
In Vitro Regeneration of Stevia (Stevia rebaudiana Bertoni) and Evaluation of the Impacts of Growth Media Nutrients on the Biosynthesis of Steviol Glycosides (SGs)

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sterilization
2.2. Media Preparation
2.3. Surface Sterilization of Explants, Culture Establishment and Maintenance
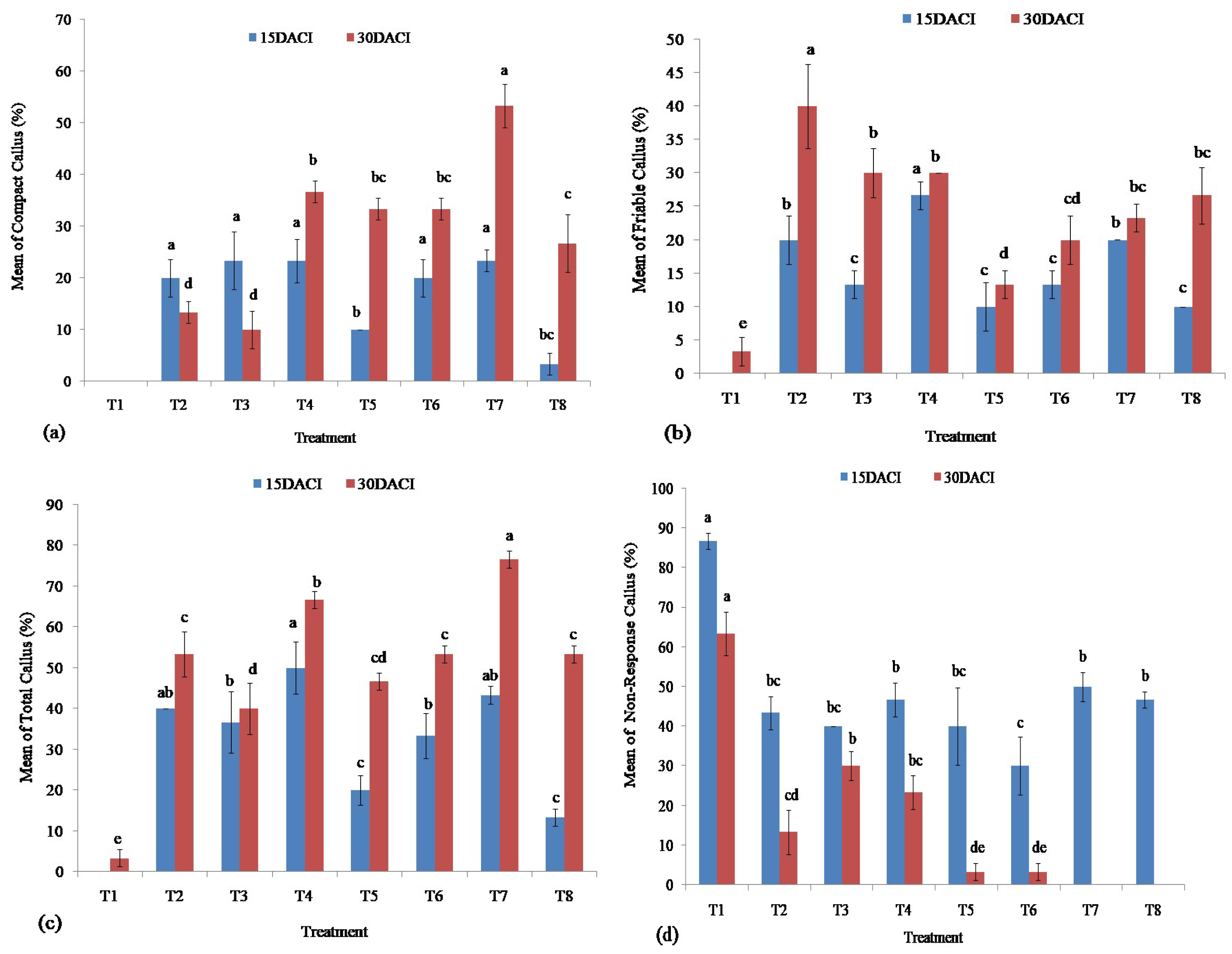
2.4. Callus Induction and Characterization
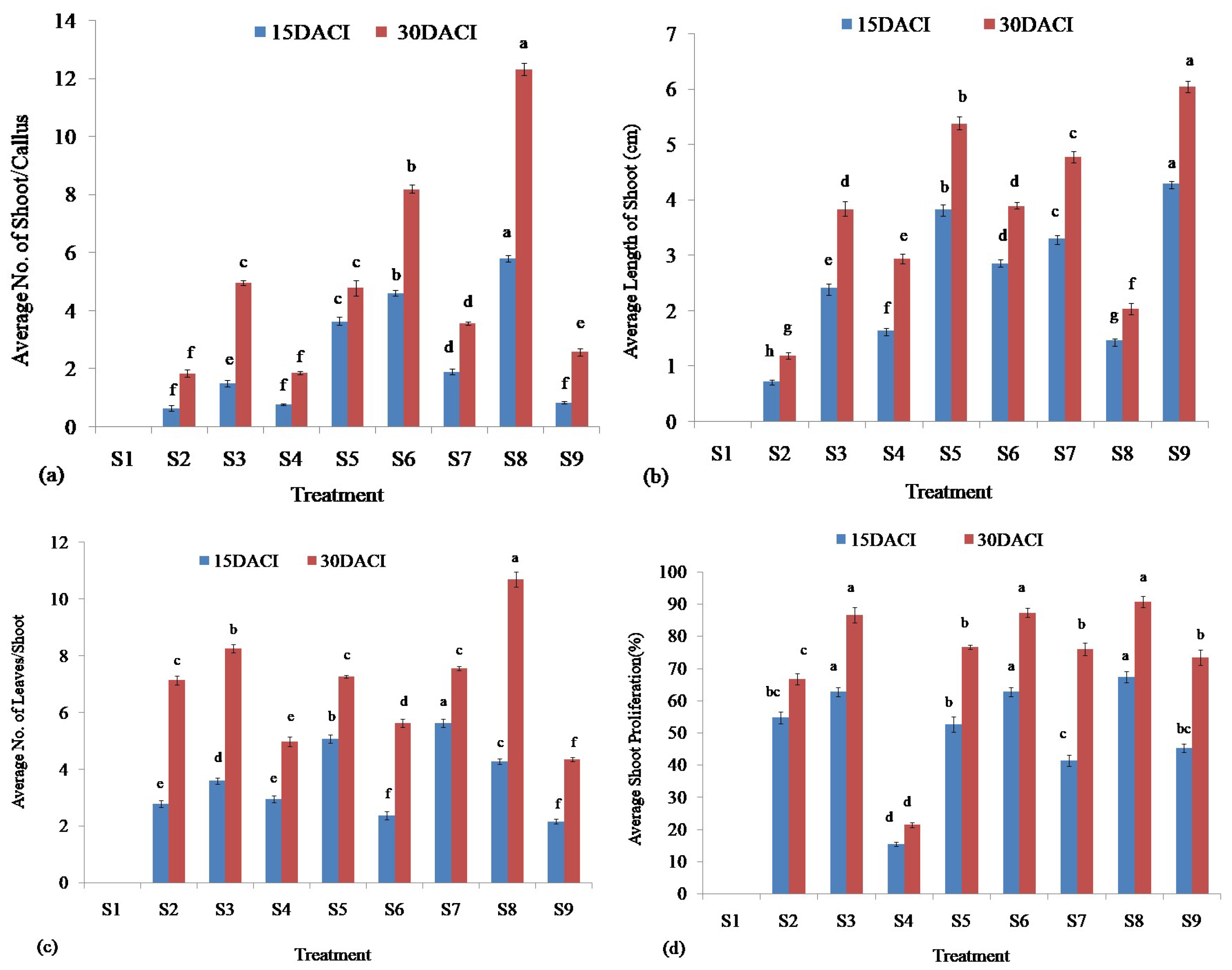
2.5. Shoot Initiation
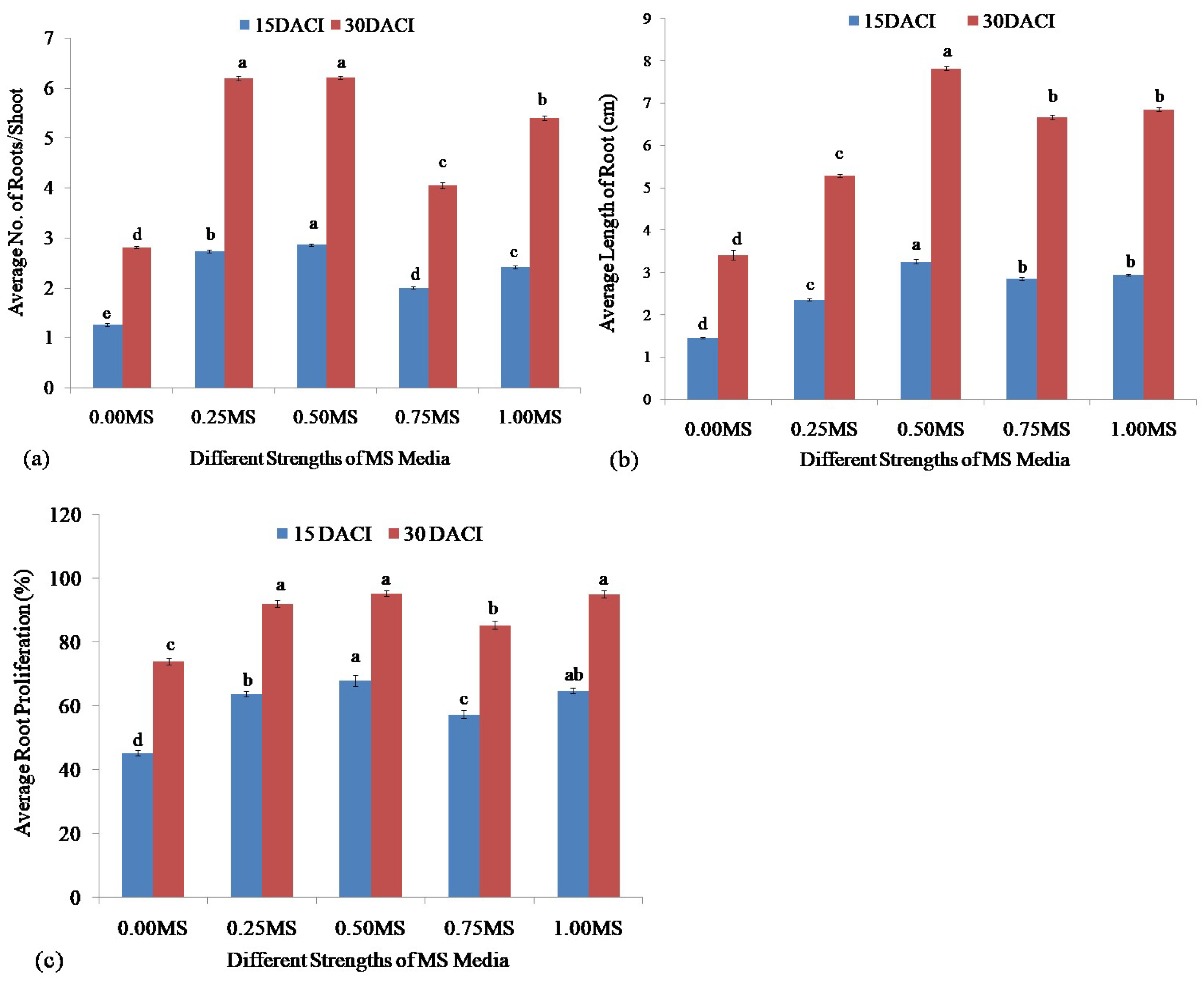
2.6. Root Induction
2.7. Steviol Glycosides (SGs) Extraction
2.8. HPLC Analysis
2.9. Extraction of Total RNA
2.10. Quantitative Reverse Transcription PCR (RT q-PCR) Analysis for UGT76G1, UGT74G1, and UGT85C2
2.11. Statistical Analysis
3. Results
3.1. Surface Sterilization of Explants
3.2. Callus Induction
3.3. Shoot Regeneration
3.4. Root Induction
3.5. HPLC Analysis for Steviol Glycosides (SGs)
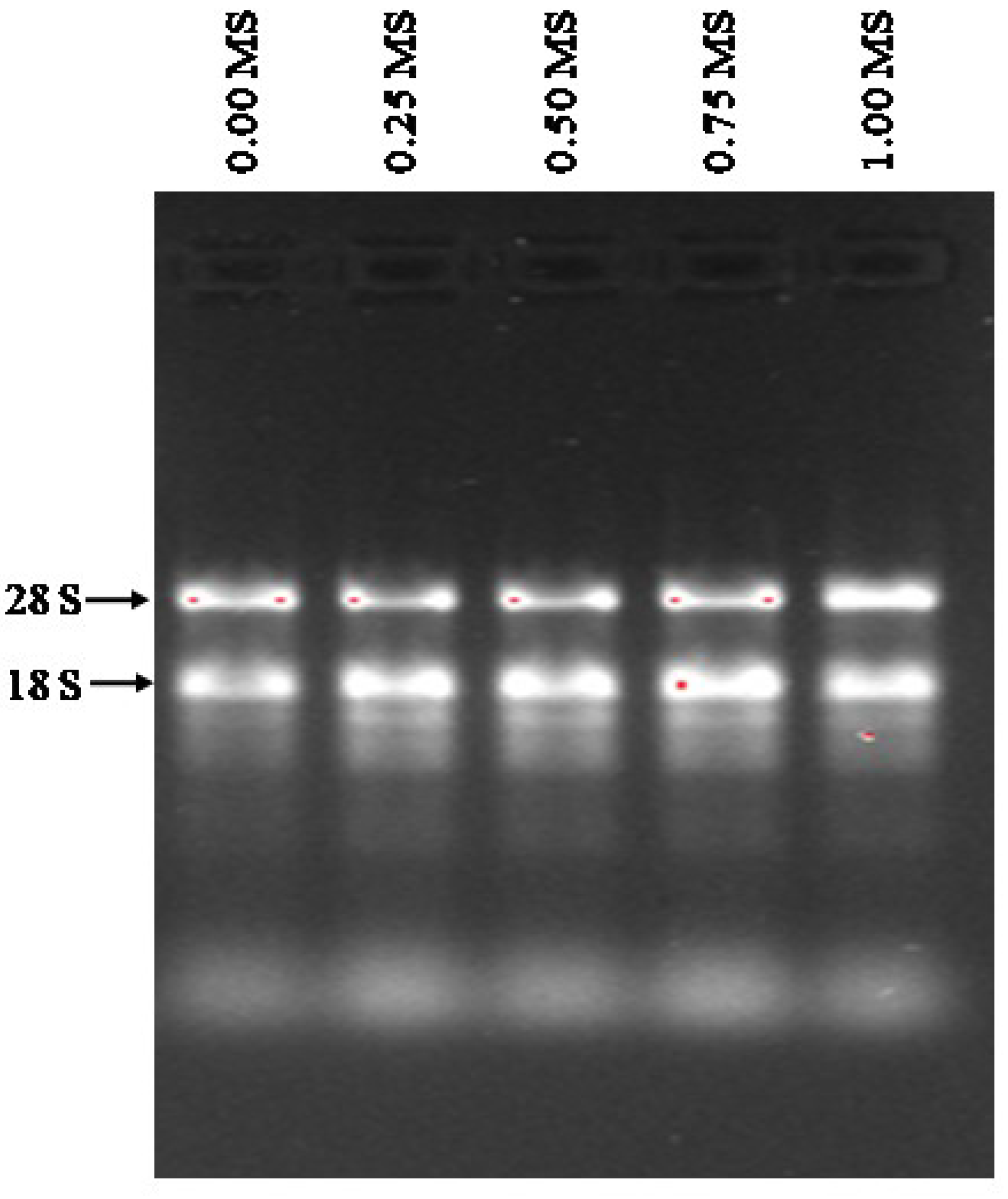
3.6. Quality and Concentrations of RNAs Isolated from Stevia Leaves
3.7. RT q-PCR Analysis for UGT76G1, UGT74G1 and UGT85C2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yadav, A.K.; Singh, S.; Dhyani, D.; Ahuja, P.S. A Review on the Improvement of Stevia [Stevia rebaudiana (Bertoni)]. Can. J. Plant Sci. 2011, 91, 1–27. [Google Scholar] [CrossRef]
- Lemus-Mondaca, R.; Vega-Gálvez, A.; Zura-Bravo, L.; Kong, A.H. Stevia rebaudiana Bertoni, Source of a High-Potency Natural Sweetener: A Comprehensive Review on the Biochemical, Nutritional and Functional Aspects. Food Chem. 2012, 132, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Shivanna, N.; Naika, M.; Khanum, F.; Kaul, V.K. Antioxidant, Anti-Diabetic and Renal Protective Properties of Stevia rebaudiana. J. Diabetes Complicat. 2013, 27, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Singh, G.; Singh, P.; Parmar, R.; Paul, N.; Vashist, R.; Swarnkar, M.K.; Kumar, A.; Singh, S.; Singh, A.K.; et al. Molecular Dissection of Transcriptional Reprogramming of Steviol Glycosides Synthesis in Leaf Tissue during Developmental Phase Transitions in Stevia rebaudiana Bert. Sci. Rep. 2017, 7, 11835. [Google Scholar] [CrossRef]
- Noranida, W.; Mohd, W.; Ibrahim, N. The Growth and Yield of Stevia (Stevia rebaudiana Bertoni) Grown on Organically Amended Sandy Medium. Int. J. Sci. Adv. Technol. 2015, 5, 14–16. [Google Scholar]
- Zhang, S.-S.; Chen, H.; Xiao, J.-Y.; Liu, Q.; Xiao, R.-F.; Wu, W. Mutations in the Uridine Diphosphate Glucosyltransferase 76G1 Gene Result in Different Contents of the Major Steviol Glycosides in Stevia rebaudiana. Phytochemistry 2019, 162, 141–147. [Google Scholar] [CrossRef]
- Ceunen, S.; Geuns, J.M.C. Steviol Glycosides: Chemical Diversity, Metabolism, and Function. J. Nat. Prod. 2013, 76, 1201–1228. [Google Scholar] [CrossRef]
- Lemus-Mondaca, R.; Vega-Gálvez, A.; Rojas, P.; Stucken, K.; Delporte, C.; Valenzuela-Barra, G.; Jagus, R.J.; Agüero, M.V.; Pasten, A. Antioxidant, Antimicrobial and Anti-Inflammatory Potential of Stevia rebaudiana Leaves: Effect of Different Drying Methods. J. Appl. Res. Med. Aromat. Plants 2018, 11, 37–46. [Google Scholar] [CrossRef]
- Panagiotou, C.; Mihailidou, C.; Brauhli, G.; Katsarou, O.; Moutsatsou, P. Effect of Steviol, Steviol Glycosides and Stevia Extract on Glucocorticoid Receptor Signaling in Normal and Cancer Blood Cells. Mol. Cell. Endocrinol. 2018, 460, 189–199. [Google Scholar] [CrossRef]
- Hsu, Y.H.; Liu, J.C.; Kao, P.F.; Lee, C.N.; Chen, Y.J.; Hsieh, M.H.; Chan, P. Antihypertensive Effect of Stevioside in Different Strains of Hypertensive Rats. Chin. Med. J. 2002, 65, 1–6. [Google Scholar]
- Brijesh, K.; Kamath, M. Experimental Evaluation of Anti-Hyperglycemic and Hypolipidemic Effects of Stevia rebaudiana, Anacardium Occidentale on Wistar Rats. Int. J. Basic Clin. Pharmacol. 2016, 5, 2463–2467. [Google Scholar] [CrossRef]
- Chatsudthipong, V.; Muanprasat, C. Stevioside and Related Compounds: Therapeutic Benefits beyond Sweetness. Pharmacol. Ther. 2009, 121, 41–54. [Google Scholar] [CrossRef]
- Casas-Grajales, S.; Ramos-Tovar, E.; Chávez-Estrada, E.; Alvarez-Suarez, D.; Hernández-Aquino, E.; Reyes-Gordillo, K.; Cerda-García-Rojas, C.M.; Camacho, J.; Tsutsumi, V.; Lakshman, M.R.; et al. Antioxidant and Immunomodulatory Activity Induced by Stevioside in Liver Damage: In Vivo, in vitro and in Silico Assays. Life Sci. 2019, 224, 187–196. [Google Scholar] [CrossRef]
- Zaidan, U.H.; Mohamad Zen, N.I.; Amran, N.A.; Shamsi, S.; Gani, S.S.A. Biochemical Evaluation of Phenolic Compounds and Steviol Glycoside from Stevia rebaudiana Extracts Associated with in vitro Antidiabetic Potential. Biocatal. Agric. Biotechnol. 2019, 18, 101049. [Google Scholar] [CrossRef]
- Kurek, J.M.; Krejpcio, Z. The Functional and Health-Promoting Properties of Stevia rebaudiana Bertoni and Its Glycosides with Special Focus on the Antidiabetic Potential—A Review. J. Funct. Foods 2019, 61, 103465. [Google Scholar] [CrossRef]
- Alavala, S.; Sangaraju, R.; Nalban, N.; Sahu, B.D.; Jerald, M.K.; Kilari, E.K.; Sistla, R. Stevioside, a Diterpenoid Glycoside, Shows Anti-Inflammatory Property against Dextran Sulphate Sodium-Induced Ulcerative Colitis in Mice. Eur. J. Pharmacol. 2019, 855, 192–201. [Google Scholar] [CrossRef]
- Ahmad, J.; Khan, I.; Johnson, S.K.; Alam, I.; Din, Z.U. Effect of Incorporating Stevia and Moringa in Cookies on Postprandial Glycemia, Appetite, Palatability, and Gastrointestinal Well-Being. J. Am. Coll. Nutr. 2018, 37, 133–139. [Google Scholar] [CrossRef]
- Kazmi, A.; Khan, M.A.; Mohammad, S.; Ali, A.; Kamil, A.; Arif, M.; Ali, H. Elicitation Directed Growth and Production of Steviol Glycosides in the Adventitious Roots of Stevia rebaudiana Bertoni. Ind. Crops Prod. 2019, 139, 111530. [Google Scholar] [CrossRef]
- Samsulrizal, N.H.; Zainuddin, Z.; Noh, A.L.; Sundram, T.C. A Review of Approaches in Steviol Glycosides Synthesis. Int. J. Life Sci. Biotechnol. 2019, 2, 145–157. [Google Scholar] [CrossRef]
- Gregersen, S.; Jeppesen, P.B.; Holst, J.J.; Hermansen, K. Antihyperglycemic Effects of Stevioside in Type 2 Diabetic Subjects. Metabolism 2004, 53, 73–76. [Google Scholar] [CrossRef]
- Chen, J.; Jeppesen, P.B.; Abudula, R.; Dyrskog, S.E.U.; Colombo, M.; Hermansen, K. Stevioside Does Not Cause Increased Basal Insulin Secretion or β-Cell Desensitization as Does the Sulphonylurea, Glibenclamide: Studies In Vitro. Life Sci. 2006, 78, 1748–1753. [Google Scholar] [CrossRef]
- Jang, M.H.; Piao, X.L.; Kim, J.M.; Kwon, S.W.; Park, J.H. Inhibition of Cholinesterase and Amyloid-&bgr; Aggregation by Resveratrol Oligomers from Vitis amurensis. Phyther. Res. 2008, 22, 544–549. [Google Scholar] [CrossRef]
- Yadav, S.K.; Guleria, P. Steviol Glycosides from Stevia: Biosynthesis Pathway Review and Their Application in Foods and Medicine. Crit. Rev. Food Sci. Nutr. 2012, 52, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, T.V.; Richman, A.S.; Menassa, R.; Brandle, J.E. Spatial Organisation of Four Enzymes from Stevia rebaudiana That are Involved in Steviol Glycoside Synthesis. Plant Mol. Biol. 2006, 61, 47–62. [Google Scholar] [CrossRef]
- Brandle, J.E.; Telmer, P.G. Steviol Glycoside Biosynthesis. Phytochemistry 2007, 68, 1855–1863. [Google Scholar] [CrossRef]
- Richman, A.; Swanson, A.; Humphrey, T.; Chapman, R.; McGarvey, B.; Pocs, R.; Brandle, J. Functional Genomics Uncovers Three Glucosyltransferases Involved in the Synthesis of the Major Sweet Glucosides of Stevia rebaudiana. Plant J. 2005, 41, 56–67. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Huang, S.-Z.; Han, Y.-L.; Yuan, H.-Y.; Gu, C.-S.; Zhao, Y.-H. Base Substitution Mutations in Uridinediphosphate-Dependent Glycosyltransferase 76G1 Gene of Stevia rebaudiana Causes the Low Levels of Rebaudioside A: Mutations in UGT76G1, A Key Gene of Steviol Glycosides Synthesis. Plant Physiol. Biochem. 2014, 80, 220–225. [Google Scholar] [CrossRef]
- Kazmi, A.; Khan, M.A.; Ali, H. Biotechnological Approaches for Production of Bioactive Secondary Metabolites in Nigella Sativa: An up-to-Date Review. Int. J. Second. Metab. 2019, 6, 172–195. [Google Scholar] [CrossRef]
- Khalil, S.A.; Zamir, R.; Ahmad, N. Selection of Suitable Propagation Method for Consistent Plantlets Production in Stevia rebaudiana (Bertoni). Saudi J. Biol. Sci. 2014, 21, 566–573. [Google Scholar] [CrossRef]
- Pande, S.S.; Gupta, P. Plant Tissue Culture of Stevia rebaudiana (Bertoni): A Review. J. Pharmacogn. Phytother. 2013, 5, 26–33. [Google Scholar] [CrossRef]
- Pratibha, G.; Satyawati, S.; Sanjay, S. Micropropagation of Stevia rebaudiana (Natural Sweetener) Using Kinetin for Steviol Glycoside Production. Res. J. Biotechnol. 2010, 5, 63–67. [Google Scholar]
- Golkar, P.; Moradi, M.; Garousi, G.A. Elicitation of Stevia Glycosides Using Salicylic Acid and Silver Nanoparticles Under Callus Culture. Sugar Tech 2019, 21, 569–577. [Google Scholar] [CrossRef]
- Idrees, M.; Sania, B.; Hafsa, B.; Kumari, S.; Khan, H.; Fazal, H.; Ahmad, I.; Akbar, F.; Ahmad, N.; Ali, S.; et al. Spectral Lights Trigger Biomass Accumulation and Production of Antioxidant Secondary Metabolites in Adventitious Root Cultures of Stevia rebaudiana (Bert.). Comptes Rendus Biol. 2018, 341, 334–342. [Google Scholar] [CrossRef]
- Lucho, S.R.; do Amaral, M.N.; Benitez, L.C.; Milech, C.; Kleinowski, A.M.; Bianchi, V.J.; Braga, E.J.B. Validation of Reference Genes for RT-qPCR Studies in Stevia rebaudiana in Response to Elicitor Agents. Physiol. Mol. Biol. Plant 2018, 24, 767–779. [Google Scholar] [CrossRef]
- Gupta, P.; Sharma, S.; Saxena, S. Effect of Salts (NaCl and Na2CO3) on Callus and Suspension Culture of Stevia rebaudiana for Steviol Glycoside Production. Appl. Biochem. Biotechnol. 2014, 172, 2894–2906. [Google Scholar] [CrossRef]
- Gupta, P.; Sharma, S.; Saxena, S. Biomass Yield and Steviol Glycoside Production in Callus and Suspension Culture of Stevia rebaudiana Treated with Proline and Polyethylene Glycol. Appl. Biochem. Biotechnol. 2015, 176, 863–874. [Google Scholar] [CrossRef]
- Kazmi, A.; Khan, M.A.; Mohammad, S.; Ali, A.; Ali, H. Biotechnological Production of Natural Calorie Free Steviol Glycosides in Stevia rebaudiana: An Update on Current Scenario. Curr. Biotechnol. 2019, 8, 70–84. [Google Scholar] [CrossRef]
- Khan, T.; Abbasi, B.H.; Khan, M.A.; Azeem, M. Production of Biomass and Useful Compounds through Elicitation in Adventitious Root Cultures of Fagonia indica. Ind. Crops Prod. 2017, 108, 451–457. [Google Scholar] [CrossRef]
- Ali, A.; Mohammad, S.; Khan, M.A.; Raja, N.I.; Arif, M.; Kamil, A.; Mashwani, Z.u.R. Silver Nanoparticles Elicited In Vitro Callus Cultures for Accumulation of Biomass and Secondary Metabolites in Caralluma tuberculata. Artif. Cells Nanomed. Biotechnol. 2019, 47, 715–724. [Google Scholar] [CrossRef]
- Ho, T.-T.; Lee, J.-D.; Jeong, C.-S.; Paek, K.-Y.; Park, S.-Y. Improvement of Biosynthesis and Accumulation of Bioactive Compounds by Elicitation in Adventitious Root Cultures of Polygonum multiflorum. Appl. Microbiol. Biotechnol. 2018, 102, 199–209. [Google Scholar] [CrossRef]
- Behroozi, P.; Baghizadeh, A.; Saei, A.; Kharazmi, S. Quantitative Analysis of Uridine Diphosphate Glycosyltransferase UGT85C2, UGT74G1 and UGT76G1 Genes Expression in Stevia rebaudiana under Different Irrigations. Russ. J. Plant Physiol. 2017, 64, 67–72. [Google Scholar] [CrossRef]
- Kahrizi, D.; Ghari, S.M.; Ghaheri, M.; Fallah, F.; Ghorbani, T.; Beheshti Ale Agha, A.; Kazemi, E.; Ansarypour, Z. Effect of KH2PO4 on Gene Expression, Morphological and Biochemical Characteristics of Stevia rebaudiana Bertoni under In Vitro Conditions. Cell. Mol. Biol. 2017, 63, 107–111. [Google Scholar] [CrossRef]
- Blinstrubien, A.; Burbulis, N.; Juškeviciute, N.; Vaitkeviciene, N.; Žukiene, R. Effect of Growth Regulators on Stevia rebaudiana Bertoni Callus Genesis and Influence of Auxin and Proline to Steviol Glycosides, Phenols, Flavonoids Accumulation, and Antioxidant Activity In Vitro. Molecules 2020, 25, 2759. [Google Scholar] [CrossRef]
- Depuydt, S.; Hardtke, C.S. Hormone Signalling Crosstalk in Plant Growth Regulation. Curr. Biol. 2011, 21, R365–R373. [Google Scholar] [CrossRef]
- Nakano, T.; Tanaka, S.; Ohtani, M.; Yamagami, A.; Takeno, S.; Hara, N.; Mori, A.; Nakano, A.; Hirose, S.; Himuro, Y.; et al. FPX Is a Novel Chemical Inducer That Promotes Callus Formation and Shoot Regeneration in Plants. Plant Cell Physiol. 2018, 59, 1555–1567. [Google Scholar] [CrossRef]
- Kahrizi, D.; Ghaheri, M.; Yari, Z.; Yari, K.; Bahraminejad, S. Investigation of different concentrations of MS media effects on gene expression and steviol glycosides accumulation in Stevia rebaudiana Bertoni. Cell. Mol. Biol. 2018, 51, 23–27. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant 1962, 15, 474–497. [Google Scholar] [CrossRef]
- Bergs, D.; Burghoff, B.; Joehnck, M.; Martin, G.; Schembecker, G. Fast and Isocratic HPLC-Method for Steviol Glycosides Analysis from Stevia rebaudiana Leaves. J. Verbrauch. Leb. 2012, 7, 147–154. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Singh, V.; Tyagi, A.; Chauhan, P.K.; Kumari, P.; Kaushal, S. Identification and Prevention of Bacterial Contimination on Explant Used in Plant Tissue Culture Labs. Int. J. Pharm. Pharm. Sci. 2011, 3, 160–163. [Google Scholar]
- Halim, M.A.; Alam, M.F.; Rahman, M.H.; Hossain, M.B.; Uddin, M.B. Sterilization Process For In Vitro Regeneration Of Stevia (Stevia Rebundiana Bertoni). Int. J. Bus. Soc. Sci. Res. 2017, 4, 320–323. [Google Scholar]
- Urbi, Z.; Zainuddin, Z. Standardization of Surface Sterilization Protocol of Field Grown Stevia rebaudiana Prior to In Vitro Clonal Propagation. J. Teknol. 2015, 77, 141–146. [Google Scholar] [CrossRef]
- Hammond, R.; Buah, J.N.; Asare, P.A.; Acheampong, S. Optimizing Sterilization Condition for the Initiation of Sweet Potato (Ipomoea batatas) Culture In Vitro. Asian J. Biotechnol. 2014, 6, 25–37. [Google Scholar] [CrossRef]
- Mng’omba, S.A.; Sileshi, G.; Toit, E.S.d.; Akinnifesi, F.K. Efficacy and Utilization of Fungicides and Other Antibiotics for Aseptic Plant Cultures. Fungic. Plant Anim. Dis. 2012, 5, 245–254. [Google Scholar] [CrossRef]
- Sen, M.K.; Hena, M.A.; Jamal, M.; Nasrin, S. Sterilization Factors Affect Seed Germination and Proliferation of Achyranthes Aspera Cultured In Vitro. Environ. Exp. Biol. 2013, 2013, 119–123. [Google Scholar]
- Çölgeçen, H.; Koca, U.; Toker, G. Influence of Different Sterilization Methods on Callus Initiation and Production of Pigmented Callus in Arnebia Densif Ora Ledeb. Turk. J. Biol. 2011, 35, 513–520. [Google Scholar] [CrossRef]
- Khatun, M.M.; Tanny, T.; Razzak, A.M.; Alam, M.F.; Uddin, M.E.; Amin, R.; Yesmin, S. Standardization of In Vitro Sterilization Procedures for Micropropagation of Ginger (Zingiber Officinale Rosc.). Int. J. Appl. Biol. Pharm. Technol. 2016, 7, 131–138. [Google Scholar]
- Zand, V.; Salem-Milani, A.; Shahi, S.; Akhi, M.T.; Vazifekhah, S. Efficacy of Different Concentrations of Sodium Hypochlorite and Chlorhexidine in Disinfection of Contaminated Resilon Cones. Med. Oral Patol. Oral Cir. Bucal 2012, 17, 352–355. [Google Scholar] [CrossRef]
- Guruchandran, V.; Sasikumar, C. Organogenic Plant Regeneration via Callus Induction in Stevia rebaudiana Bert. Int. J. Curr. Microbiol. Appl. Sci. 2013, 2, 56–61. [Google Scholar]
- Singh, M.; Saharan, V.; Dayma, J.; Rajpurohit, D.; Sen, Y.; Sharma, A. In Vitro Propagation of Stevia rebaudiana (Bertoni): An Overview. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1010–1022. [Google Scholar] [CrossRef]
- Ahmad, N.; Fazal, H.; Zamir, R.; Khalil, S.A.; Abbasi, B.H. Callogenesis and Shoot Organogenesis from Flowers of Stevia rebaudiana (Bert.). Sugar Tech 2011, 13, 174–177. [Google Scholar] [CrossRef]
- Abdelmaksood, A.W.M.; Zavdetovna, K.L.; Arnoldovna, T.O. Effect of Different Plant Growth Regulators on the In Vitro Induction and Maintenance of Callus from Different Explants of Hyoscyamus muticus L. J. Appl. Environ. Biol. Sci. 2017, 7, 27–35. [Google Scholar]
- Wada, Y.; Tamura, T.; Kodama, T.; Yamaki, T.; Uchida, Y. Callus Cultures and Morphogenesis of Stevia rebaudiana Bertoni. J. Jpn. Oil Chem. Soc. 1981, 30, 215–219. [Google Scholar] [CrossRef]
- Wan, G.L.; Naeem, M.S.; Geng, X.X.; Xu, L.; Li, B.; Jilani, G.; Zhou, W.J. Optimization of Microspore Embryogenesis and Plant Regeneration Protocols for Brassica napus. Int. J. Agric. Biol. 2011, 13, 83–88. [Google Scholar]
- Huii, L.; Guoping, Z.; Guozheng, S.; Songlin, R.; Qiaojuan, F. Callus Induction and Plant Regeneration from Mature Seeds of Salvia splendens. Int. J. Agric. Biol. 2012, 14, 445–449. [Google Scholar]
- Ahmad, N.; Fazal, H.; Abbasi, B.H.; Rashid, M.; Mahmood, T.; Fatima, N. Efficient Regeneration and Antioxidant Potential in Regenerated Tissues of Piper nigrum L. Plant Cell. Tissue Organ. Cult. 2010, 102, 129–134. [Google Scholar] [CrossRef]
- Zuraida, A.R.; Nurl Shahnadz, A.H.; Harteeni, A.; Roowi, S.; Che Radziah, C.M.Z.; Sreeramanan, S. A Novel Approach for Rapid Micropropagation of Maspine Pineapple (Ananas comosus L.) Shoots Using Liquid Shake Culture System. Afr. J. Biotechnol. 2011, 10, 3859–3866. [Google Scholar] [CrossRef]
- Hamad, A.M.; Taha, R.M. Effect of Benzylaminopurine (BAP) on In Vitro Proliferation and Growth of Pineapple (Ananas Comosus L. Merr.) Cv. Smooth Cayenne. J. Appl. Sci. 2008, 8, 4180–4185. [Google Scholar] [CrossRef]
- Thiyagarajan, M.; Venkatachalam, P. Large Scale In Vitro Propagation of Stevia rebaudiana (Bert) for Commercial Application: Pharmaceutically Important and Antidiabetic Medicinal Herb. Ind. Crops Prod. 2012, 37, 111–117. [Google Scholar] [CrossRef]
- Casal, J.J. Photoreceptor Signaling Networks in Plant Responses to Shade. Annu. Rev. Plant Biol. 2013, 64, 403–427. [Google Scholar] [CrossRef]
- Kohnen, M.V.; Schmid-Siegert, E.; Trevisan, M.; Petrolati, L.A.; Sénéchal, F.; Müller-Moulé, P.; Maloof, J.; Xenarios, I.; Fankhauser, C. Neighbor Detection Induces Organ-Specific Transcriptomes, Revealing Patterns Underlying Hypocotyl-Specific Growth. Plant Cell 2016, 28, 2889–2904. [Google Scholar] [CrossRef]
- Michaud, O.; Fiorucci, A.S.; Xenarios, I.; Fankhauser, C. Local Auxin Production Underlies a Spatially Restricted Neighbor-Detection Response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 7444–7449. [Google Scholar] [CrossRef]
- Iglesias, M.J.; Sellaro, R.; Zurbriggen, M.D.; Casal, J.J. Multiple Links between Shade Avoidance and Auxin Networks. J. Exp. Bot. 2018, 69, 213–228. [Google Scholar] [CrossRef]
- Dey, A.; Kundu, S.; Bandyopadhyay, A.; Bhattacharjee, A. Efficient Micropropagation and Chlorocholine Chloride Induced Stevioside Production of Stevia rebaudiana Bertoni. Comptes Rendus Biol. 2013, 336, 17–28. [Google Scholar] [CrossRef]
- Fakhrul, R.H.; Norrizah, J.S.; Jaapar, S.S.; Noor Anilizawatima, S. The Effect of Potassium Concentrations on the Growth and Development of Stevia rebaudiana (Bertoni) and Production of Stevioside and Rebaudioside A. Am. J. Sustain. Agric. 2014, 8, 42–51. [Google Scholar]
- Nower, A.A. In Vitro Propagation and Synthetic Seeds Production: An Efficient Methods for Stevia rebaudiana Bertoni. Sugar Tech 2014, 16, 100–108. [Google Scholar] [CrossRef]
- Ramírez-Mosqueda, M.A.; Iglesias-Andreu, L.G. Direct Organogenesis of Stevia rebaudiana Bertoni Using Thin Cell Layer (TCL) Method. Sugar Tech 2016, 18, 424–428. [Google Scholar] [CrossRef]
- Petit, E.; Jacques, A.; Daydé, J.; Vallejo, V.; Berger, M. UGT76G1 Polymorphism in Stevia rebaudiana: New Variants for Steviol Glycosides Conjugation. Plant Physiol. Biochem. 2019, 135, 563–569. [Google Scholar] [CrossRef]
- Šic Žlabur, J.; Voća, S.; Dobričević, N.; Ježek, D.; Bosiljkov, T.; Brnčić, M. Stevia rebaudiana Bertoni-A Review of Nutritional and Biochemical Properties of Natural Sweetener. Agric. Conspec. Sci. 2013, 78, 25–30. [Google Scholar]
- Chranioti, C.; Chanioti, S.; Tzia, C. Comparison of Spray, Freeze and Oven Drying as a Means of Reducing Bitter Aftertaste of Steviol Glycosides (Derived from Stevia rebaudiana Bertoni Plant)-Evaluation of the Final Products. Food Chem. 2016, 190, 1151–1158. [Google Scholar] [CrossRef]
- Goyal, S.K.; Samsher; Goyal, R.K. Stevia (Stevia rebaudiana) a Bio-Sweetener: A Review. Int. J. Food Sci. Nutr. 2010, 61, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.A.; Ceunen, S.; Geuns, J.M.; Ende, W.V.D.; De Ley, M. UDP-Dependent Glycosyltransferases Involved in the Biosynthesis of Steviol Glycosides. J. Plant Physiol. 2011, 168, 1136–1141. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Kaul, K.; Bajpai-Gupta, S.; Kaul, V.K.; Kumar, S. A Comprehensive Analysis of Fifteen Genes of Steviol Glycosides Biosynthesis Pathway in Stevia rebaudiana (Bertoni). Gene 2012, 492, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Akbari, F.; Arminian, A.; Kahrizi, D.; Fazeli, A.; Ghaheri, M. Effect of Nitrogen Sources on Gene Expression of Stevia rebaudiana (Bertoni) under In VitroConditions. Cell. Mol. Biol. 2018, 64, 11–16. [Google Scholar] [CrossRef]
- Akbari, F.; Arminian, A.; Kahrizi, D.; Fazeli, A. Effect of Nitrogen Sources on Some Morphological Characteristics of In Vitro Stevia rebaudiana Bertoni. Cell. Mol. Biol. 2017, 63, 107–111. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | 2,4-D (mg/L) | Zeatin (mg/L) |
|---|---|---|
| T1 | 0.00 | 0.00 |
| T2 | 0.10 | 0.10 |
| T3 | 0.25 | 0.10 |
| T4 | 0.50 | 0.10 |
| T5 | 0.75 | 0.10 |
| T6 | 1.00 | 0.10 |
| T7 | 1.50 | 0.10 |
| T8 | 2.00 | 0.10 |
| Treatment | BAP (mg/L) | NAA (mg/L) |
|---|---|---|
| S1 | 0.00 | 0.00 |
| S2 | 1.00 | 0.00 |
| S3 | 1.00 | 1.00 |
| S4 | 2.50 | 0.00 |
| S5 | 2.50 | 1.00 |
| S6 | 5.00 | 0.00 |
| S7 | 5.00 | 1.00 |
| S8 | 10.00 | 0.00 |
| S9 | 10.00 | 1.00 |
| Name (Accession No.) | Forward (F)/ Reverse (R) | Sequence (5′–3′) | Amplicon Size (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|
| Actin (AF548026.1) | F | CTGAGAACTGAGGGCTAGGG | 187 | 70.5 |
| R | AACCCAGCCTTGACCATTCC | |||
| Aquaporin (DQ269455.1) | F | GGAGCCGCCGTAATCTACAA | 86 | 80 |
| R | GCAATCGCCGCACCAATAAA | |||
| Calmodulin (AF474074.1) | F | ATCCGCTCACCGACGATCA | 142 | 65 |
| R | TGCAGTTCAGCTTCTGTTGG | |||
| UGT76G1 (KM206772.1) | F | CTGCCAATGCCACCGTTATT | 94 | 52 |
| R | TCATAAACCGTCTGACGCAGG | |||
| UGT74G1 (AY345982.1) | F | TTCCAGTGCTTCAACGGTGG | 124 | 61 |
| R | GTGAAGACCCAACGTGCTTG | |||
| UGT85C2 (AY345978.1) | F | CGTTCGATGAGTTGGAGCCT | 181 | 61 |
| R | AGCCACTGGAAACACTCTGG |
| Step | Temperature | Duration | Cycle (s) |
|---|---|---|---|
| Initial activation | 95 °C | 2 min. | 1 |
| Denaturation | 95 °C | 05 s | 40 |
| Annealing | * 52–80 °C | 10 s | |
| Extention | 72 °C | 15 s | |
| Melt curve | 65–95 °C | 5 s for every increment of 0.5 °C | |
| 15 Days after Culture Initiation (DACI) | 30 Days after Culture Initiation (DACI) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Period of Time (Minute) | Period of Time (Minute) | ||||||||
| 5 | 10 | 15 | 20 | 5 | 10 | 15 | 20 | ||
| Concentration of Clorox (%) | 0 | 11.11 ± 5.56 abc | 16.67 ± 0.00 ab | 5.56 ± 5.56 bc | 5.56 ± 5.56 bc | 11.11 ± 5.56 bc | 16.67 ± 0.00 b | 5.56 ± 5.56 bc | 11.11 ± 5.56 bc |
| 5 | 11.11 ± 5.56 abc | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 11.11 ± 5.56 bc | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | |
| 10 | 16.67 ± 9.62 ab | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 16.67 ± 9.62 b | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | |
| 15 | 16.67 ± 9.62 ab | 5.56 ± 5.56 bc | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 16.67 ± 9.62 b | 5.56 ± 5.56 bc | 0.00 ± 0.00 c | 0.00 ± 0.00 c | |
| 20 | 0.00 ± 0.00 c | 22.23 ± 5.56 a | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 38.89 ± 5.56 a | 0.00 ± 0.00 c | 0.00 ± 0.00 c | |
| 30 | 5.56 ± 5.56 bc | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 5.56 ± 5.56 bc | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | |
| 15 Days after Culture Initiation (DACI) | 30 Days after Culture Initiation (DACI) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Period of Time (Minute) | Period of Time (Minute) | ||||||||
| 5 | 10 | 15 | 20 | 5 | 10 | 15 | 20 | ||
| Concentration of Clorox (%) | 0 | 55.56 ± 5.56 ab | 38.89 ± 5.55 bc | 55.57 ± 20.03 ab | 66.68 ± 9.63 a | 66.67 ± 9.62 a | 66.67 ± 0.00 a | 66.67 ± 9.62 a | 66.67 ± 9.62 a |
| 5 | 66.68± 0.00 a | 16.67 ± 9.62 de | 5.56 ± 5.56 e | 27.78 ± 5.56 dc | 72.22 ± 5.55 a | 38.89 ± 14.70 b | 55.56 ± 5.56 de | 27.78 ± 5.55 bc | |
| 10 | 38.89 ± 5.55 bc | 11.11 ± 5.56 de | 16.67 ± 9.62 de | 11.11 ± 5.56 de | 38.89 ± 5.56 b | 11.11 ± 5.56 cde | 22.22 ± 5.55 bcd | 11.11 ± 5.56 cde | |
| 15 | 16.67 ± 9.62 de | 5.56 ± 5.56 e | 11.11 ± 5.56 de | 38.89 ± 11.11 bc | 22.22 ± 11.11 bcd | 5.56 ± 5.56 de | 22.22 ± 5.55 bcd | 38.89 ± 11.11 b | |
| 20 | 5.56 ± 5.56 e | 27.78 ± 5.56 dc | 0.00 ± 0.00 e | 5.56 ± 5.56 e | 11.11 ± 5.56 cde | 27.78 ± 5.55 bc | 11.11 ± 5.56 cde | 11.11 ± 5.56 cde | |
| 30 | 0.00 ± 0.00 e | 5.56 ± 5.56 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 5.56 ± 5.56 de | 16.67 ± 9.62 cde | 0.00 ± 0.00 e | 0.00 ± 0.00 e | |
| 15 Days after Inoculation (DACI) | 30 Days after Inoculation (DACI) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Period of Time (Minute) | Period of Time (Minute) | ||||||||
| 5 | 10 | 15 | 20 | 5 | 10 | 15 | 20 | ||
| Concentration of Clorox (%) | 0 | 16.67 ± 0.00 cd | 27.78 ± 5.56 bc | 22.23 ± 5.56 bcd | 11.11 ± 5.56 cd | 5.56 ± 5.56 de | 0.00 ± 0.00 e | 11.11 ± 5.56 cde | 5.56 ± 5.56 de |
| 5 | 5.56 ± 5.56 d | 50.01 ± 16.67 a | 11.11 ± 11.11 cd | 22.22 ± 14.70 bcd | 0.00 ± 0.00 e | 27.78 ± 5.55 bc | 22.22 ± 11.11 bcd | 16.67 ± 9.62 bcde | |
| 10 | 16.67 ± 9.62 cd | 38.89 ± 5.55 ab | 5.56 ± 5.56 d | 5.56 ± 5.56 d | 22.22 ± 5.55 bcd | 0.00 ± 0.00 e | 16.67 ± 0.00 bcde | 5.56 ± 5.56 de | |
| 15 | 16.67 ± 0.00 cd | 22.23 ± 5.56 bcd | 11.11 ± 5.56 cd | 16.67 ± 0.00 cd | 33.33 ± 9.62 b | 27.78 ± 5.55 bc | 11.11 ± 5.56 cde | 27.78 ± 5.55 bc | |
| 20 | 27.78 ± 5.56 bc | 16.67 ± 0.00 cd | 50.01 ± 9.62 a | 11.11 ± 5.56 cd | 61.11 ± 5.56 a | 11.11 ± 5.56 cde | 72.23 ± 5.56 a | 27.78 ± 5.55 bc | |
| 30 | 5.56 ± 5.56 d | 11.11 ± 5.56 cd | 16.67 ± 9.62 cd | 11.11 ± 5.56 cd | 66.67 ± 9.62 a | 0.00 ± 0.00 e | 16.67 ± 9.62 bcde | 55.56 ± 5.56 a | |
| 15 Days after Inoculation (DACI) | 30 Days after Inoculation (DACI) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Period of Time (Minute) | Period of Time (Minute) | ||||||||
| 5 | 10 | 15 | 20 | 5 | 10 | 15 | 20 | ||
| Concentration of Clorox (%) | 0 | 16.67 ± 0.00 d | 16.67 ± 0.00 d | 16.67 ± 9.62 d | 16.67 ± 9.62 d | 16.67 ± 0.00 f | 16.67 ± 0.00 f | 16.67 ± 9.62 f | 16.67 ± 9.62 f |
| 5 | 16.67 ± 9.62 d | 33.34 ± 9.62 cd | 83.34 ± 9.62 a | 50.01 ± 9.62 bc | 16.67 ± 9.62 f | 33.33 ± 9.62 ef | 72.22 ± 5.55 abc | 55.56 ± 5.56 cd | |
| 10 | 27.78 ± 5.56 cd | 50.01 ± 9.62 bc | 77.79 ± 11.11 a | 83.35 ± 0.00 a | 22.23 ± 5.56 f | 88.89 ± 5.56 a | 61.11 ± 5.56 cd | 83.33 ± 0.00 ab | |
| 15 | 50.00 ± 0.00 bc | 66.68 ± 9.63 ab | 77.79 ± 5.56 a | 44.45 ± 11.11 bc | 27.78 ± 5.55 ef | 61.11 ± 5.56 cd | 66.67 ± 9.62 bc | 27.78 ± 5.55 ef | |
| 20 | 66.68 ± 9.63 ab | 33.34 ± 9.62 cd | 50.01 ± 9.62 bc | 83.34 ± 9.62 a | 27.78 ± 5.55 ef | 22.22 ± 5.55 f | 16.67 ± 0.00 f | 61.11 ± 5.56 cd | |
| 30 | 88.90 ± 5.55 a | 83.35 ± 5.56 a | 83.34 ± 9.62 a | 88.90 ± 5.55 a | 22.22 ± 5.55 f | 83.33 ± 9.62 ab | 83.33 ± 9.62 ab | 44.44 ± 5.56 de | |
| RNA | Before DNAse I Treatment | After DNAse I Treatment | ||
|---|---|---|---|---|
| Yield (ng/µL) | Purity(A260/A280) | Yield (ng/µL) | Purity(A260/A280) | |
| 0.00 MS | 390.8 | 1.90 | 139.5 | 2.03 |
| 0.25 MS | 395.3 | 1.91 | 138.9 | 2.02 |
| 0.50 MS | 418.4 | 1.94 | 159.0 | 2.03 |
| 0.75 MS | 311.5 | 1.89 | 151.3 | 2.03 |
| 1.00 MS | 357.5 | 1.96 | 178.3 | 2.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghose, A.K.; Abdullah, S.N.A.; Md Hatta, M.A.; Megat Wahab, P.E. In Vitro Regeneration of Stevia (Stevia rebaudiana Bertoni) and Evaluation of the Impacts of Growth Media Nutrients on the Biosynthesis of Steviol Glycosides (SGs). Agronomy 2022, 12, 1957. https://doi.org/10.3390/agronomy12081957
Ghose AK, Abdullah SNA, Md Hatta MA, Megat Wahab PE. In Vitro Regeneration of Stevia (Stevia rebaudiana Bertoni) and Evaluation of the Impacts of Growth Media Nutrients on the Biosynthesis of Steviol Glycosides (SGs). Agronomy. 2022; 12(8):1957. https://doi.org/10.3390/agronomy12081957
Chicago/Turabian StyleGhose, Asish Kumar, Siti Nor Akmar Abdullah, Muhammad Asyraf Md Hatta, and Puteri Edaroyati Megat Wahab. 2022. "In Vitro Regeneration of Stevia (Stevia rebaudiana Bertoni) and Evaluation of the Impacts of Growth Media Nutrients on the Biosynthesis of Steviol Glycosides (SGs)" Agronomy 12, no. 8: 1957. https://doi.org/10.3390/agronomy12081957