
Seed Soaking with Sodium Selenate as a Biofortification Approach in Bread Wheat: Effects on Germination, Seedling Emergence, Biomass and Responses to Water Deficit

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Germination Experiment
2.2.1. Electrolyte Leakage
2.2.2. Selenium Determination
2.3. Pot Trial Description
2.4. Physiological Assay
2.4.1. Gas Exchange Parameters
2.4.2. Chlorophyll a Fluorescence
2.5. Biochemical Assay
2.5.1. Photosynthetic Pigments Quantification
2.5.2. Total Soluble Sugars and Proteins
2.5.3. Phenolic Composition Extration
2.5.4. Total Phenolic Content
2.5.5. Total Flavonoids Content
2.5.6. Ortho-Diphenols Content
2.5.7. Antioxidant Activity—ABTS+
2.6. Morphology and Yield Assessment
2.7. Statistical Analysis
3. Results
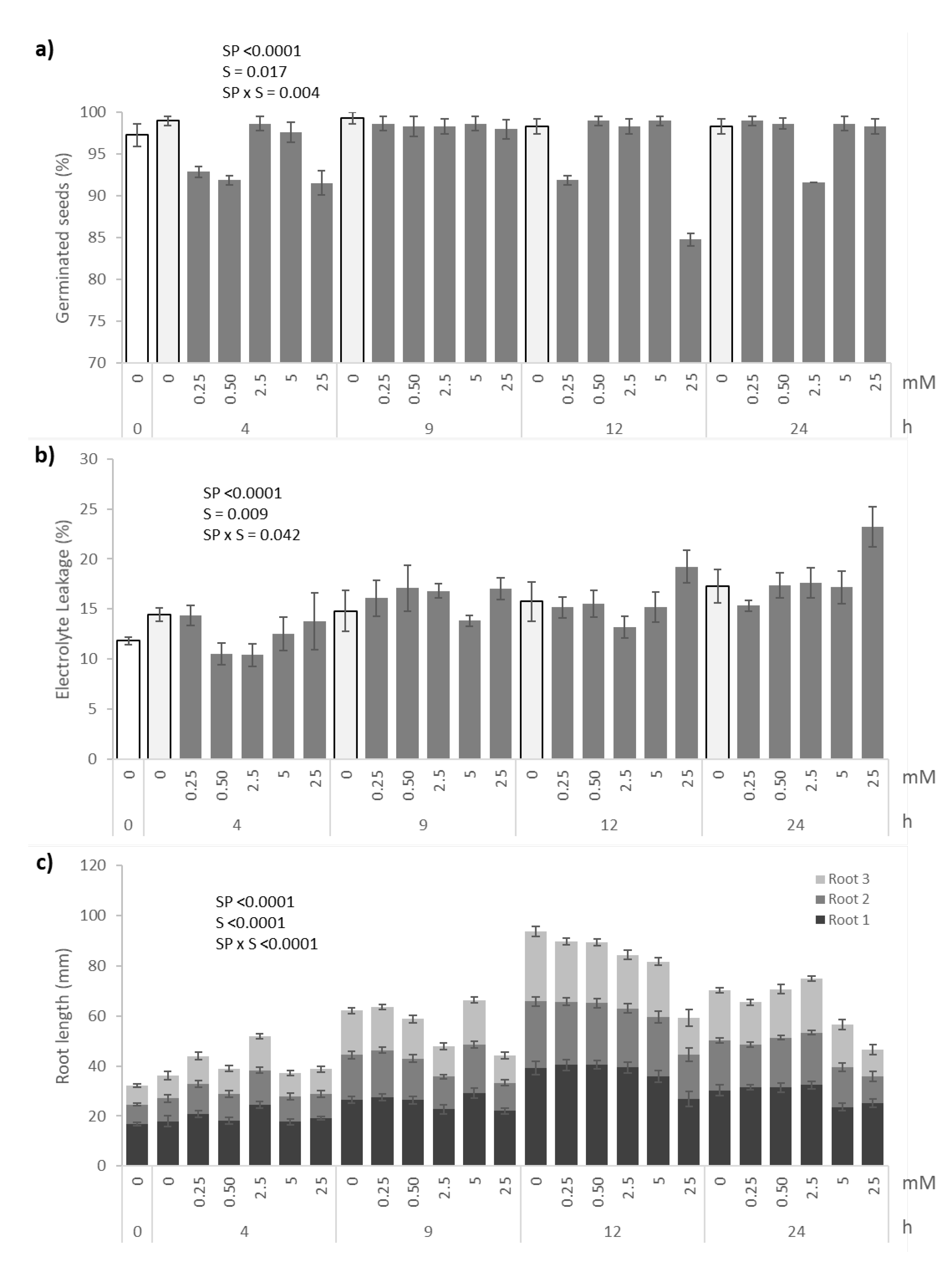
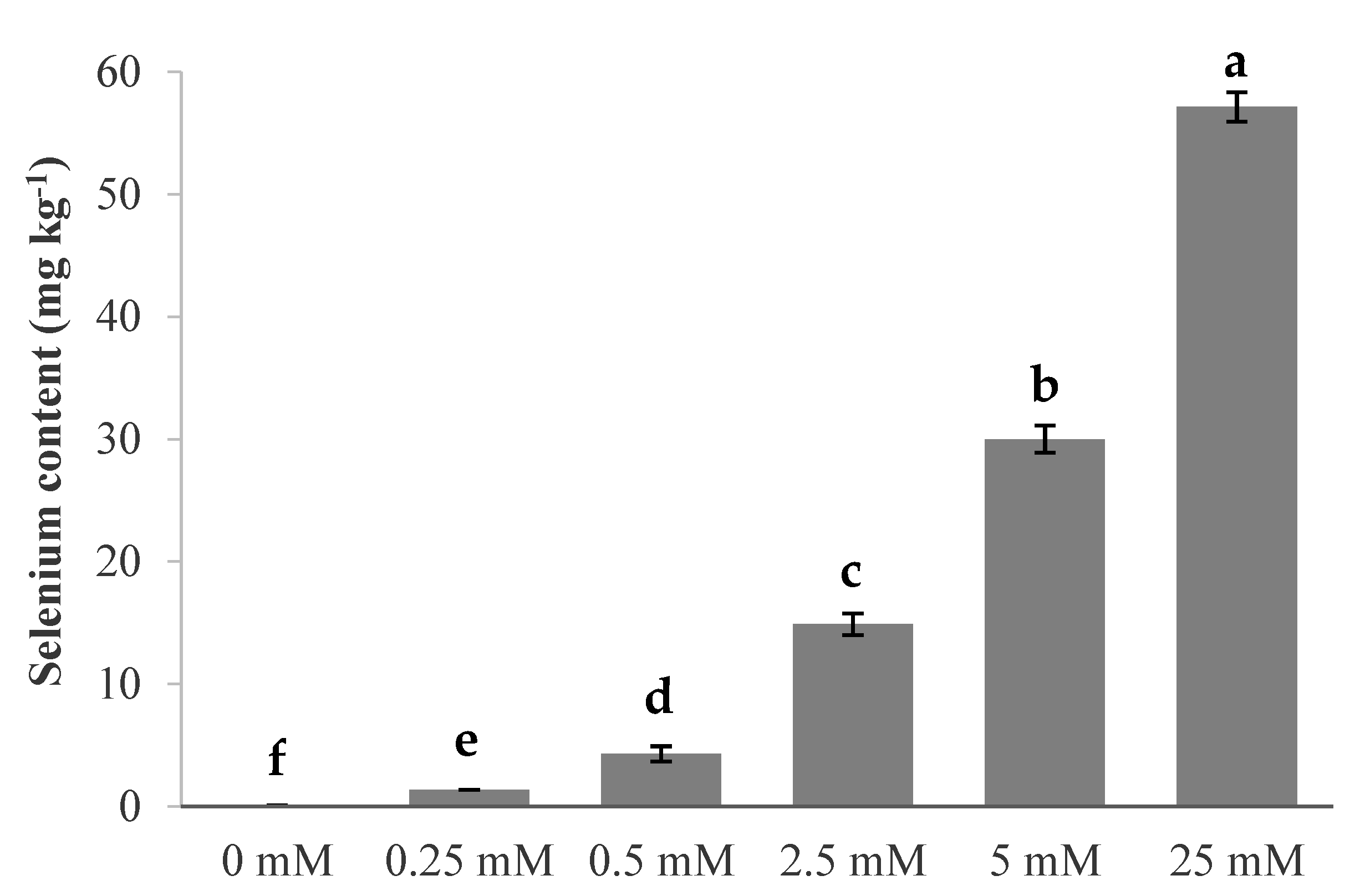
3.1. Germination, Electrolyte Leakage, Root Growth and Selenium Content
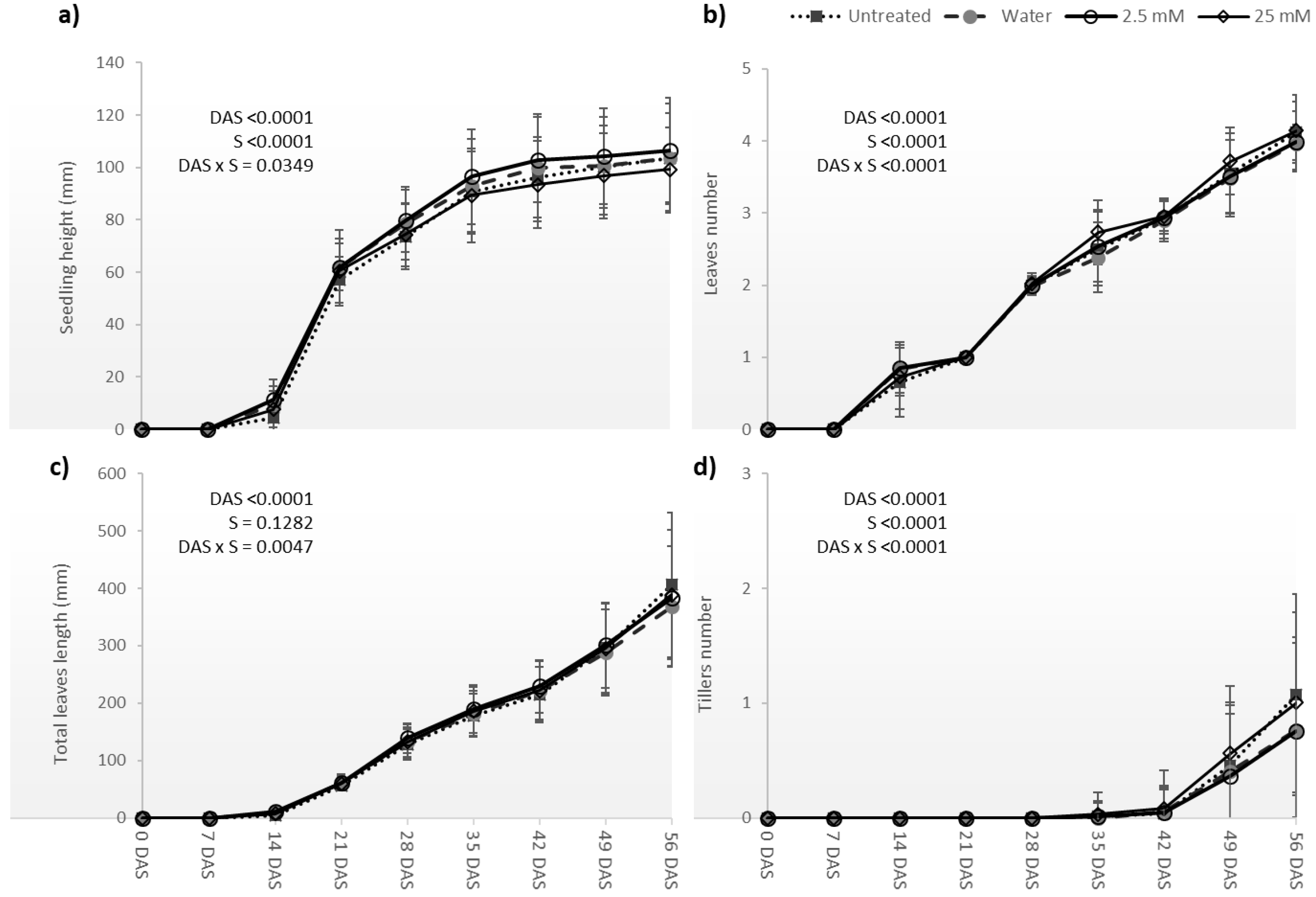
3.2. Germination, Emergence and Plant Development of Pot Trial
3.3. Leaf Gas Exchange and Chlorophyll a Fluorescence
3.4. Biochemical Analysis of Leaf
3.5. Straw and Grain Biomass Production
4. Discussion
4.1. Germination, Electrolyte Leakage, Root Growth, and Se Content
4.2. Coleoptile Emergence and Plant Growth before Stress Imposition
4.3. Physiological, Biochemical and Productivity Responses upon Water Deficit
4.3.1. Leaf Gas Exchange, Chlorophyll a Fluorescence Traits
4.3.2. Biochemical Analysis of Leaf
4.4. Straw and Grain Biomass Parameters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernandes, K.F.M.; Berton, R.; Coscione, A. Selenium biofortification of rice and radish: Effect of soil texture and efficiency of two extractants. Plant Soil Environ. 2014, 60, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Lyons, G.; Stangoulis, J.; Graham, R. High-selenium wheat: Biofortification for better health. Nutr. Res. Rev. 2003, 16, 45–60. [Google Scholar] [CrossRef] [Green Version]
- Puccinelli, M.; Pezzarossa, B.; Rosellini, I.; Malorgio, F. Selenium enrichment enhances the quality and shelf life of Basil leaves. Plants 2020, 9, 801. [Google Scholar] [CrossRef]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Roman Viñas, B.; Ribas Barba, L.; Ngo, J.; Gurinovic, M.; Novakovic, R.; Cavelaars, A.; de Groot, L.; van’t Veer, P.; Matthys, C.; Serra Majem, L. Projected prevalence of inadequate nutrient intakes in Europe. Ann. Nutr. Metab. 2011, 59, 84–95. [Google Scholar] [CrossRef]
- Jiang, Y.; Zeng, Z.H.; Bu, Y.; Ren, C.Z.; Li, J.Z.; Han, J.J.; Tao, C.; Zhang, K.; Wang, X.X.; Lu, G.X.; et al. Effects of selenium fertilizer on grain yield, Se uptake and distribution in common buckwheat (Fagopyrum esculentum Moench). Plant Soil Environ. 2016, 61, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Feghhenabi, F.; Hadi, H.; Khodaverdiloo, H.; van Genuchten, M.T. Seed priming alleviated salinity stress during germination and emergence of wheat (Triticum aestivum L.). Agric. Water Manag. 2020, 231, 106002. [Google Scholar] [CrossRef]
- Ali, Q.; Daud, M.K.; Haider, M.Z.; Ali, S.; Rizwan, M.; Aslam, N.; Noman, A.; Iqbal, N.; Shahzad, F.; Deeba, F.; et al. Seed priming by sodium nitroprusside improves salt tolerance in wheat (Triticum aestivum L.) by enhancing physiological and biochemical parameters. Plant Physiol. Biochem. 2017, 119, 50–58. [Google Scholar] [CrossRef]
- Farooq, M.; Usman, M.; Nadeem, F.; Rehman, H.u.; Wahid, A.; Basra, S.M.A.; Siddique, K.H.M. Seed priming in field crops: Potential benefits, adoption and challenges. Crop Pasture Sci. 2019, 70, 731–771. [Google Scholar] [CrossRef]
- Jiménez-Arias, D.; Pérez, J.A.; Luis, J.C.; Martín-Rodríguez, V.; Valdés-González, F.; Borges, A.A. Treating seeds in menadione sodium bisulphite primes salt tolerance in Arabidopsis by inducing an earlier plant adaptation. Environ. Exp. Bot. 2015, 109, 23–30. [Google Scholar] [CrossRef]
- Migahid, M.M.; Elghobashy, R.M.; Bidak, L.M.; Amin, A.W. Priming of Silybum marianum (L.) Gaertn seeds with H2O2 and magnetic field ameliorates seawater stress. Heliyon 2019, 5, e01886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paparella, S.; Araújo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT, Food and Agriculture Organization of the United Nations, Statistical Database. Available online: https://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 22 June 2022).
- Liu, J.; Feng, H.; He, J.; Chen, H.; Ding, D. The effects of nitrogen and water stresses on the nitrogen-to-protein conversion factor of winter wheat. Agric. Water Manag. 2018, 210, 217–223. [Google Scholar] [CrossRef]
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on wheat yield and quality with reduced nitrogen supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef] [Green Version]
- Ducsay, L.; Ložek, O.; Marček, M.; Varényiová, M.; Hozlár, P.; Lošák, T. Possibility of selenium biofortification of winter wheat grain. Plant Soil Environ. 2016, 62, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Ramkissoon, C.; Degryse, F.; da Silva, R.; Baird, R.; Young, S.; Bailey, E.; McLaughlin, M. Improving the efficacy of selenium fertilizers for wheat biofortification. Sci. Rep. 2019, 9, 19520. [Google Scholar] [CrossRef]
- Galinha, C.; Freitas, M.C.; Pacheco, A.M.G.; Coutinho, J.; Maçãs, B.; Almeida, A.S. Determination of selenium in bread-wheat samples grown under a Se-supplementation regime in actual field conditions. J. Radioanal. Nucl. Chem. 2012, 291, 231–235. [Google Scholar] [CrossRef]
- Galinha, C.; Pacheco, A.M.G.; Freitas, M.D.; Costa, A.R.P.; Pinheiro, N.M.B.; Maçãs, B.; Almeida, A.S.; Wolterbeek, H.T. Selenium characterisation of the Portuguese bread-wheat archival collection. Crop Pasture Sci. 2015, 66, 1111–1117. [Google Scholar] [CrossRef]
- Manojlović, M.S.; Lončarić, Z.; Cabilovski, R.R.; Popović, B.; Karalić, K.; Ivezić, V.; Ademi, A.; Singh, B.R. Biofortification of wheat cultivars with selenium. Acta Agric. Scand. B Soil Plant Sci. 2019, 69, 715–724. [Google Scholar] [CrossRef]
- Min, W.; Ali, F.; Wang, M.; Dinh, T.; Fei, Z.; Banuelos, G.; Liang, D. Understanding boosting selenium accumulation in wheat (Triticum aestivum L.) following foliar selenium application at different stages, forms, and doses. Environ. Sci. Pollut. Res. Int. 2019, 27, 717–728. [Google Scholar] [CrossRef]
- Galinha, C.; Freitas, M.C.; Pacheco, A.M.G.; Kamenik, J.; Kucera, J.; Anawar, H.M.; Coutinho, J.; Macas, B.; Almeida, A.S. Selenium determination in cereal plants and cultivation soils by radiochemical neutron activation analysis. J. Radioanal. Nucl. Chem. 2012, 294, 349–354. [Google Scholar] [CrossRef]
- de Oliveira, V.C.; Faquin, V.; Andrade, F.R.; Carneiro, J.P.; da Silva Júnior, E.C.; de Souza, K.R.D.; Pereira, J.; Guilherme, L.R.G. Physiological and physicochemical responses of potato to selenium biofortification in tropical soil. Potato Res. 2019, 62, 315–331. [Google Scholar] [CrossRef]
- Khaliq, A.; Aslam, F.; Matloob, A.; Hussain, S.; Geng, M.; Wahid, A.; ur Rehman, H. Seed priming with selenium: Consequences for emergence, seedling growth, and biochemical attributes of rice. Biol. Trace Elem. Res. 2015, 166, 236–244. [Google Scholar] [CrossRef]
- da Silva, D.F.; Cipriano, P.E.; de Souza, R.R.; Siueia Júnior, M.; da Silva, R.F.; Faquin, V.; de Souza Silva, M.L.; Guimarães Guilherme, L.R. Anatomical and physiological characteristics of Raphanus sativus L. submitted to different selenium sources and forms application. Sci. Hortic. 2020, 260, 108839. [Google Scholar] [CrossRef]
- Ahmed, M.; Fayyaz-ul, H. Response of spring wheat (Triticum aestivum L.) quality traits and yield to sowing date. PLoS ONE 2015, 10, e0126097. [Google Scholar] [CrossRef]
- Nawaz, F.; Ashraf, M.Y.; Ahmad, R.; Waraich, E.A. Selenium (Se) seed priming induced growth and biochemical changes in wheat under water deficit conditions. Biol. Trace Elem. Res. 2013, 151, 284–293. [Google Scholar] [CrossRef]
- Catálogo nacional de variedades e espécies agrícolas e hortícolas 2020. Available online: https://www.dgav.pt/wp-content/uploads/2021/02/CATALOGO-NACIONAL-DE-VARIEDADES_2020.pdf (accessed on 22 June 2022).
- Almeida, A.; Maçãs, B.; Rodrigues, V.; Torrão, M. Wheat breeding: Country perspectives. In The History of Wheat Breeding in Portugal; Bonjean, A.P., Angus, W.J., Van Ginkel, M., Eds.; Lavoisier S.A.S.: Paris, France, 2016; pp. 93–125. [Google Scholar]
- Tounekti, T.; Mahdhi, M.; Al-Faifi, Z.; Khemira, H. Priming improves germination and seed reserve utilization, growth, antioxidant responses and membrane stability at early seedling stage of Saudi sorghum varieties under drought stress. Not. Bot. Horti Agrobot. Cluj Napoca 2020, 48, 938–953. [Google Scholar] [CrossRef]
- von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Hendrickson, L.; Furbank, R.; Chow, W. A Simple alternative approach to assessing the fate of absorbed light energy using chlorophyll fluorescence. Photosynth. Res. 2004, 82, 73–81. [Google Scholar] [CrossRef]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta—Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H. K Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Meth. Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Šesták, J.; Berggren, G. Study of the kinetics of the mechanism of solid-state reactions at increasing temperatures. Thermochim. Acta 1971, 3, 1–12. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Dinis, L.T.; Oliveira, M.M.; Almeida, J.; Costa, R.; Gomes-Laranjo, J.; Peixoto, F. Antioxidant activities of chestnut nut of Castanea sativa Mill. (cultivar ‘Judia’) as function of origin ecosystem. Food Chem. 2012, 132, 1–8. [Google Scholar] [CrossRef]
- Ayuso-Mateos, J.L.; Vázquez-Barquero, J.L.; Dowrick, C.; Lehtinen, V.; Dalgard, O.S.; Casey, P.; Wilkinson, C.; Lasa, L.; Page, H.; Dunn, G.; et al. Depressive disorders in Europe: Prevalence figures from the ODIN study. Br. J. Psychiatry 2001, 179, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Barros, L.; Cabrita, L.; Boas, M.V.; Carvalho, A.M.; Ferreira, I.C.F.R. Chemical, biochemical and electrochemical assays to evaluate phytochemicals and antioxidante activity of wild plants. Food Chem. 2011, 127, 1600–1608. [Google Scholar] [CrossRef]
- Swanson, C.A.; Reamer, D.C.; Veillon, C.; King, J.C.; Levander, O.A. Quantitative and qualitative aspects of selenium utilization in pregnant and nonpregnant women: An application of stable isotope methodology. Am. J. Clin. Nutr. 1983, 38, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Costa-Silva, F.; Maia, M.; Matos, C.C.; Calçada, E.; Barros, A.I.R.N.A.; Nunes, F.M. Selenium content of Portuguese unifloral honeys. J. Food Compos. Anal. 2011, 24, 351–355. [Google Scholar] [CrossRef]
- Boldrin, P.F.; Faquin, V.; Ramos, S.J.; Guilherme, L.R.G.; Bastos, C.E.A.; Carvalho, G.S.; Costa, E.T.D.S. Selenato e selenito na produção e biofortificação agronômica com selênio em arroz. Pesqui. Agropecu. Bras. 2012, 47, 831–837. [Google Scholar] [CrossRef] [Green Version]
- Ali, W.; Zhang, H.; Junaid, M.; Mao, K.; Xu, N.; Chang, C.; Rasool, A.; Wajahat Aslam, M.; Ali, J.; Yang, Z. Insights into the mechanisms of arsenic-selenium interactions and the associated toxicity in plants, animals, and humans: A critical review. Crit. Rev. Environ. Sci. Technol. 2021, 51, 704–750. [Google Scholar] [CrossRef]
- Choudhary, S.K.; Kumar, V.; Singhal, R.K.; Bose, B.; Chauhan, J.; Alamri, S.; Siddiqui, M.H.; Javed, T.; Shabbir, R.; Rajendran, K.; et al. Seed priming with Mg(NO3)2 and ZnSO4 salts triggers the germination and growth attributes synergistically in wheat varieties. Agronomy 2021, 11, 2110. [Google Scholar] [CrossRef]
- Begum, N.; Hasanuzzaman, M.; Li, Y.; Akhtar, K.; Zhang, C.; Zhao, T. Seed germination behavior, growth, physiology and antioxidant metabolism of four contrasting cultivars under combined drought and salinity in soybean. Antioxidants 2022, 11, 498. [Google Scholar] [CrossRef]
- Nawaz, J.; Hussain, M.; Jabbar, A.; Nadeem, G.; Sajid, M.; Subtain, M.; Shabbir, I. Seed Priming A Technique. Int. J. Agric. Crop Sci. 2013, 6, 1373–1381. [Google Scholar]
- Wolny, E.; Betekhtin, A.; Rojek, M.; Braszewska-Zalewska, A.; Lusinska, J.; Hasterok, R. Germination and the early stages of seedling development in Brachypodium distachyon. Int. J. Mol. Sci. 2018, 19, 2916. [Google Scholar] [CrossRef] [Green Version]
- Komalasari, O.; Arief, R. Effect of soaking duration in hydropriming on seed vigor of sorghum (Sorghum bicolor L. moench). IOP Conf. Ser.: Earth Environ. Sci. 2020, 484, 012121. [Google Scholar] [CrossRef]
- Du, B.; Luo, H.; He, L.; Zhang, L.; Liu, Y.; Mo, Z.; Pan, S.; Tian, H.; Duan, M.; Tang, X. Rice seed priming with sodium selenate: Effects on germination, seedling growth, and biochemical attributes. Sci. Rep. 2019, 9, 4311. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.; Maroco, J.; Pereira, J. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Luo, H.W.; He, L.X.; Du, B.; Wang, Z.M.; Zheng, A.X.; Lai, R.F.; Tang, X.R. Foliar application of selenium (Se) at heading stage induces regulation of photosynthesis, yield formation, and quality characteristics in fragrant rice. Photosynthetica 2019, 57, 1007–1014. [Google Scholar] [CrossRef] [Green Version]
- Guidi, L.; Lo Piccolo, E.; Landi, M. Chlorophyll Fluorescence, photoinhibition and abiotic stress: Does it make any difference the fact to be a C3 or C4 species? Front Plant Sci. 2019, 10, 174. [Google Scholar] [CrossRef]
- Haghighi, M.; Sheibanirad, A.; Pessarakli, M. Effects of selenium as a beneficial element on growth and photosynthetic attributes of greenhouse cucumber. J. Plant Nutr. 2016, 39, 1493–1498. [Google Scholar] [CrossRef] [Green Version]
- Mazzafera, P. Growth and biochemical alterations in coffee due to selenite toxicity. Plant Soil 1998, 201, 189–196. [Google Scholar] [CrossRef]
- Jain, M.; Gadre, R. Inhibition of chlorophyll synthesis and enzymes of nitrogen assimilation by selenite in excised maize leaf sefments during greening. Water Air Soil Pollut. 1998, 104, 161–166. [Google Scholar] [CrossRef]
- Padmaja, K.; Prasad, D.D.; Prasad, A.R. Selenium as a novel regulator of porphyrin biosynthesis in germinating seedlings of mung bean (Phaseolus vulgaris). Biochem. Int. 1990, 22, 441–446. [Google Scholar]
- Yao, X.; Chu, J.; Wang, G. Effects of Selenium on Wheat Seedlings Under Drought Stress. Biol. Trace Elem. Res. 2009, 130, 283–290. [Google Scholar] [CrossRef]
- Nawaz, F.; Naeem, M.; Ashraf, M.Y.; Tahir, M.N.; Zulficiar, B.; Salahuddin, M.; Shabbir, R.N.; Aslam, M. Selenium supplementation affects physiological and biochemical processes to improve fodder yield and quality of maize (Zea mays L.) under water deficit conditions. Front. Plant Sci. 2016, 7, 1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, C.; Rodrigues, M.Â.; Pinto, L.; Gonçalves, A.; Silva, E.; Martins, S.; Rocha, L.; Pavia, I.; Arrobas, M.; Ribeiro, A.C.; et al. Grey and Black Anti-Hail Nets Ameliorated Apple (Malus × domestica Borkh. cv. Golden Delicious) Physiology under Mediterranean Climate. Plants 2021, 10, 2578. [Google Scholar] [CrossRef] [PubMed]
- Taha, R.S.; Seleiman, M.F.; Shami, A.; Alhammad, B.A.; Mahdi, A.H.A. Integrated Application of Selenium and Silicon Enhances Growth and Anatomical Structure, Antioxidant Defense System and Yield of Wheat Grown in Salt-Stressed Soil. Plants 2021, 10, 1040. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, A.; Baker, D.A. The effect of water stress on the activities of key regulatory enzymes of the sucrose to starch pathway in wheat. Plant Growth Regul. 2001, 35, 81–91. [Google Scholar] [CrossRef]
- Saffaryazdi, A.; Lahouti, M.; Ganjeali, A.; Bayat, H. Impact of selenium supplementation on growth and selenium accumulation on spinach (Spinacia oleracea L.) plants. Not. Sci. Biol. 2012, 4, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Ríos, J.J.; Blasco, B.; Cervilla, L.M.; Rosales, M.A.; Sanchez-Rodriguez, E.; Romero, L.; Ruiz, J.M. Production and detoxification of H2O2 in lettuce plants exposed to selenium. Ann. Appl. Biol. 2009, 154, 107–116. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stem Length (mm) | Spike Length (mm) | Tillers Number (nº) | Grain Number (nº) | Thousand Grain (g) | Grain Biomass (mg Plant−1) | Straw Biomass (mg Plant−1) | Above-Ground Biomass (mg Plant−1) | Harvest Index (%) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Watering Regime (W) | ||||||||||
| WW | 583 ± 6.4 a | 83 ± 1.05 | 4.1 ± 0.2 a | 124 ± 4.3 a | 32 ± 0.43 | 3909 ± 125 a | 3273 ± 49 a | 7182 ± 146 a | 54 ± 0.82 a | |
| WD | 541 ± 6.5 b | 85 ± 1.20 | 3.3 ± 0.18 b | 66 ± 2.4 b | 32 ± 0.46 | 2108 ± 80 b | 3122 ± 80 b | 5231 ± 103 b | 40 ± 1.13 b | |
| Soaking (S) | ||||||||||
| Untreated | 544 ± 10.6 b | 79 ± 1.80 b | 4.0 ± 0.26 | 86 ± 5.1 | 33 ± 0.71 a | 2809 ± 174 b | 2834 ± 52 c | 5642 ± 196 c | 49 ± 1.62 a | |
| Water | 559 ± 7.7 ab | 87 ± 1.17 a | 3.5 ± 0.24 | 100 ± 7.5 | 33 ± 0.52 a | 3266 ± 231 a | 3032 ± 72 b | 6298 ± 287 b | 50 ± 1.39 a | |
| 2.5 mM Se | 589 ± 9.0 a | 90 ± 1.27 a | 3.9 ± 0.27 | 96 ± 6.9 | 31 ± 0.59 ab | 2988 ± 211 ab | 3933 ± 72 a | 6921 ± 201 a | 41 ± 1.93 b | |
| 25 mM Se | 551 ± 9.9 b | 80 ± 1.43 b | 3.5 ± 0.32 | 98 ± 7.5 | 31 ± 0.61 b | 2939 ± 207 b | 2923 ± 41 c | 5862 ± 213 c | 49 ± 1.83 a | |
| W × S | ||||||||||
| WW | Untreated | 560 ± 13.3 | 82 ± 2.02 cd | 3.7 ± 0.18 ab | 106 ± 4.7 | 33 ± 0.99 | 3488 ± 174 | 2897 ± 83 c | 6385 ± 214 b | 55 ± 1.26 ab |
| Water | 580 ± 10.1 | 86 ± 1.91 bc | 4.2 ± 0.38 a | 134 ± 10.9 | 33 ± 0.72 | 4353 ± 317 | 3457 ± 42 b | 7810 ± 334 a | 55 ± 1.87 ab | |
| 2.5 mM Se | 612 ± 11.5 | 87 ± 1.81 b | 4.3 ± 0.37 a | 131 ± 7.0 | 31 ± 0.80 | 4032 ± 218 | 3765 ± 28 b | 7797 ± 220 a | 51 ± 1.40 b | |
| 25 mM Se | 576 ± 13.2 | 77 ± 2.03 de | 4.3 ± 0.54 a | 127 ± 10.1 | 30 ± 0.78 | 3788 ± 257 | 2962 ± 41 c | 6750 ± 260 b | 57 ± 1.80 a | |
| WD | Untreated | 527 ± 16.1 | 76 ± 2.97 e | 4.4 ± 0.50 a | 63 ± 4.9 | 33 ± 1.07 | 2045 ± 173 | 2762 ± 57 c | 4807 ± 181 c | 41 ± 1.95 c |
| Water | 541 ± 10.0 | 88 ± 1.40 ab | 3.0 ± 0.26 | 71 ± 3.8 | 33 ± 0.76 | 2343 ± 133 | 2670 ± 46 c | 5013 ± 138 c | 46 ± 1.55 c | |
| 2.5 mM Se | 567 ± 12.2 | 93 ± 1.61 a | 3.5 ± 0.38 ab | 61 ± 3.9 | 32 ± 0.89 | 1944 ± 144 | 4100 ± 133 a | 6044 ± 189 b | 32 ± 1.92 d | |
| 25 mM Se | 526 ± 12.9 | 82 ± 1.93 cd | 2.8 ± 0.24 b | 68 ± 6.2 | 31 ± 0.95 | 2089 ± 183 | 2885 ± 70 c | 4974 ± 190 c | 42 ± 2.19 c | |
| Two-way ANOVA (p-values) | ||||||||||
| W | <0.0001 | 0.2229 | 0.0073 | <0.0001 | 0.5636 | <0.0001 | 0.0013 | <0.0001 | <0.0001 | |
| S | 0.0018 | <0.0001 | 0.5026 | 0.0860 | 0.0182 | 0.0492 | <0.0001 | <0.0001 | <0.0001 | |
| W × S | 0.9190 | 0.0220 | 0.0196 | 0.2981 | 0.9317 | 0.3945 | <0.0001 | 0.0299 | 0.0344 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, L.; Silva, E.; Pavia, I.; Ferreira, H.; Matos, C.; Osca, J.M.; Moutinho-Pereira, J.; Lima-Brito, J. Seed Soaking with Sodium Selenate as a Biofortification Approach in Bread Wheat: Effects on Germination, Seedling Emergence, Biomass and Responses to Water Deficit. Agronomy 2022, 12, 1975. https://doi.org/10.3390/agronomy12081975
Rocha L, Silva E, Pavia I, Ferreira H, Matos C, Osca JM, Moutinho-Pereira J, Lima-Brito J. Seed Soaking with Sodium Selenate as a Biofortification Approach in Bread Wheat: Effects on Germination, Seedling Emergence, Biomass and Responses to Water Deficit. Agronomy. 2022; 12(8):1975. https://doi.org/10.3390/agronomy12081975
Chicago/Turabian StyleRocha, Luís, Ermelinda Silva, Ivo Pavia, Helena Ferreira, Carlos Matos, José M. Osca, José Moutinho-Pereira, and José Lima-Brito. 2022. "Seed Soaking with Sodium Selenate as a Biofortification Approach in Bread Wheat: Effects on Germination, Seedling Emergence, Biomass and Responses to Water Deficit" Agronomy 12, no. 8: 1975. https://doi.org/10.3390/agronomy12081975