
Genome-Wide Identification of GASA Gene Family in Ten Cucurbitaceae Species and Expression Analysis in Cucumber

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Chromosomal Mapping of GASA Family Genes in 10 Cucurbitaceae Species
2.2. Gene Structure Analysis and Phylogenetic Tree Construction of GASA Family Genes in 10 Cucurbitaceae Species
2.3. Gene Duplication and Synteny Analysis of GASA Family Genes
2.4. RNA-seq Reanalysis with Cucumber Transcriptome Sequencing Data
2.5. Tissue-Specific Expression Analysis of Cucumber GASA Family Genes
2.6. The Expression Analysis of Cucumber GASA Family Genes under Stresses
3. Results
3.1. Genome-Wide Identification of GASA Genes in 10 Cucurbitaceae Species
3.2. Phylogenetic Analysis of GASA Family Genes in 10 Cucurbitaceae Species
3.3. The Gene Structure and Conserved Motifs of GASA Family Genes in 10 Cucurbitaceae Species
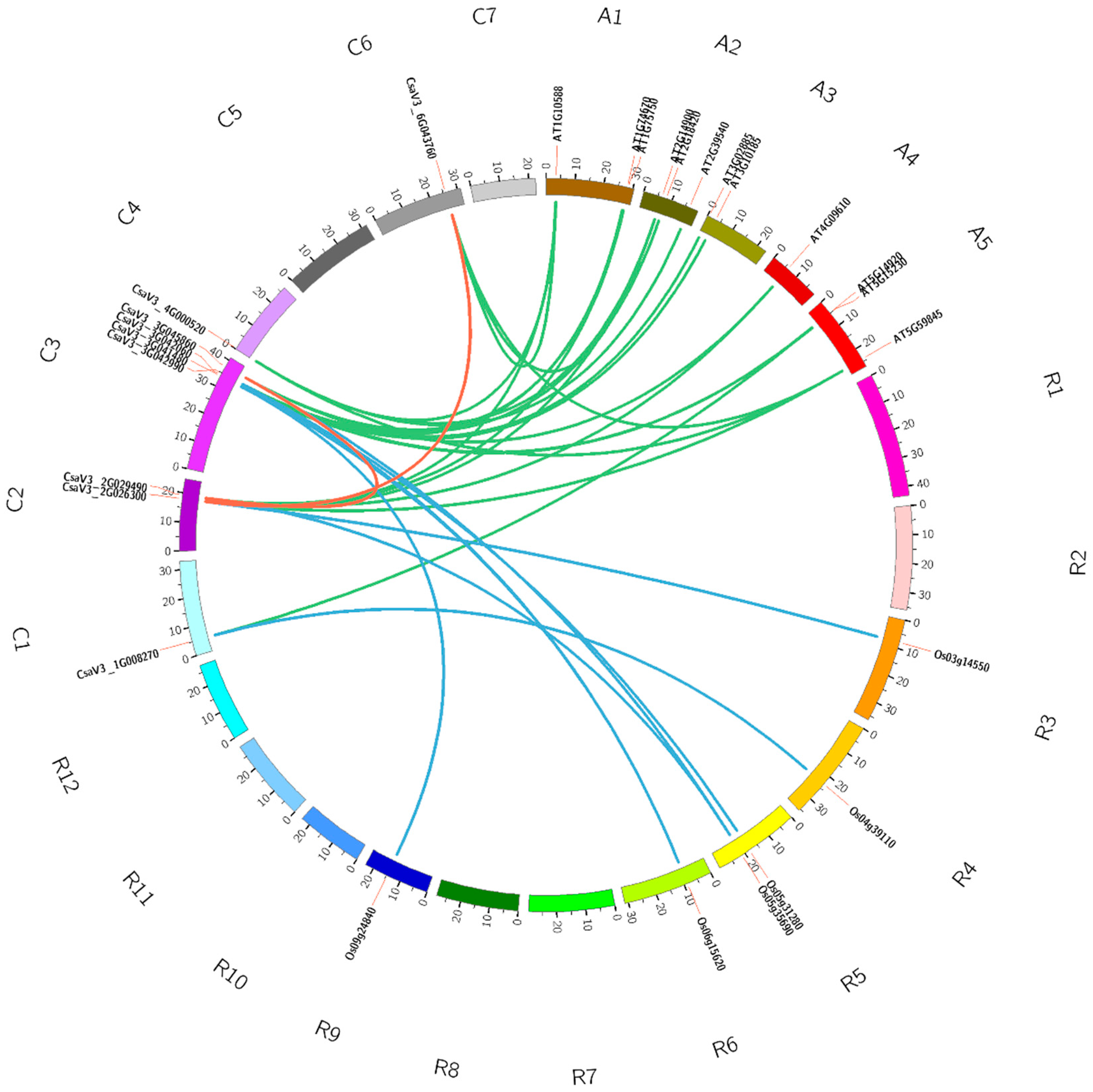
3.4. Synteny Analysis of GASA Genes among Cucumber, Arabidopsis and Rice
3.5. Analysis of Cis-Acting Elements in Cucumber GASA Genes
3.6. Tissue-Specific Expression Analysis of Cucumber GASA Genes
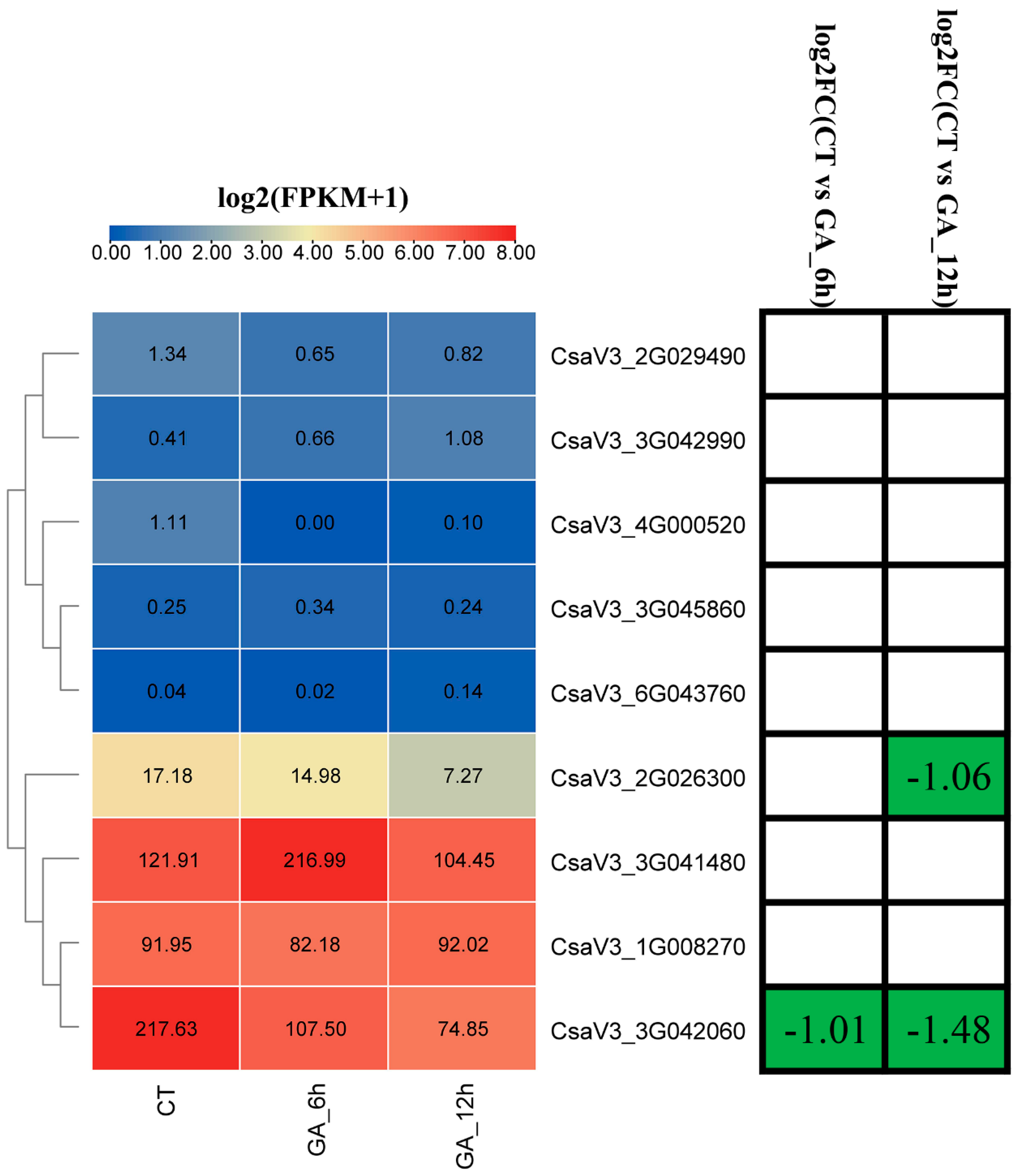
3.7. Expression Profiles of Cucumber GASA Genes under GA Treatment
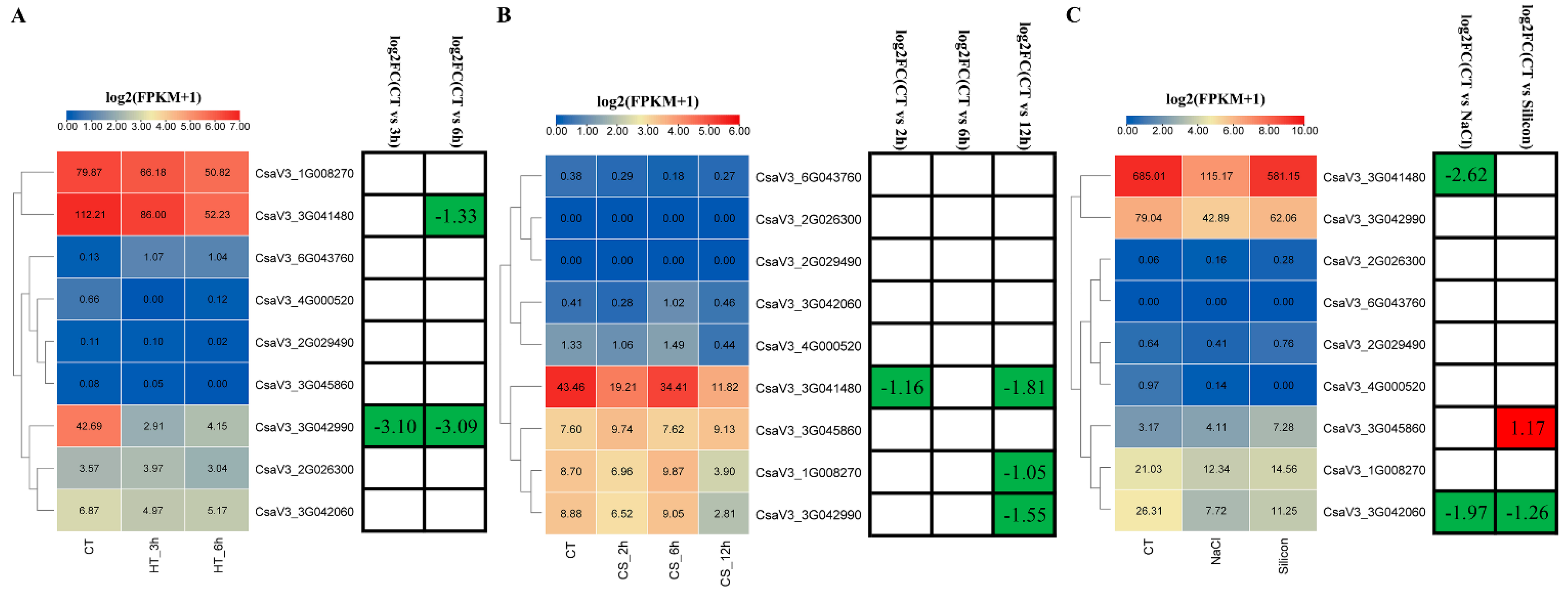
3.8. Expression Profiles of Cucumber GASA Genes under Abiotic Stresses
3.9. Expression Profiles of Cucumber GASA Genes under Biotic Stresses
3.10. Regulation Patterns of Cucumber GASA Genes under Stresses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Silverstein, K.A.; Moskal, W.A., Jr.; Wu, H.C.; Underwood, B.A.; Graham, M.A.; Town, C.D.; VandenBosch, K.A. Small cysteine-rich peptides resembling antimicrobial peptides have been under-predicted in plants. Plant J. 2007, 51, 262–280. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, A.; Kumar, P.; Sarkar, A.K. Giberellic Acid-Stimulated Transcript Proteins Evolved through Successive Conjugation of Novel Motifs and Their Subfunctionalization. Plant Physiol. 2019, 180, 998–1012. [Google Scholar] [CrossRef]
- Aubert, D.; Chevillard, M.; Dorne, A.-M.; Arlaud, G.; Herzog, M. Expression patterns of GASA genes in Arabidopsis thaliana: The GASA4 gene is up-regulated by gibberellins in meristematic regions. Plant Mol. Biol. 1998, 36, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Z.; Xu, Y.; Joo, S.-H.; Kim, S.-K.; Xue, Z.; Xu, Z.; Wang, Z.; Chong, K. OsGSR1 is involved in crosstalk between gibberellins and brassinosteroids in rice. Plant J. 2009, 57, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, X. Expression pattern of GASA, downstream genes of DELLA, in Arabidopsis. Chin. Sci. Bull. 2008, 53, 3839–3846. [Google Scholar] [CrossRef] [Green Version]
- Ben-Nissan, G.; Lee, J.-Y.; Borohov, A.; Weiss, D. GIP, a Petunia hybrida GA-induced cysteine-rich protein: A possible role in shoot elongation and transition to flowering. Plant J. 2004, 37, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Gast, R.T.; Gopalraj, M.; Olszewski, N.E. Characterization of a shoot-specific, GA3- and ABA-regulated gene from tomato. Plant J. 1992, 2, 153–159. [Google Scholar]
- Herzog, M.; Dorne, A.-M.; Grellet, F. GASA, a gibberellin-regulated gene family from Arabidopsis thaliana related to the tomato GAST1 gene. Plant Mol. Biol. 1995, 27, 743–752. [Google Scholar] [CrossRef]
- Roxrud, I.; Lid, S.E.; Fletcher, J.C.; Schmidt, E.D.L.; Opsahl-Sorteberg, H.-G. GASA4, One of the 14-Member Arabidopsis GASA Family of Small Polypeptides, Regulates Flowering and Seed Development. Plant Cell Physiol. 2007, 48, 471–483. [Google Scholar] [CrossRef]
- Fan, S.; Zhang, D.; Zhang, L.; Gao, C.; Xin, M.; Tahir, M.M.; Li, Y.; Ma, J.; Han, M. Comprehensive analysis of GASA family members in the Malus domestica genome: Identification, characterization, and their expressions in response to apple flower induction. BMC Genom. 2017, 18, 827. [Google Scholar] [CrossRef]
- Zimmermann, R.; Sakai, H.; Hochholdinger, F. The Gibberellic Acid Stimulated-Like Gene Family in Maize and Its Role in Lateral Root Development. Plant Physiol. 2010, 152, 356–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahirñak, V.; Rivarola, M.; De Urreta, M.G.; Paniego, N.; Hopp, H.E.; Almasia, N.I.; Vazquez-Rovere, C. Genome-wide Analysis of the Snakin/GASA Gene Family in Solanum tuberosum cv. Kennebec. Am. Potato J. 2016, 93, 172–188. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, S.; Xu, D.; Liu, X.; Li, X.; Xiao, W.; Cao, J.; Jiang, H.; Min, X.; Wang, J.; et al. Identification and Analysis of the GASR Gene Family in Common Wheat (Triticum aestivum L.) and Characterization of TaGASR34, a Gene Associated With Seed Dormancy and Germination. Front. Genet. 2019, 10, 980. [Google Scholar] [CrossRef] [PubMed]
- Rezaee, S.; Ahmadizadeh, M.; Heidari, P. Genome-wide characterization, expression profiling, and post-transcriptional study of GASA gene family. Gene Rep. 2020, 20, 100795. [Google Scholar] [CrossRef]
- Ahmad, B.; Yao, J.; Zhang, S.; Li, X.; Zhang, X.; Yadav, V.; Wang, X. Genome-Wide Characterization and Expression Profiling of GASA Genes during Different Stages of Seed Development in Grapevine (Vitis vinifera L.) Predict Their Involvement in Seed Development. Int. J. Mol. Sci. 2020, 21, 1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Jiao, Z.; Niu, M.-X.; Yu, X.; Huang, M.; Liu, C.; Wang, H.-L.; Zhou, Y.; Mao, W.; Wang, X.; et al. Genome-Wide Comprehensive Analysis of the GASA Gene Family in Populus. Int. J. Mol. Sci. 2021, 22, 12336. [Google Scholar] [CrossRef] [PubMed]
- Rubinovich, L.; Ruthstein, S.; Weiss, D. The Arabidopsis Cysteine-Rich GASA5 Is a Redox-Active Metalloprotein that Suppresses Gibberellin Responses. Mol. Plant 2014, 7, 244–247. [Google Scholar] [CrossRef] [Green Version]
- Rubinovich, L.; Weiss, D. The Arabidopsis cysteine-rich protein GASA4 promotes GA responses and exhibits redox activity in bacteria and in planta. Plant J. 2010, 64, 1018–1027. [Google Scholar] [CrossRef]
- Dong, L.; Wang, F.; Liu, T.; Dong, Z.; Li, A.; Jing, R.; Mao, L.; Li, Y.; Liu, X.; Zhang, K.; et al. Natural variation of TaGASR7-A1 affects grain length in common wheat under multiple cultivation conditions. Mol. Breed. 2014, 34, 937–947. [Google Scholar] [CrossRef]
- Wigoda, N.; Ben-Nissan, G.; Granot, D.; Schwartz, A.; Weiss, D. The gibberellin-induced, cysteine-rich protein GIP2 from Petunia hybrida exhibits in planta antioxidant activity. Plant J. 2006, 48, 796–805. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, C.; Peng, J.; Sun, S.; Wang, X. GASA5, a regulator of flowering time and stem growth in Arabidopsis thaliana. Plant Mol. Biol. 2009, 69, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Wang, H.; Yu, H.; Zhong, C.; Zhang, X.; Peng, J.; Wang, X. GASA14 regulates leaf expansion and abiotic stress resistance by modulating reactive oxygen species accumulation. J. Exp. Bot. 2013, 64, 1637–1647. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, J.I.; Amaya, I.; Castillejo, C.; Sevilla, J.F.S.; Quesada, M.A.; Botella, M.; Valpuesta, V. The strawberry gene FaGAST affects plant growth through inhibition of cell elongation. J. Exp. Bot. 2006, 57, 2401–2411. [Google Scholar] [CrossRef]
- Moyano-Cañete, E.; Bellido, M.L.; García-Caparrós, N.; Medina-Puche, L.; Amil-Ruiz, F.; González-Reyes, J.A.; Caballero, J.L.; Muñoz-Blanco, J.; Portales, R.B. FaGAST2, a Strawberry Ripening-Related Gene, Acts Together with FaGAST1 to Determine Cell Size of the Fruit Receptacle. Plant Cell Physiol. 2012, 54, 218–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, C.-B.; Woo, Y.-M.; Lee, D.J.; Lee, M.-C.; Kim, C.S. Enhanced tolerance to heat stress in transgenic plants expressing the GASA4 gene. Plant Physiol. Biochem. 2007, 45, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, X. Overexpression of GASA5 increases the sensitivity of Arabidopsis to heat stress. J. Plant Physiol. 2011, 168, 2093–2101. [Google Scholar] [CrossRef]
- Hou, D.; Bai, Q.; Li, J.; Xie, L.; Li, X.; Cheng, Z.; Gao, J. The Gibberellic Acid-Stimulated Transcript Gene Family in Moso Bamboo: A Genome-Wide Survey and Expression Profiling During Development and Abiotic Stresses. J. Plant Growth Regul. 2018, 37, 1135–1147. [Google Scholar] [CrossRef]
- Alonso-Ramírez, A.; Rodríguez, D.; Reyes, D.; Jiménez, J.A.; Nicolás, G.; López-Climent, M.; Gómez-Cadenas, A.; Nicolas, C. Evidence for a Role of Gibberellins in Salicylic Acid-Modulated Early Plant Responses to Abiotic Stress in Arabidopsis Seeds. Plant Physiol. 2009, 150, 1335–1344. [Google Scholar] [CrossRef] [Green Version]
- Segura, A.; Moreno, M.; Madueño, F.; Molina, A.; García-Olmedo, F. Snakin-1, a Peptide from Potato That Is Active Against Plant Pathogens. Mol. Plant-Microbe Interact. 1999, 12, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Berrocal-Lobo, M.; Segura, A.; Moreno, M.; López, G.; García-Olmedo, F.; Molina, A. Snakin-2, an Antimicrobial Peptide from Potato Whose Gene Is Locally Induced by Wounding and Responds to Pathogen Infection. Plant Physiol. 2002, 128, 951–961. [Google Scholar] [CrossRef] [Green Version]
- Faraji, S.; Mehmood, F.; Malik, H.; Ahmed, I.; Heidari, P.; Poczai, P. The GASA Gene Family in Cacao (Theobroma cacao, Malvaceae): Genome Wide Identification and Expression Analysis. Agronomy 2021, 11, 1425. [Google Scholar] [CrossRef]
- Ma, L.; Wang, Q.; Zheng, Y.; Guo, J.; Yuan, S.; Fu, A.; Bai, C.; Zhao, X.; Zheng, S.; Wen, C.; et al. Cucurbitaceae genome evolution, gene function, and molecular breeding. Hortic. Res. 2022, 9, uhab057. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wang, Q.; Mu, J.; Fu, A.; Wen, C.; Zhao, X.; Gao, L.; Li, J.; Shi, K.; Wang, Y.; et al. The genome and transcriptome analysis of snake gourd provide insights into its evolution and fruit development and ripening. Hortic. Res. 2020, 7, 199. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Nie, J.; Li, X.; Scanlon, M.J.; Zhang, C.; Zheng, Y.; Ma, S.; Shan, N.; Fei, Z.; Turgeon, R. Transcriptomic and functional analysis of cucumber (Cucumis sativus L.) fruit phloem during early development. Plant J. 2018, 96, 982–996. [Google Scholar] [CrossRef] [Green Version]
- Bhowmick, B.; Jha, S. Dynamics of sex expression and chromosome diversity in Cucurbitaceae: A story in the making. J. Genet. 2015, 94, 793–808. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Li, H.; Huang, W.; Xu, Y.; Zhou, Q.; Wang, S.; Ruan, J.; Huang, S.; Zhang, Z. A chromosome-scale genome assembly of cucumber (Cucumis sativus L.). GigaScience 2019, 8, giz072. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Mas, J.; Benjak, A.; Sanseverino, W.; Bourgeois, M.; Mir, G.; González, V.M.; Hénaff, E.; Câmara, F.; Cozzuto, L.; Lowy, E. The genome of melon (Cucumis melo L.). Proc. Natl. Acad. Sci. USA 2012, 109, 11872–11877. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Zhang, J.; Sun, H.; Salse, J.; Lucas, W.J.; Zhang, H.; Zheng, Y.; Mao, L.; Ren, Y.; Wang, Z.; et al. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Wu, S.; Zhang, G.; Jiao, C.; Guo, S.; Ren, Y.; Zhang, J.; Zhang, H.; Gong, G.; Jia, Z.; et al. Karyotype Stability and Unbiased Fractionation in the Paleo-Allotetraploid Cucurbita Genomes. Mol. Plant 2017, 10, 1293–1306. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.; Xu, Y.; Wang, J.; Liu, W.; Zhou, Q.; Luo, S.; Huang, W.; He, X.; Li, Q.; Peng, Q.; et al. The wax gourd genomes offer insights into the genetic diversity and ancestral cucurbit karyotype. Nat. Commun. 2019, 10, 5185. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Shamimuzzaman, M.D.; Sun, H.; Salse, J.; Sui, X.; Wilder, A.; Wu, Z.; Levi, A.; Xu, Y.; Ling, K.; et al. The bottle gourd genome provides insights into Cucurbitaceae evolution and facilitates mapping of a Papaya ring-spot virus resistance locus. Plant J. 2017, 92, 963–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pootakham, W.; Sonthirod, C.; Naktang, C.; Nawae, W.; Yoocha, T.; Kongkachana, W.; Sangsrakru, D.; Jomchai, N.; U-Thoomporn, S.; Sheedy, J.R.; et al. De novo assemblies of Luffa acutangula and Luffa cylindrica genomes reveal an expansion associated with substantial accumulation of transposable elements. Mol. Ecol. Resour. 2021, 21, 212–225. [Google Scholar] [CrossRef]
- Urasaki, N.; Takagi, H.; Natsume, S.; Uemura, A.; Taniai, N.; Miyagi, N.; Fukushima, M.; Suzuki, S.; Tarora, K.; Tamaki, M.; et al. Draft genome sequence of bitter gourd (Momordica charantia), a vegetable and medicinal plant in tropical and subtropical regions. DNA Res. 2017, 24, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, A.; Wang, Q.; Mu, J.; Ma, L.; Wen, C.; Zhao, X.; Gao, L.; Li, J.; Shi, K.; Wang, Y.; et al. Combined genomic, transcriptomic, and metabolomic analyses provide insights into chayote (Sechium edule) evolution and fruit development. Hortic. Res. 2021, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Jiang, W.; Zhang, Y.; Yu, H.; Mao, Z.; Gu, X.; Huang, S.; Xie, B. Genome-wide analysis of WRKY gene family in Cucumis sativus. BMC Genom. 2011, 12, 471. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Li, H.; Yang, Y.; Wang, Y.; Mo, Y.; Zhang, R.; Zhang, Y.; Ma, J.; Wei, C.; Zhang, X. Identification and expression analyses of WRKY genes reveal their involvement in growth and abiotic stress response in watermelon (Citrullus lanatus). PLoS ONE 2018, 13, e0191308. [Google Scholar] [CrossRef]
- Bankaji, I.; Sleimi, N.; Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Identification and expression of the Cucurbita WRKY transcription factors in response to water deficit and salt stress. Sci. Hortic. 2022, 256, 108562. [Google Scholar] [CrossRef]
- Chen, Y.; Jing, X.; Wang, S.; Wang, J.; Zhang, S.; Shi, Q. Genome-wide Analysis of WRKY Transcription Factor Family in Melon (Cucumis melo L.) and Their Response to Powdery Mildew. Plant Mol. Biol. Rep. 2021, 39, 686–699. [Google Scholar] [CrossRef]
- Hu, L.F.; Liu, S.Q. Genome-wide analysis of the MADS-box gene family in cucumber. Genome 2012, 55, 245–256. [Google Scholar] [CrossRef]
- Wang, P.; Wang, S.; Chen, Y.; Xu, X.; Guang, X.; Zhang, Y. Genome-wide Analysis of the MADS-Box Gene Family in Watermelon. Comput. Biol. Chem. 2019, 80, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Fu, Y.; Zhao, W.; Liu, L.; Bade, R.; Hasi, A.; Hao, J. Genome-wide Identification and Analysis of the MADS-box Gene Family in Melon. J. Am. Soc. Hortic. Sci. 2016, 141, 507–519. [Google Scholar] [CrossRef] [Green Version]
- Wan, H.; Yuan, W.; Bo, K.; Shen, J.; Pang, X.; Chen, J. Genome-wide analysis of NBS-encoding disease resistance genes in Cucumis sativus and phylogenetic study of NBS-encoding genes in Cucurbitaceae crops. BMC Genom. 2013, 14, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, Z.; Rahim, A.; Jung, H.-J.; Park, J.-I.; Kim, H.-T.; Nou, I.-S. Genome-Wide Characterization of NBS-Encoding Genes in Watermelon and Their Potential Association with Gummy Stem Blight Resistance. Int. J. Mol. Sci. 2019, 20, 902. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yuan, Q.; Wu, Y.; Zhang, J.; Nie, J. Genome-Wide Identification and Characterization of the CC-NBS-LRR Gene Family in Cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2022, 23, 5048. [Google Scholar] [CrossRef]
- Baloglu, M.C.; Eldem, V.; Hajyzadeh, M.; Unver, T. Genome-Wide Analysis of the bZIP Transcription Factors in Cucumber. PLoS ONE 2014, 9, e96014. [Google Scholar] [CrossRef] [Green Version]
- Unel, N.M.; Cetin, F.; Karaca, Y.; Altunoglu, Y.C.; Baloglu, M.C. Comparative identification, characterization, and expression analysis of bZIP gene family members in watermelon and melon genomes. Plant Growth Regul. 2019, 87, 227–243. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Yu, C.S.; Lin, C.J.; Hwang, J.K. Predicting subcellular localization of proteins for Gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.W.; Pirrung, M.; McCue, L.A. FQC Dashboard: Integrates FastQC results into a web-based, interactive, and extensible FASTQ quality control tool. Bioinformatics 2017, 33, 3137–3139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.-Y.; Dillies, M.-A. SARTools: A DESeq2- and EdgeR-based R pipeline for comprehensive differential analysis of RNA-Seq data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Z.; Yan, P.; Huang, S.; Fei, Z.; Lin, K. RNA-Seq improves annotation of protein-coding genes in the cucumber genome. BMC Genom. 2011, 12, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhao, G.; Li, Y.; Mo, N.; Zhang, J.; Liang, Y. Transcriptomic Analysis Implies That GA Regulates Sex Expression via Ethylene-Dependent and Ethylene-Independent Pathways in Cucumber (Cucumis sativus L.). Front. Plant Sci. 2017, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, X.; Han, J.; Lu, W.; Ren, Z. Genome-wide analysis of the WRKY gene family in the cucumber genome and transcriptome-wide identification of WRKY transcription factors that respond to biotic and abiotic stresses. BMC Plant Biol. 2020, 20, 443. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yin, J.; Liang, Y.; Liu, J.; Jia, J.; Huo, H.; Wu, Z.; Yang, R.; Gong, H. Transcriptomic dynamics provide an insight into the mechanism for silicon-mediated alleviation of salt stress in cucumber plants. Ecotoxicol. Environ. Saf. 2019, 174, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, A.; Day, B. Transcriptome and Small RNAome Dynamics during a Resistant and Susceptible Interaction between Cucumber and Downy Mildew. Plant Genome 2016, 9, 69. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Xu, X.; Shi, Y.; Qi, X.; Chen, X. Elucidation of the molecular responses of a cucumber segment substitution line carrying Pm5.1 and its recurrent parent triggered by powdery mildew by comparative transcriptome profiling. BMC Genom. 2017, 18, 21. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cheng, C.; Zhang, K.; Tian, Z.; Xu, J.; Yang, S.; Lou, Q.; Li, J.; Chen, J.-F. Comparative transcriptomics reveals suppressed expression of genes related to auxin and the cell cycle contributes to the resistance of cucumber against Meloidogyne incognita. BMC Genom. 2018, 19, 583. [Google Scholar] [CrossRef]
- Sun, Y.; Shang, L.; Zhu, Q.-H.; Fan, L.; Guo, L. Twenty years of plant genome sequencing: Achievements and challenges. Trends Plant Sci. 2021, 27, 391–401. [Google Scholar] [CrossRef]
- Wu, K.; Qu, Y.; Rong, H.; Han, X.; Tian, Y.; Xu, L. Identification and Expression Analysis of the Populus trichocarpa GASA-Gene Family. Int. J. Mol. Sci. 2022, 23, 1507. [Google Scholar] [CrossRef]
- Muhammad, I.; Li, W.-Q.; Jing, X.-Q.; Zhou, M.-R.; Shalmani, A.; Ali, M.; Wei, X.-Y.; Sharif, R.; Liu, W.-T.; Chen, K.-M. A systematic in silico prediction of gibberellic acid stimulated GASA family members: A novel small peptide contributes to floral architecture and transcriptomic changes induced by external stimuli in rice. J. Plant Physiol. 2019, 234–235, 117–132. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, C.; Li, L.; Meng, L.; Singh, J.; Jiang, N.; Deng, X.-W.; He, Z.-H.; Lemaux, P.G. Evolutionary Expansion, Gene Structure, and Expression of the Rice Wall-Associated Kinase Gene Family. Plant Physiol. 2005, 139, 1107–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, M.; Tyagi, A.K.; Khurana, J.P. Genome-wide analysis, evolutionary expansion, and expression of early auxin-responsive SAUR gene family in rice (Oryza sativa). Genomics 2006, 88, 360–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Yu, S.; Liu, D.; Xin, M.; Zhou, X. Cloning and bioinformatics analysis of CsGASA4 gene in cucumber. J. Northeast. Agric. Univ. 2018, 49, 19–26. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene ID | Chromosome Location | Number of Amino Acid (aa) | Molecular Weight (kDa) | Theoretical pI | Instability Index | Aliphatic Index | Grand Average of Hydropathicity | Subcellular Location |
|---|---|---|---|---|---|---|---|---|---|
| Wax gourd (Benincasa hispida) | Bhi03M000750 | Chr3: 19,311,494 .. 19,311,920 (−) | 80 | 8.77 | 9.04 | 39.58 | 82.88 | −0.021 | Extracellular |
| Bhi04M000133 | Chr4: 3,757,574 .. 3,758,363 (+) | 97 | 10.33 | 9.21 | 53.79 | 68.45 | 0.073 | Extracellular | |
| Bhi04M000374 | Chr4: 10,323,755 .. 10,323,929 (−) | 57 | 6.15 | 9.33 | 32.81 | 22.28 | −0.749 | Nuclear | |
| Bhi08M001209 | Chr8: 42,837,600 .. 42,840,156 (+) | 115 | 12.91 | 8.91 | 44.95 | 59.30 | −0.355 | Extracellular | |
| Bhi09M001786 | Chr9: 59,471,263 .. 59,472,200 (+) | 139 | 15.00 | 9.10 | 55.67 | 89.14 | 0.310 | Extracellular | |
| Bhi09M001788 | Chr9: 59,495,483 .. 59,496,419 (+) | 101 | 11.01 | 7.99 | 43.64 | 65.74 | 0.031 | Extracellular | |
| Bhi11M000974 | Chr11: 34,037,800 .. 34,038,470 (+) | 89 | 10.08 | 9.40 | 32.31 | 63.48 | 0.111 | Extracellular | |
| Bhi11M001327 | Chr11: 45,095,365 .. 45,097,267 (−) | 133 | 14.80 | 8.97 | 64.51 | 61.58 | −0.074 | Extracellular | |
| Bhi11M002565 | Chr11: 83,283,688 .. 83,287,470 (−) | 232 | 24.48 | 9.84 | 89.32 | 57.93 | −0.311 | Nuclear | |
| Watermelon (Citrullus lanatus) | Cla97C01G013050.1 | Chr1: 26,879,226 .. 26,881,027 (−) | 114 | 12.72 | 8.97 | 45.94 | 56.40 | −0.393 | Extracellular |
| Cla97C01G016400.2 | Chr1: 30,120,921 .. 30,122,856 (−) | 99 | 10.84 | 9.24 | 52.87 | 70.91 | −0.159 | Extracellular | |
| Cla97C02G033610.2 | Chr2: 7,155,664 .. 7,157,420 (+) | 88 | 10.11 | 9.62 | 33.11 | 59.89 | −0.086 | Extracellular | |
| Cla97C05G099600.2 | Chr5: 28,631,501 .. 28,637,708 (+) | 216 | 23.23 | 8.76 | 45.58 | 65.51 | −0.024 | Extracellular | |
| Cla97C08G157880.1 | Chr8: 25,344,837 .. 25,345,286 (+) | 97 | 10.18 | 9.07 | 45.74 | 78.56 | 0.187 | Extracellular | |
| Cla97C10G193530.1 | Chr10: 21,997,173 .. 21,997,578 (+) | 89 | 10.08 | 9.15 | 37.27 | 53.71 | −0.054 | Extracellular | |
| Cla97C10G196435.1 | Chr10: 26,191,272 .. 26,194,258 (−) | 141 | 15.55 | 9.07 | 66.45 | 45.67 | −0.453 | Extracellular | |
| Cla97C10G197380.1 | Chr10: 27,180,180 .. 27,181,843 (+) | 216 | 22.78 | 9.68 | 89.57 | 54.58 | −0.385 | Nuclear | |
| Cla97C11G215390.1 | Chr11: 11,861,823 .. 11,863,862 (−) | 106 | 11.78 | 9.46 | 50.63 | 71.79 | −0.183 | Extracellular | |
| Pumpkin (Cucurbita moschata) | CmoCh01G020290.1 | Chr1: 14,246,135 .. 14,247,088 (−) | 106 | 10.90 | 8.61 | 54.84 | 81.04 | 0.235 | Extracellular |
| CmoCh05G001560.1 | Chr5: 678,878 .. 679,725 (+) | 85 | 9.18 | 9.19 | 32.68 | 74.59 | 0.054 | Extracellular | |
| CmoCh07G006840.1 | Chr7: 3,085,589 .. 3,086,076 (+) | 94 | 10.04 | 8.97 | 43.34 | 71.60 | −0.068 | Extracellular | |
| CmoCh09G000990.1 | Chr9: 477,271 .. 484,805 (+) | 516 | 58.17 | 5.99 | 54.74 | 85.41 | −0.103 | Nuclear | |
| CmoCh11G010460.1 | Chr11: 5,816,242 .. 5,817,492 (−) | 108 | 11.89 | 9.45 | 36.93 | 50.65 | −0.201 | Extracellular | |
| CmoCh11G014010.1 | Chr11: 9,853,936 .. 9,855,463 (−) | 88 | 9.86 | 9.63 | 29.27 | 65.45 | 0.036 | Extracellular | |
| CmoCh12G003610.1 | Chr12: 2,236,117 .. 2,237,074 (+) | 97 | 10.16 | 9.08 | 53.11 | 75.57 | 0.186 | Extracellular | |
| CmoCh14G006410.1 | Chr14: 3,223,478 .. 3,225,060 (−) | 203 | 21.47 | 9.82 | 84.49 | 60.00 | −0.342 | Nuclear | |
| CmoCh14G011310.1 | Chr14: 7,958,822 .. 7,959,972 (−) | 117 | 12.79 | 8.82 | 50.21 | 49.23 | −0.406 | Extracellular | |
| CmoCh15G000310.1 | Chr15: 218,843 .. 220,014 (−) | 106 | 11.90 | 9.59 | 64.32 | 71.79 | −0.166 | Extracellular | |
| CmoCh18G001170.1 | Chr18: 823,704 .. 825,231 (+) | 115 | 12.22 | 9.36 | 51.06 | 54.26 | −0.184 | Extracellular | |
| Cucumber (Cucumis sativus) | CsaV3_1G008270.1 | Chr1: 5,210,806 .. 5,212,803 (+) | 115 | 13.05 | 9.19 | 44.35 | 53.39 | −0.414 | Extracellular |
| CsaV3_2G026300.1 | Chr2: 18,030,665 .. 18,031,342 (+) | 85 | 9.47 | 9.20 | 34.97 | 64.24 | −0.171 | Extracellular | |
| CsaV3_2G029490.1 | Chr2: 19,316,885 .. 19,317,559 (−) | 61 | 6.59 | 9.11 | 56.39 | 36.89 | −0.408 | Nuclear | |
| CsaV3_3G041480.1 | Chr3: 33,839,191 .. 33,840,892 (−) | 104 | 11.46 | 9.64 | 34.12 | 54.52 | −0.183 | Extracellular | |
| CsaV3_3G042060.1 | Chr3: 34,217,470 .. 34,219,983 (−) | 231 | 24.24 | 9.77 | 84.05 | 67.06 | −0.263 | Nuclear | |
| CsaV3_3G042990.1 | Chr3: 34,899,642 .. 34,900,907 (+) | 117 | 12.94 | 9.12 | 54.69 | 61.62 | −0.215 | Extracellular | |
| CsaV3_3G045860.1 | Chr3: 37,493,613 .. 37,494,832 (−) | 102 | 10.59 | 8.57 | 60.59 | 76.57 | 0.249 | Extracellular | |
| CsaV3_4G000520.1 | Chr4: 298,937 .. 299,297 (+) | 89 | 10.16 | 9.41 | 30.95 | 59.10 | −0.115 | Extracellular | |
| CsaV3_6G043760.1 | Chr6: 25,805,953 .. 25,806,385 (−) | 80 | 8.82 | 9.15 | 37.53 | 70.62 | −0.148 | Extracellular | |
| Sponge gourd (Luffa cylindrica) | Lcy01g018340.1 | Chr1: 39,876,992 .. 39,877,586 (−) | 85 | 9.39 | 9.27 | 26.87 | 63.06 | −0.111 | Extracellular |
| Lcy04g015090.1 | Chr4: 47,422,624 .. 47,423,017 (−) | 94 | 10.08 | 8.87 | 46.90 | 75.64 | 0.007 | Extracellular | |
| Lcy06g003590.1 | Chr6: 3,388,158 .. 3,389,543 (+) | 114 | 12.74 | 8.88 | 54.68 | 55.53 | −0.361 | Extracellular | |
| Lcy07g018230.1 | Chr7: 46,850,291 .. 46,852,453 (−) | 109 | 11.92 | 9.57 | 45.40 | 52.94 | −0.043 | Extracellular | |
| Lcy09g000520.1 | Chr9: 508,156 .. 510,023 (−) | 109 | 12.19 | 9.61 | 50.43 | 68.07 | −0.186 | Extracellular | |
| Lcy09g017660.1 | Chr9: 35,628,176 .. 35,629,130 (−) | 106 | 11.81 | 9.67 | 32.92 | 51.60 | −0.288 | Extracellular | |
| Lcy09g018840.1 | Chr9: 41,976,742 .. 41,978,205 (−) | 221 | 23.44 | 9.79 | 80.57 | 58.19 | −0.388 | Nuclear | |
| Lcy12g021390.1 | Chr12: 48,438,825 .. 48,439,301 (−) | 101 | 10.80 | 8.29 | 40.87 | 82.08 | 0.156 | Extracellular | |
| Lcy12g021440.1 | Chr12: 48,464,326 .. 48,464,817 (−) | 106 | 11.42 | 8.65 | 44.30 | 79.15 | 0.072 | Extracellular | |
| Bottle gourd (Lagenaria siceraria) | Lsi01G009320.1 | Chr1: 7,534,930 .. 7,535,476 (+) | 80 | 8.82 | 9.04 | 37.17 | 75.50 | −0.077 | Extracellular |
| Lsi03G005590.1 | Chr3: 6,743,934 .. 6,745,226 (−) | 100 | 11.34 | 9.47 | 38.43 | 65.30 | 0.034 | Extracellular | |
| Lsi03G010390.1 | Chr3: 19,955,854 .. 19,956,253 (+) | 89 | 10.18 | 9.33 | 34.28 | 59.10 | −0.003 | Extracellular | |
| Lsi03G014250.1 | Chr3: 25,218,255 .. 25,221,595 (+) | 212 | 22.55 | 9.45 | 92.37 | 51.42 | −0.493 | Nuclear | |
| Lsi04G017450.1 | Chr4: 24,750,180 .. 24,751,157 (−) | 111 | 11.86 | 8.81 | 48.82 | 72.07 | 0.142 | Extracellular | |
| Lsi08G013250.1 | Chr8: 21,566,057 .. 21,567,220 (+) | 147 | 15.87 | 9.36 | 51.98 | 82.38 | 0.149 | Extracellular | |
| Lsi08G015100.1 | Chr8: 23,193,981 .. 23,195,157 (−) | 142 | 15.46 | 8.98 | 23.27 | 66.62 | −0.246 | Extracellular | |
| Lsi09G009210.1 | Chr9: 11,377,348 .. 11,377,967 (+) | 82 | 9.37 | 9.50 | 22.47 | 40.37 | −0.466 | Extracellular | |
| Bitter gourd (Momordica charantia) | MC01g0899.1 | Chr1: 14,806,676 .. 14,807,137 (−) | 95 | 10.15 | 8.87 | 39.92 | 78.95 | 0.022 | Extracellular |
| MC02g0120.1 | Chr2: 1,223,343 .. 1,224,955 (+) | 108 | 11.64 | 8.89 | 51.46 | 51.67 | −0.060 | Extracellular | |
| MC03g1060.1 | Chr3: 16,943,540 .. 16,943,725 (−) | 62 | 6.65 | 9.32 | 46.78 | 31.45 | −0.556 | Nuclear | |
| MC04g0594.1 | Chr4: 5,232,091 .. 5,232,267 (−) | 59 | 6.78 | 9.84 | 61.72 | 29.83 | −0.764 | Nuclear | |
| MC04g1197.1 | Chr4: 19,993,730 .. 19,993,924 (+) | 65 | 7.30 | 9.00 | 26.91 | 24.00 | −0.606 | Extracellular | |
| MC04g1983.1 | Chr4: 26,610,157 .. 26,612,959 (+) | 109 | 12.18 | 9.75 | 67.07 | 71.65 | −0.229 | Extracellular | |
| MC04g_new0671.1 | Chr4: 26,612,371 .. 26,616,386 (−) | 370 | 41.55 | 8.95 | 44.27 | 74.08 | −0.235 | Extracellular | |
| MC08g0576.1 | Chr8: 4,660,922 .. 4,661,179 (+) | 86 | 9.83 | 9.03 | 31.67 | 62.33 | −0.073 | Extracellular | |
| MC08g0741.1 | Chr8: 6,030,387 .. 6,030,572 (−) | 62 | 6.49 | 8.98 | 43.72 | 44.19 | −0.285 | Extracellular | |
| MC08g1334.1 | Chr8: 12,401,247 .. 12,401,444 (+) | 66 | 7.58 | 9.03 | 36.59 | 56.06 | −0.459 | Extracellular | |
| MC08g1395.1 | Chr8: 14,414,051 .. 14,414,239 (+) | 63 | 7.16 | 9.13 | 45.20 | 16.98 | −0.767 | Nuclear | |
| MC10g0377.1 | Chr10: 3,011,243 .. 3,011,419 (+) | 59 | 6.61 | 9.18 | 34.38 | 24.75 | −0.781 | Nuclear | |
| MC11g0104.1 | Chr11: 764,783 .. 764,971 (+) | 63 | 6.49 | 8.89 | 75.97 | 43.49 | −0.068 | Extracellular | |
| MC11g0105.1 | Chr11: 765,959 .. 768,132 (+) | 153 | 16.95 | 8.65 | 54.36 | 85.42 | 0.065 | Extracellular | |
| MC00g0794.1 | scaffold142: 23,557 .. 24,251 (−) | 116 | 12.96 | 9.10 | 55.48 | 47.07 | −0.324 | Extracellular | |
| Melon (Cucumis melo) | MELO3C018503.2.1 | Chr1: 721,831 .. 722,928 (−) | 88 | 10.13 | 9.71 | 30.06 | 61.02 | −0.174 | Extracellular |
| MELO3C011008.2.1 | Chr3: 27,015,343 .. 27,016,361 (+) | 97 | 10.47 | 9.34 | 59.97 | 75.46 | 0.080 | Extracellular | |
| MELO3C010822.2.1 | Chr3: 28,229,740 .. 28,230,991 (−) | 85 | 9.44 | 9.30 | 33.74 | 67.65 | −0.080 | Extracellular | |
| MELO3C003753.2.1 | Chr4: 3,727,349 .. 3,728,176 (+) | 100 | 10.81 | 8.65 | 45.75 | 70.30 | 0.079 | Extracellular | |
| MELO3C026601.2.1 | Chr4: 25,918,328 .. 25,919,317 (−) | 117 | 12.95 | 9.12 | 52.40 | 57.52 | −0.302 | Extracellular | |
| MELO3C009926.2.1 | Chr4: 26,961,415 .. 26,964,335 (+) | 222 | 23.64 | 9.72 | 93.63 | 55.72 | −0.386 | Nuclear | |
| MELO3C009872.2.1 | Chr4: 27,411,710 .. 27,412,273 (+) | 80 | 8.93 | 9.55 | 34.32 | 23.12 | −0.761 | Extracellular | |
| MELO3C017982.2.1 | Chr7: 26,076,528 .. 26,081,332 (−) | 89 | 10.21 | 9.36 | 35.50 | 50.34 | −0.134 | Extracellular | |
| MELO3C007600.2.1 | Chr8: 3,965,613 .. 3,966,256 (−) | 94 | 10.19 | 8.96 | 38.10 | 67.34 | −0.077 | Extracellular | |
| MELO3C002243.2.1 | Chr12: 23,960,061 .. 23,961,357 (−) | 126 | 14.28 | 9.07 | 52.56 | 58.81 | −0.456 | Extracellular | |
| Chayote (Sechium edule) | Sed0003696.1 | Chr1: 12,188,492 .. 12,189,979 (−) | 112 | 12.57 | 8.87 | 49.75 | 54.02 | −0.271 | Extracellular |
| Sed0004582.1 | Chr1: 18,633,871 .. 18,636,202 (−) | 108 | 11.53 | 8.79 | 41.99 | 66.02 | 0.053 | Extracellular | |
| Sed0003671.1 | Chr1: 60,857,945 .. 60,859,823 (−) | 109 | 11.98 | 8.47 | 62.21 | 57.43 | −0.079 | Extracellular | |
| Sed0018489.1 | Chr3: 4,097,163 .. 4,099,736 (+) | 109 | 12.02 | 9.33 | 50.58 | 64.50 | −0.239 | Extracellular | |
| Sed0001116.1 | Chr3: 5,503,219 .. 5,504,432 (−) | 95 | 10.71 | 9.01 | 26.56 | 61.58 | 0.155 | Extracellular | |
| Sed0023008.1 | Chr4: 35,040,789 .. 35,043,255 (−) | 115 | 13.11 | 9.17 | 55.32 | 61.91 | −0.248 | Extracellular | |
| Sed0019149.1 | Chr6: 5,108,016 .. 5,110,361 (+) | 223 | 23.49 | 9.57 | 89.94 | 70.36 | −0.114 | Nuclear | |
| Sed0023122.1 | Chr6: 5,770,438 .. 5,772,489 (+) | 104 | 11.48 | 9.61 | 31.14 | 57.21 | −0.129 | Extracellular | |
| Sed0021074.1 | Chr8: 27,782,007 .. 27,783,423 (−) | 106 | 11.91 | 9.78 | 43.65 | 54.34 | −0.238 | Extracellular | |
| Sed0023101.1 | Chr10: 25,350,679 .. 25,351,022 (+) | 86 | 9.64 | 8.87 | 23.26 | 62.33 | −0.071 | Extracellular | |
| Sed0017754.1 | Chr10: 36,389,392 .. 36,391,330 (−) | 88 | 10.01 | 9.62 | 36.59 | 62.05 | −0.042 | Extracellular | |
| Sed0026538.1 | Chr11: 26,467,504 .. 26,468,286 (−) | 98 | 10.52 | 8.81 | 56.25 | 72.76 | 0.095 | Extracellular | |
| Sed0005060.1 | Chr12: 7,036,681 .. 7,038,107 (+) | 111 | 12.46 | 9.16 | 71.39 | 57.93 | −0.377 | Extracellular | |
| Sed0023421.1 | Chr13: 20,866,691 .. 20,868,746 (+) | 107 | 11.16 | 8.24 | 49.56 | 79.25 | 0.396 | Extracellular | |
| Sed0020400.1 | Chr13: 20,881,190 .. 20,882,081 (+) | 106 | 11.37 | 8.49 | 37.98 | 72.74 | −0.007 | Extracellular | |
| Sed0017657.1 | Chr14: 17,756,540 .. 17,757,298 (−) | 96 | 10.35 | 8.97 | 27.91 | 71.98 | −0.072 | Extracellular | |
| Snake gourd (Trichosanthes anguina) | Tan0003426.1 | Chr1: 6,253,789 .. 6,254,353 (+) | 89 | 10.18 | 9.42 | 27.92 | 59.10 | 0.010 | Extracellular |
| Tan0008780.1 | Chr1: 115,842,003 .. 115,843,341 (−) | 96 | 10.50 | 8.64 | 47.66 | 87.40 | 0.329 | Extracellular | |
| Tan0000180.1 | Chr1: 115,844,096 .. 115,847,359 (−) | 104 | 11.06 | 8.86 | 49.88 | 72.31 | 0.176 | Extracellular | |
| Tan0005239.1 | Chr1: 115,863,453 .. 115,866,091 (−) | 103 | 10.95 | 8.92 | 52.45 | 83.40 | 0.235 | Extracellular | |
| Tan0020894.1 | Chr2: 14,079,327 .. 14,080,681 (−) | 89 | 10.29 | 9.68 | 44.19 | 63.60 | −0.048 | Extracellular | |
| Tan0006783.1 | Chr2: 81,798,300 .. 81,799,073 (+) | 85 | 9.32 | 9.27 | 28.27 | 64.24 | −0.115 | Extracellular | |
| Tan0018763.1 | Chr2: 90,173,196 .. 90,174,289 (−) | 92 | 9.87 | 9.19 | 40.27 | 63.70 | 0.142 | Extracellular | |
| Tan0011393.1 | Chr3: 72,374,679 .. 72,375,090 (−) | 80 | 8.73 | 9.17 | 32.55 | 74.25 | −0.138 | Extracellular | |
| Tan0001400.1 | Chr5: 70,569,691 .. 70,571,409 (−) | 111 | 12.50 | 8.60 | 62.49 | 57.03 | −0.353 | Extracellular | |
| Tan0005034.1 | Chr6: 74,131,427 .. 74,134,375 (+) | 116 | 13.00 | 9.29 | 49.14 | 56.47 | −0.337 | Extracellular | |
| Tan0020433.1 | Chr7: 12,409,140 .. 12,411,750 (+) | 114 | 12.98 | 9.90 | 55.96 | 43.68 | −0.402 | Extracellular | |
| Tan0017829.1 | Chr7: 48,550,357 .. 48,551,678 (−) | 106 | 11.70 | 9.61 | 25.70 | 50.66 | −0.282 | Extracellular | |
| Tan0016087.1 | Chr7: 61,289,275 .. 61,291,288 (−) | 234 | 25.13 | 9.75 | 80.79 | 54.49 | −0.357 | Nuclear | |
| Tan0009191.1 | Chr10: 27,664,301 .. 27,664,945 (+) | 103 | 11.45 | 8.87 | 69.36 | 41.65 | −0.545 | Extracellular | |
| Tan0010233.1 | Chr11: 862,587 .. 863,083 (+) | 95 | 10.55 | 9.09 | 39.86 | 63.68 | −0.161 | Extracellular | |
| Tan0017564.1 | Chr11: 863,623 .. 865,822 (−) | 109 | 12.17 | 9.48 | 50.35 | 77.80 | −0.024 | Extracellular | |
| Tan0010840.1 | Chr11: 12,544,995 .. 12,546,455 (−) | 95 | 10.88 | 9.30 | 33.22 | 45.26 | −0.180 | Extracellular | |
| Tan0013993.1 | Contig00035: 215,038 .. 215,449 (−) | 94 | 10.13 | 8.86 | 38.19 | 69.36 | −0.170 | Extracellular |
| Motif | Sequence | Number of Amino Acid | Pfam Annotation |
|---|---|---|---|
| motif 1 | CMRACGTCCARCKCVPPGTYGNKEVCP | 27 | GASA (Significant) |
| motif 2 | QPIDCGGACARRCSKASRKKR | 21 | GASA (Insignificant) |
| motif 3 | CYABMTTH | 8 | - |
| motif 4 | PPVKPPYT | 8 | - |
| motif 5 | LLSSLLLLLSFLDSS | 15 | - |
| motif 6 | GRPKCP | 6 | - |
| motif 7 | MKYCRICCSKCKCVP | 15 | GASA (Insignificant) |
| motif 8 | ANGLSQEKDAVYPHVPVPVPA | 21 | - |
| motif 9 | SGQMVITTQVDNPLP | 15 | - |
| motif 10 | MVNSIDGVAABPVKI | 15 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.; Hu, Y.; Yang, D.; Yan, C.; Li, N.; Li, Z.; Njogu, M.K.; Wang, X.; Jia, L. Genome-Wide Identification of GASA Gene Family in Ten Cucurbitaceae Species and Expression Analysis in Cucumber. Agronomy 2022, 12, 1978. https://doi.org/10.3390/agronomy12081978
Zhang K, Hu Y, Yang D, Yan C, Li N, Li Z, Njogu MK, Wang X, Jia L. Genome-Wide Identification of GASA Gene Family in Ten Cucurbitaceae Species and Expression Analysis in Cucumber. Agronomy. 2022; 12(8):1978. https://doi.org/10.3390/agronomy12081978
Chicago/Turabian StyleZhang, Kaijing, Yuchao Hu, Dekun Yang, Congsheng Yan, Nanyang Li, Ziang Li, Martin Kagiki Njogu, Xing Wang, and Li Jia. 2022. "Genome-Wide Identification of GASA Gene Family in Ten Cucurbitaceae Species and Expression Analysis in Cucumber" Agronomy 12, no. 8: 1978. https://doi.org/10.3390/agronomy12081978