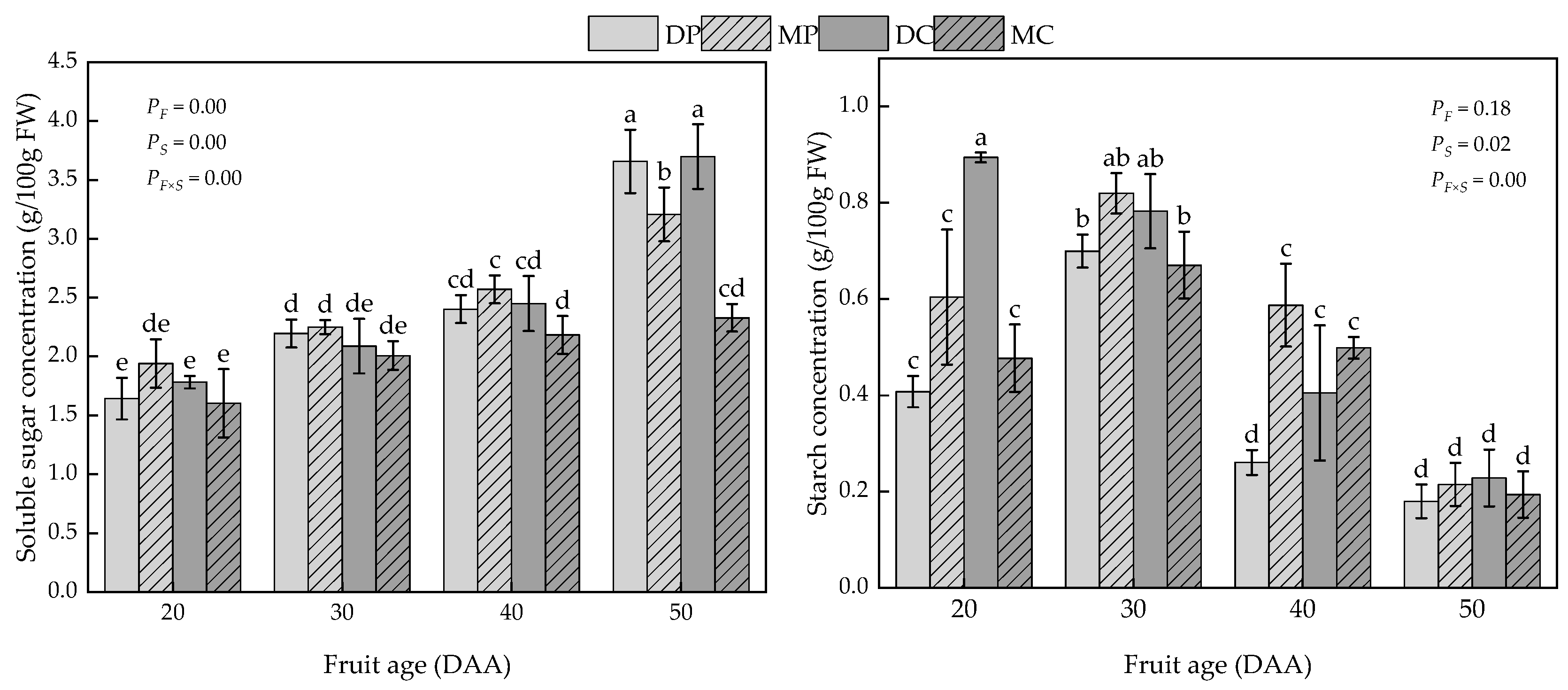
3.3. Carbon Supply Flux and Carbohydrate Contents of Tomato Fruits
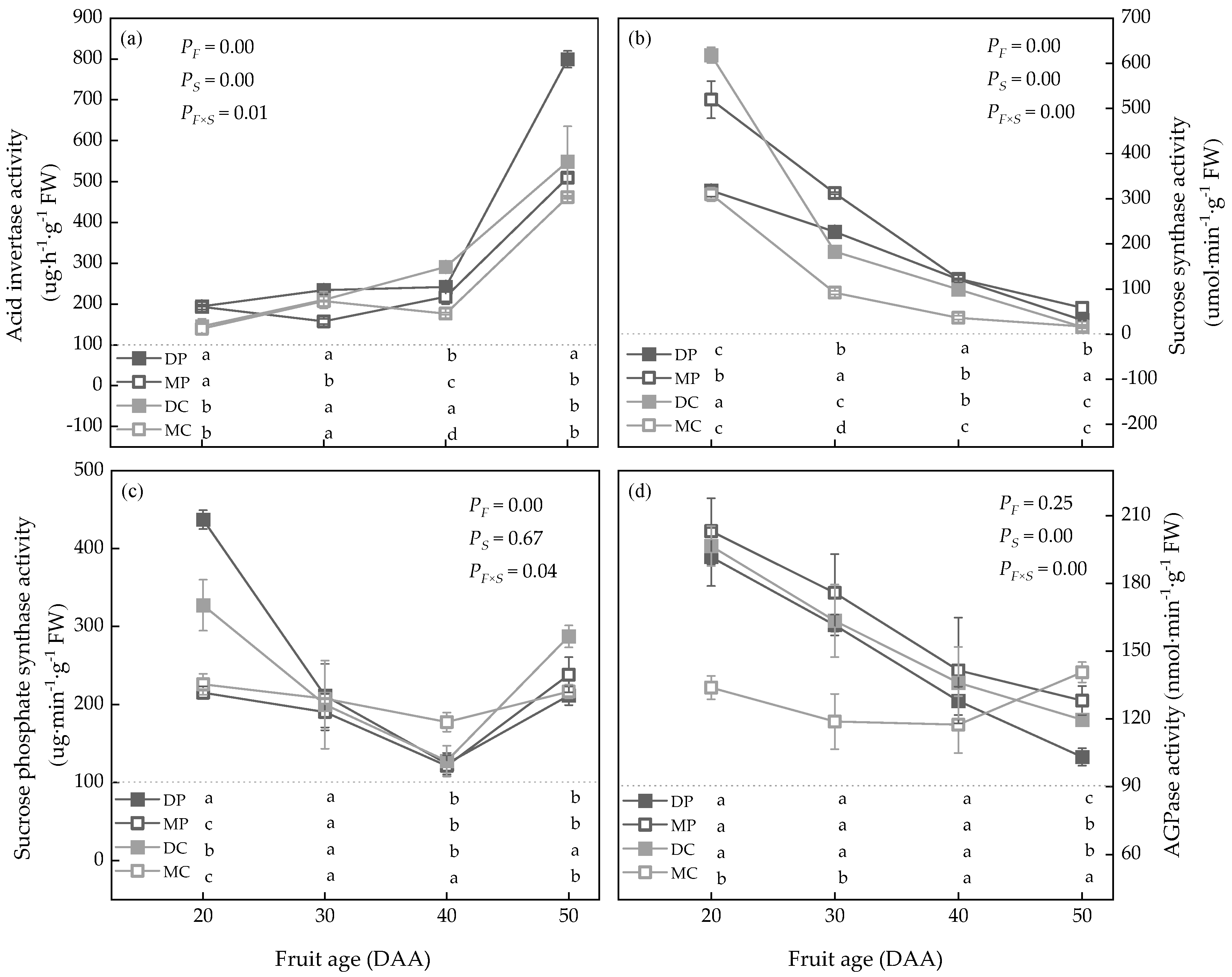
3.4. The Variation of Carbohydrate Metabolism Enzymes of Tomato Fruit and Correlation Analysis of Carbohydrate Metabolism
4. Discussion
4.1. Yield and Total Biomass Production
4.2. Photosynthesis and Carbohydrate Metabolism in Tomato Leaves
4.3. Carbohydrate Contents of Tomato Fruits
4.4. Carbohydrate Metabolism Enzymes Affect Fruit Yield and Quality
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valentinuzzi, F.; Cavani, L.; Porfido, C.; Terzano, R.; Pii, Y.; Cesco, S.; Marzadori, C.; Mimmo, T. The fertilising potential of manure-based biogas fermentation residues: Pelleted vs. liquid digestate. Heliyon 2020, 6, e03325. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Selim, S.; Jaakkola, S.; Mäkelä, P.S. Chemical composition and in vitro digestibility of whole-crop maize fertilized with synthetic fertilizer or digestate and harvested at two maturity stages in Boreal growing conditions. Agric. Food Sci. 2017, 26, 47–55. [Google Scholar] [CrossRef]
- Surendra, K.C.; Takara, D.; Hashimoto, A.G.; Khanal, S.K. Biogas as a sustainable energy source for developing countries: Opportunities and challenges. Renew. Sustain. Energy Rev. 2014, 31, 846–859. [Google Scholar] [CrossRef]
- Li, J.; Duan, N.; Guo, S.; Shao, L.; Lin, C.; Wang, J.; Hou, J.; Hou, Y.; Meng, J.; Han, M. Renewable resource for agricultural ecosystem in China: Ecological benefit for biogas by-product for planting. Ecol. Inform. 2012, 12, 101–110. [Google Scholar] [CrossRef]
- You, L.; Yu, S.; Liu, H.; Wang, C.; Zhou, Z.; Zhang, L.; Hu, D. Effects of biogas slurry fertilization on fruit economic traits and soil nutrients of Camellia oleifera Abel. PLoS ONE. 2019, 14, e0208289. [Google Scholar] [CrossRef]
- Wu, J.; Yang, Q.; Yang, G.; Shen, F.; Zhang, X.-H.; Zhang, Y.-Z. Effects of Biogas Slurry on Yield and Quality of Oil-Seed Rape. J. Plant Nutr. 2013, 36, 2084–2098. [Google Scholar] [CrossRef]
- Xu, Z.-M.; Wang, Z.; Gao, Q.; Wang, L.-L.; Chen, L.-L.; Li, Q.-G.; Jiang, J.-J.; Ye, H.-J.; Wang, D.-S.; Yang, P. Influence of irrigation with microalgae-treated biogas slurry on agronomic trait, nutritional quality, oxidation resistance, and nitrate and heavy metal residues in Chinese cabbage. J. Environ. Manag. 2019, 244, 453–461. [Google Scholar] [CrossRef]
- Cristina, G.; Camelin, E.; Tommasi, T.; Fino, D.; Pugliese, M. Anaerobic digestates from sewage sludge used as fertilizer on a poor alkaline sandy soil and on a peat substrate: Effects on tomato plants growth and on soil properties. J. Environ. Manag. 2020, 269, 110767. [Google Scholar] [CrossRef]
- Zheng, J.; Ma, J.; Feng, Z.; Zhu, C.; Wang, J.; Wang, Y. Effects of Biogas Slurry Irrigation on Tomato (Solanum Lycopersicum L.) Physiological and Ecological Indexes, Yield and Quality as Well as Soil Environment. Appl. Ecol. Environ. Res. 2020, 18, 1013–1029. [Google Scholar] [CrossRef]
- Morra, L.; Cozzolino, E.; Salluzzo, A.; Modestia, F.; Bilotto, M.; Baiano, S.; Del Piano, L. Plant Growth, Yields and Fruit Quality of Processing Tomato (Solanum lycopersicon L.) as Affected by the Combination of Biodegradable Mulching and Digestate. Agronomy 2021, 11, 100. [Google Scholar] [CrossRef]
- Kanayama, Y. Sugar Metabolism and Fruit Development in the Tomato. Hortic. J. 2017, 86, 417–425. [Google Scholar] [CrossRef]
- Granato, D.; Branco, F.G.; Nazzaro, F.; Cruz, A.G.; Faria, J.A.F. Functional Foods and Nondairy Probiotic Food Development: Trends, Concepts, and Products. Compr. Rev. Food Sci. Food Saf. 2010, 9, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.-B.; Luo, X.-P.; Song, C.-F.; Zhang, M.-X.; Shan, S.-D. Concentrated biogas slurry enhanced soil fertility and tomato quality. Acta Agric. Scand. Sect. B Soil Plant Sci. 2010, 60, 262–268. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, J.; Ren, R.; Fan, B.; Hou, L.; Li, J. Effects of different organic nutrient solution formulations and supplementation on tomato fruit quality and aromatic volatiles. Arch. Agron. Soil Sci. 2020, 67, 563–575. [Google Scholar] [CrossRef]
- Möller, K.; Stinner, W.; Deuker, A.; Leithold, G. Effects of different manuring systems with and without biogas digestion on nitrogen cycle and crop yield in mixed organic dairy farming systems. Nutr. Cycl. Agroecosyst. 2008, 82, 209–232. [Google Scholar] [CrossRef]
- Nkoa, R. Agricultural benefits and environmental risks of soil fertilization with anaerobic digestates: A review. Agron. Sustain. Dev. 2013, 34, 473–492. [Google Scholar] [CrossRef]
- Bilalis, D.; Krokida, M.; Roussis, I.; Papastylianou, P.; Travlos, I.; Cheimona, N.; Dede, A. Effects of organic and inorganic fertilization on yield and quality of processing tomato (Lycopersicon esculentum Mill.). Folia Hortic. 2018, 30, 321–332. [Google Scholar] [CrossRef]
- Barzee, T.J.; Edalati, A.; El-Mashad, H.; Wang, D.; Scow, K.; Zhang, R. Digestate Biofertilizers Support Similar or Higher Tomato Yields and Quality Than Mineral Fertilizer in a Subsurface Drip Fertigation System. Front. Sustain. Food Syst. 2019, 3, 58. [Google Scholar] [CrossRef]
- Li, M.; Feng, F.; Cheng, L. Expression patterns of genes involved in sugar metabolism and accumulation during apple fruit development. PLoS ONE 2012, 7, e33055. [Google Scholar] [CrossRef]
- Osorio, S.; Ruan, Y.L.; Fernie, A.R. An update on source-to-sink carbon partitioning in tomato. Front. Plant Sci. 2014, 5, 516. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ma, L.; Li, J. Effects of all-organic nutrient solution and water coupling on quality, yield and water use efficiency of tomato. Sci. Agric. Sin. 2018, 51, 2788–2798. [Google Scholar] [CrossRef]
- Wang, H.; Lin, L.; Tang, Y.; Li, Y.; Sun, J.; Zhang, S.; Wu, X. Effects of nutrient solution based on biogas slurry on diurnal change leaf gas exchange of strawberry in soilless cultivation. J. Agric. Univ. Hebei 2019, 42, 57–64. [Google Scholar] [CrossRef]
- Xu, C.; Tian, Y.; Sun, Y.; Dong, L. Effects of Biogas Slurry Irrigation on Growth, Photosynthesis, and Nutrient Status of Perilla frutescens Seedlings. Commun. Soil Sci. Plant Anal. 2013, 44, 3381–3390. [Google Scholar] [CrossRef]
- Patanè, C.; Saita, A. Biomass, fruit yield, water productivity and quality response of processing tomato to plant density and deficit irrigation under a semi-arid Mediterranean climate. Crop Pasture Sci. 2015, 66, 224–234. [Google Scholar] [CrossRef]
- Sachdeva, M.; Mann, A.; Batta, S.K. Sucrose metabolism and expression of key enzyme activities in low and high sucrose storing sugarcane genotypes. Sugar Tech 2003, 5, 265–271. [Google Scholar] [CrossRef]
- Gruda, N.; Caron, J.; Prasad, M.; Maher, M.J. Growing Media. In Encyclopedia of Soil Sciences; Taylor & Francis Group: Abingdon, UK, 2016. [Google Scholar]
- Xie, X.; Machikowa, T.; Wonprasaid, S. Fertigation based on a nutrient balance model for cassava production in two different textured soils. Plant Prod. Sci. 2020, 23, 407–416. [Google Scholar] [CrossRef]
- Liao, L.; Dong, T.; Qiu, X.; Rong, Y.; Zhu, J. Nitrogen nutrition is a key modulator of the sugar and organic acid content in citrus fruit. PLoS ONE 2019, 14, e0223356. [Google Scholar] [CrossRef]
- Chantigny, M.H.; Angers, D.A.; Bélanger, G.; Rochette, P.; Eriksen-Hamel, N.; Bittman, S.; Buckley, K.; Massé, D.; Gasser, M.O. Yield and Nutrient Export of Grain Corn Fertilized with Raw and Treated Liquid Swine Manure. Agron. J. 2008, 100, 1303–1309. [Google Scholar] [CrossRef]
- Pan, W.L.; Madsen, I.J.; Bolton, R.P.; Graves, L.; Sistrunk, T. Ammonia/Ammonium Toxicity Root Symptoms Induced by Inorganic and Organic Fertilizers and Placement. Agron. J. 2016, 108, 2485–2492. [Google Scholar] [CrossRef]
- Reddy, K.R.; Matcha, S.K. Quantifying nitrogen effects on castor bean (Ricinus communis L.) development, growth, and photosynthesis. Ind. Crop. Prod. 2010, 31, 185–191. [Google Scholar] [CrossRef]
- Rossi, M.; Bermudez, L.; Carrari, F. Crop yield: Challenges from a metabolic perspective. Curr. Opin. Plant Biol. 2015, 25, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Luo, A.; Zhou, C.; Chen, J. The Associated With Carbon Conversion Rate and Source-Sink Enzyme Activity in Tomato Fruit Subjected to Water Stress and Potassium Application. Front Plant Sci. 2021, 12, 681145. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, Z.A.; Walker., R.R.; Liao., S.; Barril., C.; Deloire., A.J.; Clarke., S.J.; Tyerman., S.D.; Rogiers., S.Y. Expression Patterns of Genes Encoding Sugar and Potassium Transport Proteins Are Simultaneously Upregulated or Downregulated When Carbon and Potassium Availability Is Modified in Shiraz (Vitis vinifera L.) Berries. Plant Cell Physiol. 2019, 10, 2331–2342. [Google Scholar] [CrossRef] [PubMed]
- Zahoor, R.; Zhao, W.; Dong, H.; Snider, J.L.; Abid, M.; Iqbal, B.; Zhou, Z. Potassium improves photosynthetic tolerance to and recovery from episodic drought stress in functional leaves of cotton (Gossypium hirsutum L.). Plant Physiol. Bioch. 2017, 119, 21–32. [Google Scholar] [CrossRef]
- Moya, C.; Oyanedel, E.; Verdugo, G.; Flores, M.F.; Urrestarazu, M.; Álvaro, J.E. Increased Electrical Conductivity in Nutrient Solution Management Enhances Dietary and Organoleptic Qualities in Soilless Culture Tomato. HortScience 2017, 52, 868–872. [Google Scholar] [CrossRef]
- Yin, Y.G.; Kobayashi, Y.; Sanuki, A.; Kondo, S.; Fukuda, N.; Ezura, H.; Sugaya, S.; Matsukura, C. Salinity induces carbohydrate accumulation and sugar-regulated starch biosynthetic genes in tomato (Solanum lycopersicum L. cv. ‘Micro-Tom’) fruits in an ABA- and osmotic stress-independent manner. J. Exp. Bot. 2010, 61, 563–574. [Google Scholar] [CrossRef]
- Biais, B.; Benard, C.; Beauvoit, B.; Colombie, S.; Prodhomme, D.; Menard, G.; Bernillon, S.; Gehl, B.; Gautier, H.; Ballias, P.; et al. Remarkable reproducibility of enzyme activity profiles in tomato fruits grown under contrasting environments provides a roadmap for studies of fruit metabolism. Plant Physiol. 2014, 164, 1204–1221. [Google Scholar] [CrossRef]
- Colombie, S.; Beauvoit, B.; Nazaret, C.; Benard, C.; Vercambre, G.; Le Gall, S.; Biais, B.; Cabasson, C.; Maucourt, M.; Bernillon, S.; et al. Respiration climacteric in tomato fruits elucidated by constraint-based modelling. New Phytol. 2017, 213, 1726–1739. [Google Scholar] [CrossRef]
- Braun, D.M.; Wang, L.; Ruan, Y.L. Understanding and manipulating sucrose phloem loading, unloading, metabolism, and signalling to enhance crop yield and food security. J. Exp. Bot. 2014, 65, 1713–1735. [Google Scholar] [CrossRef]
- Patrick, J.W. PHLOEM UNLOADING: Sieve Element Unloading and Post-Sieve Element Transport. Annu Rev. Plant Physiol Plant Mol. Biol 1997, 48, 191–222. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Wang, X.L.; Wang, X.F.; Xia, G.H.; Pan, Q.H.; Fan, R.C.; Wu, F.Q.; Yu, X.C.; Zhang, D.P. A shift of Phloem unloading from symplasmic to apoplasmic pathway is involved in developmental onset of ripening in grape berry. Plant Physiol. 2006, 142, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Beauvoit, B.; Genard, M.; Colombie, S.; Moing, A.; Vercambre, G.; Gomes, E.; Gibon, Y.; Dai, Z. Modelling predicts tomatoes can be bigger and sweeter if biophysical factors and transmembrane transports are fine-tuned during fruit development. New Phytol. 2021, 230, 1489–1502. [Google Scholar] [CrossRef]
- Ikeda, H.; Hiraga, M.; Shirasawa, K.; Nishiyama, M.; Kanayama, Y. Analysis of a tomato introgression line, IL8-3, with increased Brix content. Sci. Hortic. 2013, 153, 103–108. [Google Scholar] [CrossRef]
- Romero-Aranda, R.; Soria, T.; Cuartero, J. Tomato plant-water uptake and plant-water relationships under saline growth conditions. Plant Sci. 2001, 160, 265–272. [Google Scholar] [CrossRef]
- Sun, L.; Wang, J.; Song, J.; Du, X.; Liu, W.; Zhao, W.; Yang, L.; Li, C.; Qin, Y.; Yang, R. Systematic Analysis of Sugar Unloading and Accumulation Mechanism in Sucrose- And Hexose Accumulating Tomato Fruits. 2021; preprint. [Google Scholar] [CrossRef]
- Hubbard, N.L.; Pharr, D.M.; Huber, S.C. Sucrose phosphate synthase and other sucrose metabolizing enzymes in fruits of various species. Physiol. Plant. 2006, 82, 191–196. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| pH | Electrical Conductivity (ms/cm) | Total Organic Carbon (mg/L) | N-Tot (mg/L) | NH4+-N (mg/L) | P-Tot (mg/L) | K-Tot (mg/L) |
|---|---|---|---|---|---|---|
| 8.01 ± 0.01 | 6.88 ± 0.20 | 213.03 ± 3.01 | 1000 ± 40 | 813.33 ± 6.43 | 232.5 ± 0.71 | 697 ± 2.83 |
| Treatment | Base Fertilizer (67%) | Topdressing Fertilizer 1 (33%) |
|---|---|---|
| DP (Digested pig slurry +Peat substrate) | 2.06 L | 0.35 L time−1 × 10 times |
| MP (Mineral fertilizer + Peat substrate) | N: 2.06 g; P2O5: 1.91 g; K2O: 6.24 g | N: 0.35 g; P2O5: 0.12 g; K2O: 0.44 g |
| DC (Digested pig slurry + Cinder substrate) | 5.91 L | 0.35 L time−1 × 10 times |
| MC (Mineral fertilizer + Cinder substrate) | N: 5.91 g; P2O5: 2.60 g; K2O: 7.05 g | N: 0.35 g; P2O5: 0.12 g; K2O: 0.44 g |
| Treatment | Dry Biomass (g per Plant) | ||||
|---|---|---|---|---|---|
| Roots | Stems | Leaves | Fruits | Total | |
| DP | 8.1 ± 3.0 ab | 50.2 ± 13.4 a | 49.7 ± 9.5 a | 154.1 ± 9.7 a | 262.1 ± 27.4 a |
| MP | 6.4 ± 2.4 b | 40.1 ± 2.7 a | 48.7 ± 5.5 a | 137.5 ± 4.3 a | 233.2 ± 7.8 a |
| DC | 11.2 ± 3.3 a | 38.3 ± 5.0 a | 40.4 ± 5.4 a | 95.0 ± 15.2 b | 185.5 ± 25.6 b |
| MC | 5.4 ± 0.8 b | 23.0 ± 4.6 b | 28.9 ± 5.1 b | 51.1 ± 3.2 c | 108.5 ± 10.0 c |
| Treatment | Yield (kg·m−2) | Fruit Number (no. per Plant) | Fruit Fresh Weight (g per Fruit) |
|---|---|---|---|
| DP | 6.004 ± 0.38 a | 11.3 ± 1.2 ab | 177.1 ± 8.4 a |
| MP | 5.811 ± 0.18 a | 13.3 ± 0.6 a | 145.3 ± 3.4 b |
| DC | 3.391 ± 0.54 b | 10.0 ± 2.0 b | 103.6 ± 14.3 c |
| MC | 2.223 ± 0.14 c | 7.7 ± 0.6 c | 96.7 ± 3.3 c |
| Significance | |||
| Fertilizer | * | ns | * |
| Substrate | * | * | * |
| Fertilizer×Substrate | * | * | * |
| Treatment | Sucrose (mg·g−1 FW) | Fructose (mg·g−1 FW) | Glucose (mg·g−1 FW) | Starch (mg·g−1 FW) |
|---|---|---|---|---|
| DP | 1.0 ± 0.08 a | 16.4 ± 2.3 a | 0.3 ± 0.02 a | 0.8 ± 0.07 b |
| MP | 0.7 ± 0.12 b | 11.5 ± 2.0 b | 0.2 ± 0.01 b | 1.0 ± 0.08 ab |
| DC | 1.0 ± 0.12 a | 16.3 ± 1.7 a | 0.3 ± 0.02 a | 0.7 ± 0.03 b |
| MC | 0.5 ± 0.08 c | 8.6 ± 1.6 b | 0.1 ± 0.003 c | 1.1 ± 0.04 a |
| Significance | ||||
| Fertilizer | * | * | * | * |
| Substrate | * | ns | * | ns |
| Fertilizer×Substrate | ns | ns | * | * |
| Treatment | Sucrose Phosphate Synthase Activity (µg·min−1·g−1 FW) | Sucrose Synthase Activity (µmol·min−1·g−1 FW) | AGPase Activity 1 (nmol·min−1·g−1 FW) |
|---|---|---|---|
| DP | 348.8 ± 19.5 a | 241.5 ± 11.6 b | 77.8 ± 2.2 a |
| MP | 205.2 ± 18.3 c | 259.1 ± 9.4 ab | 65.2 ± 1.6 c |
| DC | 281.7 ± 10.2 b | 269.9 ± 11.6 a | 70.2 ± 0.2 b |
| MC | 262.7 ± 38.4 b | 182.1 ± 14.6 c | 63.3 ± 1.8 c |
| Significance | |||
| Fertilizer | * | * | * |
| Substrate | ns | * | * |
| Fertilizer×Substrate | * | * | ns |
| Treatment | Carbon Supply Flux (g C/day) | |||
|---|---|---|---|---|
| 0–20 days | 20–30 days | 30–40 days | 40–50 days | |
| DP | 0.04 ± 0.005 e | 0.18 ± 0.02 c | 0.26 ± 0.01 b | 0.39 ± 0.10 a |
| MP | 0.03 ± 0.004 e | 0.16 ± 0.01 cd | 0.23 ± 0.03 bc | 0.13 ± 0.06 bcd |
| DC | 0.03 ± 0.006 e | 0.16 ± 0.02 cd | 0.14 ± 0.02 bcd | 0.11 ± 0.03 d |
| MC | 0.02 ± 0.003 e | 0.09 ± 0.03 de | 0.15 ± 0.01 bcd | 0.16 ± 0.03 cd |
| Treatment | Fruit Dry Weight | Soluble Sugar Content | Starch Content | AI | SPS | Susy | AGPase |
|---|---|---|---|---|---|---|---|
| Fruit dry weight | 1 | ||||||
| Soluble sugar content | 0.876 ** | 1 | |||||
| Starch content | −0.560 * | −0.516 * | 1 | ||||
| AI | 0.874 ** | 0.882 ** | −0.596 * | 1 | |||
| SPS | −0.326 | −0.180 | 0.017 | 0.015 | 1 | ||
| Susy | −0.685 ** | −0.632 ** | 0.635 ** | −0.579 * | 0.400 | 1 | |
| AGPase | −0.666 ** | −0.613 * | 0.548 * | −0.562 * | 0.450 | 0.870 ** | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teng, Y.; Shang, B.; Tao, X. Effects of Digested Pig Slurry on Photosynthesis, Carbohydrate Metabolism and Yield of Tomato (Solanum lycopersicum L.). Agronomy 2022, 12, 2042. https://doi.org/10.3390/agronomy12092042
Teng Y, Shang B, Tao X. Effects of Digested Pig Slurry on Photosynthesis, Carbohydrate Metabolism and Yield of Tomato (Solanum lycopersicum L.). Agronomy. 2022; 12(9):2042. https://doi.org/10.3390/agronomy12092042
Chicago/Turabian StyleTeng, Yunfei, Bin Shang, and Xiuping Tao. 2022. "Effects of Digested Pig Slurry on Photosynthesis, Carbohydrate Metabolism and Yield of Tomato (Solanum lycopersicum L.)" Agronomy 12, no. 9: 2042. https://doi.org/10.3390/agronomy12092042