
Cadmium Accumulation in Cereal Crops and Tobacco: A Review


Abstract
:1. Introduction
2. Cd Toxicity
3. Cadmium Uptake, Translocation and Accumulation in Cereal Crops
3.1. Multi-Omics Approaches Are Powerful Tools for Elucidating the Molecular Mechanisms of Cd Accumulation
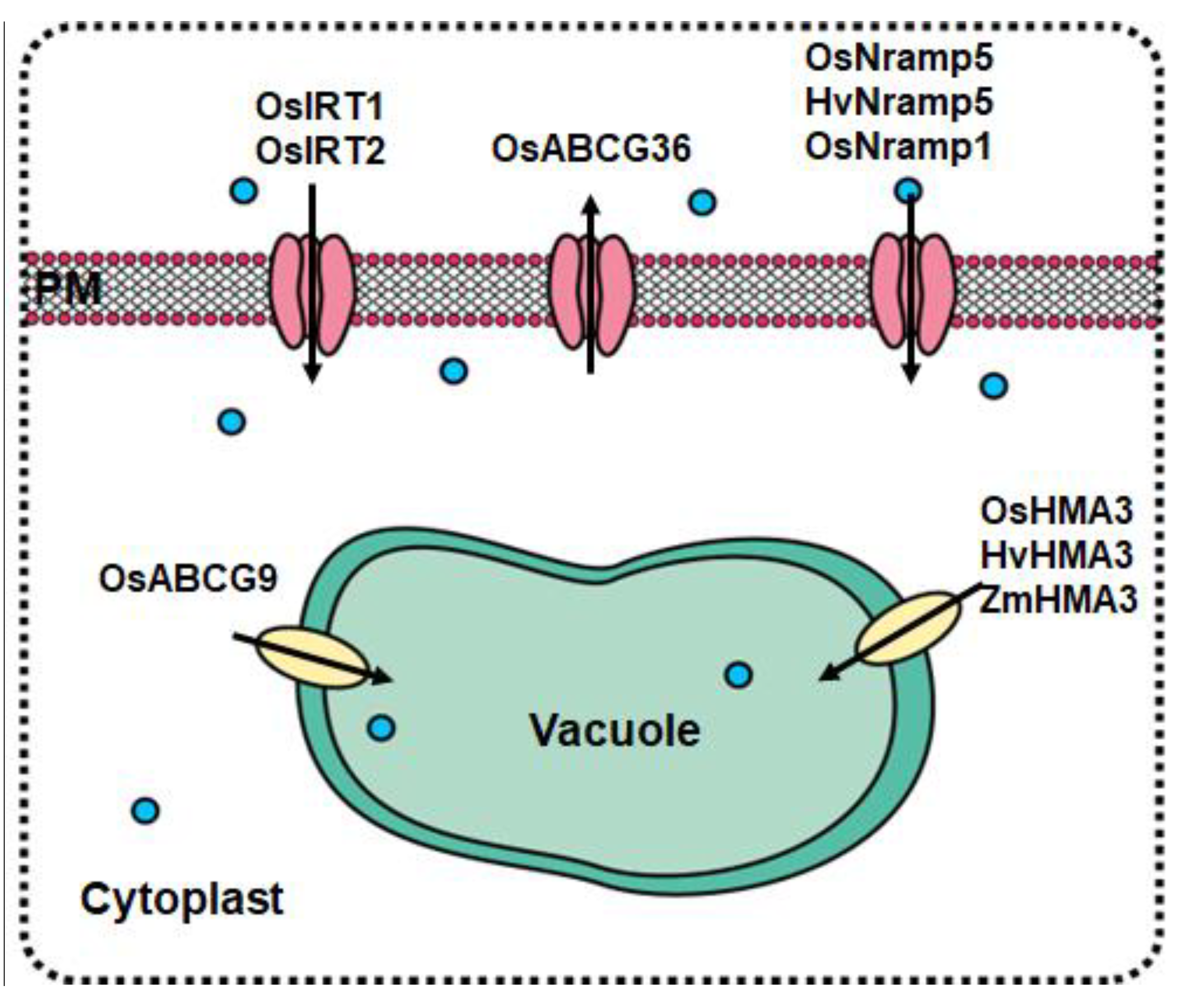
3.2. Main Transporter Families of Cd in Cereal Crops
3.3. Transcription Factors Associated with Cd Transport and Epigenetic Regulation in Cereal Crops
4. Key Genes Involved in Cd Accumulation and Tolerance in Tobacco

{kind=link}
| Category | Gene Name | Materials | Cd Treatment | Subcellular Localization | Function | Reference |
|---|---|---|---|---|---|---|
| Oryza sativa | ||||||
| Transporter | OsCd1 ↑ | Rice mutant | 1 μM/0.18 mg kg−1 | PM | Uptake of Cd | [67] |
| OsLCD ↓ | Rice mutant | 100 μM | PM | Efflux of Cd | [95] | |
| OsCCX2 ↑ | Rice mutant | 0.1 μM/3.9 mg kg−1 | PM | Cd loading into xylem | [64] | |
| OsZIP1 | Rice overexpression lines | 80 μM | PM | Efflux of Zn, Cu and Cd | [66] | |
| OsIRT1 | Rice overexpression lines | 100 μM | PM | Uptake of Fe, Zn, Mn and Cd | [96] | |
| OsIRT2 | Rice overexpression lines | 20 μM | PM | Uptake of Fe, Zn, Mn and Cd | [96] | |
| OsABCG36 | Rice mutant | 5 mM | PM | Efflux of Cd | [60] | |
| OsABCG48 | Heterologous expression in Arabidopsis | 0.5 nM | PM | Efflux of Cd | [61] | |
| OsLCT1 ↓ | Rice RNAi lines | 0.198 mg kg−1 | PM | Efflux of Cd, Ca, Mg, and Mn | [45] | |
| OsNramp1 | Rice mutant | 0.5 μM/0.88 mg kg−1 | PM | Uptake of Mn and Cd | [56] | |
| OsNramp5 ↑ | Rice mutant | 0.5 and 2.5 μM, 1.69, 2.37 and 1.53 mg kg−1 | PM | Uptake of Mn and Cd | [97] | |
| OsHMA3 ↓ | Rice overexpression lines | 1 μM | Tonoplast | Cd sequestration in root vacuoles | [39,98] | |
| OsHMA2 ↑ | Rice overexpression lines | 50 nM | PM | Cd loading to the xylem | [58] | |
| Zea mays | ||||||
| ZmHMA3 ↓ | EMS mutant | 2.24 mg kg−1 | Tonoplast | Cd sequestration in root vacuoles | [33] | |
| Hordeum vulgare | ||||||
| HvNramp5 | Barley mutant | 1 μM | PM | Uptake of Mn and Cd | [54] | |
| HvHMA3 ↓ | Rice mutant | 0.5 μM/0.63 mg kg−1 | Tonoplast | Cd sequestration in root vacuoles | [57] | |
| Triticum aestivum | ||||||
| Transcription Factor | TaWRKY70 | Heterologous expression in Arabidopsis | 20 μM | Nucleus | decreases Cd influx. | [73] |
| TdSHN1 | Transgenic tobacco | 20 μM | Nucleus | confers Cd, Cu, and Zn tolerance in yeast and transgenic tobacco plants. | [76] | |
| Zea mays | ||||||
| ZmWRKY4 | Maize RNAi lines | 400 μM | Nucleus | upregulates activities of antioxidant enzymes. | [74] | |
| Aegilops markgrafii | ||||||
| AemNAC2 ↓ | Heterologous expression in wheat | 1,050,100 μM | Nucleus | enhances Cd stress tolerance in wheat. | [78] | |
| AemNAC3 | Heterologous expression in wheat | 1,050,100 μM | Nucleus | enhances Cd stress tolerance in wheat. | [78] | |
| Epigenetic Regulator | Oryza sativa | |||||
| miRNA390 ↓ | Rice overexpression lines | 100,200 μM | displays reduced Cd tolerance and higher Cd accumulation through regulation of its target gene, OsSRK. | [99] | ||
| miRNA166 ↓ | Rice overexpression lines | 2 μM | miR166 decreases Cd accumulation and tolerance through regulation of its target gene, OsHB4. | [86] | ||
5. Approaches to Decrease Cd Accumulation
6. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Ma, J.F.; Shen, R.F.; Shao, J.F. Transport of cadmium from soil to grain in cereal crops: A review. Pedosphere 2021, 31, 3–10. [Google Scholar] [CrossRef]
- Kubier, A.; Wilkin, R.T.; Pichler, T. Cadmium in soils and groundwater: A review. Appl. Geochem. 2019, 108, 104388. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, H.P.; Kopittke, P.M.; Zhao, F.-J. Cadmium contamination in agricultural soils of China and the impact on food safety. Environ. Pollut. 2019, 249, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Sigel, A.; Sigel, H.; Sigel, R.K.O. (Eds.) Metal Ions in Life Sciences 11. Cadmium: From Toxicity to Essentiality; Springer: Cham, The Netherlands, 2013. [Google Scholar]
- Kumar, D.A.; Subrahmanyam, G.; Mondal, R.; Cabral-Pinto, M.; Shabnam, A.A.; Jigyasu, D.K.; Malyan, S.K.; Fagodiya, R.K.; Khan, S.A.; Yu, Z.-G. Bio-remediation approaches for alleviation of cadmium contamination in natural resources. Chemosphere 2021, 268, 128855. [Google Scholar] [CrossRef] [PubMed]
- Kabata-Pendias, A.; Pendias, H. Biogeochemistry of Trace Elements, Trace Elements in Soils and Plants; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Zhao, F.-J.; Ma, Y.B.; Zhu, Y.-G.; Tang, Z.; McGrath, S.P. Soil contamination in China: Current status and mitigation strategies. Environ. Sci. Technol. 2015, 49, 750–759. [Google Scholar] [CrossRef]
- Zhang, S.; Song, J.; Cheng, Y.; Liu, G.; Wallace, A.R. Trace metal(loid)s exposure through soil–tobacco–human pathway: Availability in metal-contaminated agricultural soils, transfer models and health risk assessment. Ecotoxicol. Environ. Saf. 2018, 148, 1034–1041. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, T.; Li, Y.; Wang, M.; Chen, W.; Dai, Y. Mitigating cadmium contamination of rice soils supporting tobacco–rice rotation in southern China: Win–win or lose–lose? J. Hazard. Mater. 2021, 425, 128052. [Google Scholar] [CrossRef]
- World Health Organization. Health Risks of Heavy Metals from Long-Range Transboundary Air Pollution; Regional Office for Europe: Copenhagen, Denmark, 2007. [Google Scholar]
- Zou, C.; Li, Y.; Huang, W.; Zhao, G.; Pu, G.; Su, J.; Coyne, M.S.; Chen, Y.; Wang, L.; Hu, X.; et al. Rotation and manure amendment increase soil macro-aggregates and associated carbon and nitrogen stocks in flue-cured tobacco production. Geofis. Int. 2018, 325, 49–58. [Google Scholar] [CrossRef]
- Cao, F.B.; Dai, H.X.; Hao, P.F.; Wu, F.B. Silicon regulates the expression of vacuolar H+-pyrophosphatase 1 and decreases cadmium accumulation in rice (Oryza sativa L.). Chemosphere 2020, 240, 124907. [Google Scholar] [CrossRef]
- An, M.; Hong, D.; Chang, D.; Zhang, C.; Fan, H.; Wang, K. Polymer amendment regulates cadmium migration in cadmium contaminated cotton field: Insights from genetic adaptation and phenotypic plasticity. Sci. Total Environ. 2022, 807, 151075. [Google Scholar] [CrossRef]
- Ghosh, R.; Roy, S. Cadmium toxicity in plants: Unveiling the physicochemical and molecular aspects. In Cadmium Tolerance in Plants; Academic Press: Cambridge, MA, USA, 2019; pp. 223–246. [Google Scholar]
- Lin, K.N.; Williams, D.V.; Zeng, M.; Ahmed, I.M.; Dai, H.X.; Cao, F.B.; Wu, F.B. Identification of low grain cadmium accumulation genotypes and its physiological mechanism in maize (Zea mays L.). Environ. Sci. Pollut. Res. 2022, 29, 20721–20730. [Google Scholar] [CrossRef] [PubMed]
- Quadros, I.P.S.; Madeira, N.N.; Loriato, V.A.P.; Saia, T.F.F.; Silva, J.C.; Soares, F.A.F.; Carvalho, J.R.; Reis, P.A.B.; Fontes Wellington, E.P.B.; Clarindo, C.; et al. Cadmium-mediated toxicity in plant cells is associated with the DCD/NRP-mediated cell death response. Plant Cell Environ. 2022, 45, 556–571. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.B.; Chen, F.; Sun, H.Y.; Zhang, G.P.; Chen, Z.H.; Wu, F.B. Genome-wide transcriptome and functional analysis of two contrasting genotypes reveals key genes for cadmium tolerance in barley. BMC Genom. 2014, 15, 611. [Google Scholar] [CrossRef] [PubMed]
- Faiz, S.; Shah, A.A.; Naveed, N.H.; Nijabat, A.; Yasin, N.A.; Batool, A.I.; Ali, H.M.; Javed, T.; Simon, P.W.; Ali, A. Synergistic application of silver nanoparticles and indole acetic acid alleviate cadmium induced stress and improve growth of Daucus carota L. Chemosphere 2022, 290, 133200. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Tang, L.; Peng, Y.; Shen, Z. The root iron transporter 1 governs cadmium uptake in Vicia sativa roots. J. Hazard. Mater. 2020, 398, 122873. [Google Scholar] [CrossRef]
- Baghaie, A.H.; Aghili, F.; Jafarinia, R. Soil-indigenous arbuscular mycorrhizal fungi and zeolite addition to soil synergistically increase grain yield and reduce cadmium uptake of bread wheat (through improved nitrogen and phosphorus nutrition and immobilization of Cd in roots). Environ. Sci. Pollut. Res. 2019, 26, 30794–30807. [Google Scholar] [CrossRef]
- Uraguchi, S.; Fujiwara, T. Cadmium transport and tolerance in rice: Perspectives for reducing grain cadmium accumulation. Rice 2012, 5, 5. [Google Scholar] [CrossRef]
- Chen, Q.; Wu, F.-B. Breeding for low cadmium accumulation cereals. J. Zhejiang Univ. Sci. B 2020, 21, 442–459. [Google Scholar] [CrossRef]
- Ognjanovic, B.I.; Pavlovic, S.Z.; Maletic, S.D.; Zikic, R.V.; Stajn, A.S.; Radojicic, R.M.; Saicic, Z.S.; Petrovic, V.M. Protective influence of vitamin E on antioxidant defense system in the blood of rats treated with cadmium. Physiol. Res. 2003, 52, 563–570. [Google Scholar]
- El-Demerdash, F.M.; Yousef, M.I.; Kedwany, F.S.; Baghdadi, H.H. Cadmium-induced changes in lipid peroxidation, blood hematology, biochemical parameters and semen quality of male rats: Protective role of vitamin E and β-carotene. Food Chem. Toxicol. 2004, 42, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The effects of cadmium toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.N.; Huang, Y.; Yao, W.B.; Ba, Q.; Wang, H. Effects of cadmium exposure on the immune system and immunoregulation. Front. Immunol. 2021, 12, 695484. [Google Scholar] [CrossRef] [PubMed]
- Giaginis, C.; Gatzidou, E.; Theocharis, S. DNA repair systems as targets of cadmium toxicity. Toxicol. Appl. Pharmacol. 2006, 213, 282–290. [Google Scholar] [CrossRef]
- Skipper, A.; Sims, J.N.; Yedjou, C.G.; Tchounwou, P.B. Cadmium chloride induces DNA damage and apoptosis of human liver carcinoma cells via oxidative stress. Int. J. Environ. Res. Public Health 2016, 13, 88. [Google Scholar] [CrossRef]
- Clemens, S.; Ma, J.F. Toxic heavy metal and metalloid accumulation in crop plants and foods. Annu. Rev. Plant Biol. 2016, 67, 489–512. [Google Scholar] [CrossRef]
- Cao, F.B.; Wang, R.F.; Cheng, W.D.; Zeng, F.R.; Ahmed, I.M.; Hu, X.N.; Zhang, G.P.; Wu, F.B. Genotypic and environmental variation in cadmium, chromium, lead and copper in rice and approaches for reducing the accumulation. Sci. Total Environ. 2014, 496, 275–281. [Google Scholar] [CrossRef]
- Ali, U.; Zhong, M.; Shar, T.; Fiaz, S.; Xie, L.H.; Jiao, G.A.; Ahmad, S.; Sheng, Z.H.; Tang, S.Q.; Wei, X.J.; et al. The influence of pH on cadmium accumulation in seedlings of rice (Oryza sativa L.). J. Plant Growth Regul. 2020, 39, 930–940. [Google Scholar] [CrossRef]
- Tang, B.; Luo, M.J.; Zhang, Y.X.; Guo, H.L.; Li, J.N.; Song, W.; Zhang, R.Y.; Feng, Z.; Kong, M.S.; Li, H.; et al. Natural variations in the P-type ATPase heavy metal transporter gene ZmHMA3 control cadmium accumulation in maize grains. J. Exp. Bot. 2021, 72, 6230–6246. [Google Scholar] [CrossRef]
- Xie, L.P.; Hao, P.F.; Cheng, Y.; Ahmed, I.M.; Cao, F.B. Effect of combined application of lead, cadmium, chromium and copper on grain, leaf and stem heavy metal contents at different growth stages in rice. Ecotoxicol. Environ. Saf. 2018, 162, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.F.; Zheng, C.; Hu, Y.T.; Jiang, T.; Liu, Y.; Dong, N.Y.; Yang, J.L.; Zheng, S.J. Cadmium-induced oxalate secretion from root apex is associated with cadmium exclusion and resistance in Lycopersicon esulentum. Plant Cell Environ. 2011, 34, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Liu, Q.; Min, J.; Zhang, S.; Li, S.; Chen, Y.; Dai, J. Specific bacterial communities in the rhizosphere of low-cadmium and high-zinc wheat (Triticum aestivum L.). Sci. Total Environ. 2022, 838, 156484. [Google Scholar] [CrossRef]
- Liu, L.; Qian, Q. Analysis of factors affecting cadmium absorption by crops and countermeasures for soil cadmium pollution. J. Zhejiang Agric. 2005, 17, 111–116. [Google Scholar]
- Zhao, F.J.; Wang, P. Arsenic and cadmium accumulation in rice and mitigation strategies. Plant Soil 2020, 446, 1–21. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Ma, J.F. Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice. J. Exp. Bot. 2014, 65, 6013–6021. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, S.; Cheng, Z.; Li, T.; Jia, Y.; Wang, G.; Yang, Z.; Xian, J.; Yang, Y.; Zhou, W. Transcriptome analysis revealed cadmium accumulation mechanisms in hyperaccumulator Siegesbeckia orientalis L. Environ. Sci. Pollut. Res. 2020, 27, 18853–18865. [Google Scholar] [CrossRef]
- Paola, A.B.; Jonathan, P.R.; Rocío, C.O. Metal and metalloid toxicity in plants: An overview on molecular aspects. Plants 2021, 10, 635. [Google Scholar]
- Huang, W.-X.; Zhang, D.-M.; Cao, Y.-Q.; Dang, B.-J.; Jia, W.; Xu, Z.-C.; Han, D. Differential cadmium translocation and accumulation between Nicotiana tabacum L. and Nicotiana rustica L. by transcriptome combined with chemical form analyses. Ecotoxicol. Environ. Saf. 2021, 208, 111412. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yu, H.; Zhang, X.; Ye, D.; Huang, H.; Wang, Y.; Zheng, Z.; Li, T. A transcriptomic view of cadmium retention in roots of cadmium-safe rice line (Oryza sativa L.). J. Hazard. Mater. 2021, 418, 126379. [Google Scholar] [CrossRef]
- Huang, Y.M.; Chen, H.Q.; Reinfelder, J.R.; Liang, X.Y.; Sun, C.J. A transcriptomic (RNA-seq) analysis of genes responsive to both cadmium and arsenic stress in rice root. Sci. Total Environ. 2019, 666, 445–460. [Google Scholar] [CrossRef]
- Uraguchi, S.; Kamiya, T.; Sakamoto, T.; Kasai, K.; Sato, Y.; Nagamura, Y.; Yoshida, A.; Kyozuka, J.; Ishikawa, S.; Fujiwara, T. Low-affinity cation transporter (OsLCT1) regulates cadmium transport into rice grains. Proc. Natl. Acad. Sci. USA 2011, 108, 20959–20964. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zeng, X.; Song, Q.; Sun, Y.; Feng, Y.; Lai, Y. Identification of key genes and modules in response to Cadmium stress in different rice varieties and stem nodes by weighted gene co-expression network analysis. Hortic. Res. 2020, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Sun, L.; Meng, Q.; Yu, J.; Xiao, G. Phenotypic and genetic dissection of cadmium accumulation in roots, nodes and grains of rice hybrids. Plant Soil 2021, 463, 39–53. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, C.; Du, B.; Lu, B.; Zhou, D.; Zhou, J.; Zhou, J. Effects of node restriction on cadmium accumulation in eight Chinese wheat (Triticum turgidum) cultivars. Sci. Total Environ. 2020, 725, 138358. [Google Scholar] [CrossRef]
- Liu, A.; Zhou, Z.; Yi, Y.; Chen, G. Transcriptome analysis reveals the roles of stem nodes in cadmium transport to rice grain. BMC Genom. 2020, 21, 127. [Google Scholar] [CrossRef]
- Clemens, S.; Aarts, M.G.M.; Thomine, S.; Verbruggen, N. Plant science: The key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef]
- Lekeux, G.; Crowet, J.M.; Nouet, C.; Joris, M.; Jadoul, A.; Bosman, B.; Carnol, M.; Motte, P.; Lins, L.; Galleni, M.; et al. Homology modeling and in vivo functional characterization of the zinc permeation pathway in a heavy metal P-type ATPase. J. Exp. Bot. 2019, 70, 329–341. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Takahashi, R.; Bashir, K.; Shimo, H.; Senoura, T.; Sugimoto, K.; Ono, K.; Yano, M.; Ishikawa, S.; Arao, T.; et al. Characterizing the role of rice NRAMP5 in manganese, iron and cadmium transport. Sci. Rep. 2012, 2, 286. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Bashir, K.; Senoura, T.; Sugimoto, K.; Ono, K.; Suzui, N.; Kawachi, N.; Ishii, S.; et al. From laboratory to field: OsNRAMP5-knockdown rice is a promising candidate for Cd phytoremediation in paddy fields. PLoS ONE 2014, 9, e98816. [Google Scholar] [CrossRef]
- Wu, D.Z.; Yamaji, N.; Yamane, M.; Kashino-Fujii, M.; Sato, K.; Ma, J.F. The HvNramp5 transporter mediates uptake of cadmium and manganese, but not iron. Plant Physiol. 2016, 172, 1899–1910. [Google Scholar] [CrossRef]
- Sui, F.-Q.; Chang, J.-D.; Tang, Z.; Liu, W.-J.; Huang, X.-Y.; Zhao, F.-J. Nramp5 expression and functionality likely explain higher cadmium uptake in rice than in wheat and maize. Plant Soil 2018, 433, 377–389. [Google Scholar] [CrossRef]
- Chang, J.D.; Huang, S.; Yamaji, N.; Zhang, W.; Ma, J.F.; Zhao, F.J. OsNRAMP1 transporter contributes to cadmium and manganese uptake in rice. Plant Cell Environ. 2020, 43, 2476–2491. [Google Scholar] [CrossRef] [PubMed]
- Lei, G.J.; Fujii-Kashino, M.; Wu, D.Z.; Hisano, H.; Saisho, D.; Deng, F.L.; Yamaji, N.; Sato, K.; Zhao, F.-J.; Ma, J.F. Breeding for low cadmium barley by introgression of a Sukkula-like transposable element. Nat. Food 2020, 1, 489–499. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell and Environment 2012, 35, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; You, F.M.; Datla, R.; Ravichandran, S.; Jia, B.S.; Cloutier, S. Genome-wide identification of ATP binding cassette (ABC) transporter and heavy metal associated (HMA) gene families in flax (Linum usitatissimum L.). BMC Genom. 2020, 21, 722. [Google Scholar] [CrossRef]
- Fu, S.; Lu, Y.S.; Zhang, X.; Yang, G.; Chao, D.; Wang, Z.; Shi, M.; Chen, J.; Chao, D.Y.; Li, R. The ABC transporter OsABCG36 is required for Cd tolerance in rice. J. Exp. Bot. 2019, 70, 5909–5918. [Google Scholar] [CrossRef]
- Cai, X.Z.; Wang, M.; Jiang, Y.C.; Wang, C.H.; Ow, D.W. Overexpression of OsABCG48 lowers cadmium in rice (Oryza sativa L.). Agronomy 2021, 11, 918. [Google Scholar] [CrossRef]
- Yang, G.Z.; Fu, S.; Huang, J.J.; Li, L.Y.; Long, Y.; Wei, Q.X.; Wang, Z.G.; Chen, Z.W.; Xia, J.X. The tonoplast-localized transporter OsABCC9 is involved in cadmium tolerance and accumulation in rice. Plant Sci. 2021, 307, 110894. [Google Scholar] [CrossRef]
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, N.K. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Hao, X.H.; Zeng, M.; Wang, J.; Zeng, Z.W.; Dai, J.L.; Xie, Z.J.; Yang, Y.Z.; Tian, L.F.; Chen, L.B.; Li, D.P. A node-expressed transporter OsCCX2 is involved in grain cadmium accumulation of rice. Front. Plant Sci. 2018, 9, 476. [Google Scholar] [CrossRef]
- Liu, S.M.; Jiang, J.; Liu, Y.; Meng, J.; Xu, S.L.; Tan, Y.Y.; Li, Y.F.; Shu, Q.Y.; Huang, J.Z. Characterization and evaluation of OsLCT1 and OsNramp5 mutants generated through CRISPR/Cas9-mediated mutagenesis for breeding low Cd rice. Rice Sci. 2019, 26, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.S.; Feng, S.J.; Zhang, B.Q.; Wang, M.Q.; Cao, H.W.; Rono, J.K.; Chen, X.; Yang, Z.M. OsZIP1 functions as a metal efflux transporter limiting excess zinc, copper and cadmium accumulation in rice. BMC Plant Biol. 2019, 19, 283. [Google Scholar] [CrossRef]
- Yan, H.L.; Xu, W.X.; Xie, J.Y.; Gao, Y.W.; Wu, L.L.; Sun, L.; Feng, L.; Chen, X.; Zhang, T.; Dai, C.H.; et al. Variation of a major facilitator superfamily gene contributes to differential cadmium accumulation between rice subspecies. Nat. Commun. 2019, 10, 2562. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Huang, F.; Luo, R.; Lv, Y.; Ali, U.; Sheng, Z.; Tang, S.; Wei, X.; Hu, P. The effect of cadmium on the microRNAome, degradome and transcriptome of rice seedlings. Plant Growth Regul. 2019, 90, 15–27. [Google Scholar] [CrossRef]
- Chen, Y.H.; Yang, X.Y.; He, K.; Liu, M.H.; Li, J.G.; Gao, Z.F.; Lin, Z.Q.; Zhang, Y.F.; Wang, X.X.; Qiu, X.M.; et al. The MYB transcription factor superfamily of Arabidopsis: Expression analysis and phylogenetic comparison with the rice MYB family. Plant Mol. Biol. 2006, 60, 107–124. [Google Scholar]
- Zhang, P.; Wang, R.L.; Ju, Q.; Li, W.Q.; Tran, L.S.P.; Xu, J. The R2R3-MYB transcription factor MYB49 regulates cadmium accumulation. Plant Physiol. 2019, 180, 529–542. [Google Scholar] [CrossRef]
- Hu, S.B.; Yu, Y.; Chen, Q.H.; Mu, G.M.; Shen, Z.G.; Zheng, L.Q. OsMYB45 plays an important role in rice resistance to cadmium stress. Plant Sci. 2017, 264, 1–8. [Google Scholar] [CrossRef]
- Shim, D.; Hwang, J.-U.; Lee, J.; Lee, S.; Choi, Y.; An, G.; Martinoia, E.; Lee, Y. Orthologs of the class A4 heat shock transcription factor HsfA4a confer cadmium tolerance in wheat and rice. Plant Cell 2009, 21, 4031–4043. [Google Scholar] [CrossRef]
- Jia, Z.Z.; Li, M.Z.; Wang, H.C.; Zhu, B.; Gu, L.; Du, X.Y.; Ren, M.J. TaWRKY70 positively regulates TaCAT5 enhanced Cd tolerance in transgenic Arabidopsis. Environ. Exp. Bot. 2021, 190, 104591. [Google Scholar] [CrossRef]
- Hong, C.Y.; Cheng, D.; Zhang, G.Q.; Zhu, D.D.; Chen, Y.H.; Tan, M.P. The role of ZmWRKY4 in regulating maize antioxidant defense under cadmium stress. Biochem. Biophys. Res. Commun. 2017, 482, 1504–1510. [Google Scholar] [CrossRef]
- Broadley, M.R.; Willey, N.J.; Wilkins, J.C.; Baker, A.J.M.; Mead, A.; White, P.J. Phylogenetic variation in heavy metal accumulation in angiosperms. New Phytol. 2001, 152, 9–27. [Google Scholar] [CrossRef]
- Djemal, R.; Khoudi, H. The ethylene-responsive transcription factor of durum wheat, TdSHN1, confers cadmium, copper, and zinc tolerance to yeast and transgenic tobacco plants. Protoplasma 2022, 259, 19–31. [Google Scholar] [CrossRef]
- Hu, S.; Shinwari, K.I.; Song, Y.; Xia, J.; Xu, H.; Du, B.; Luo, L.; Zheng, L. OsNAC300 positively regulates cadmium stress responses and tolerance in rice roots. Agronomy 2021, 11, 95. [Google Scholar] [CrossRef]
- Du, X.Y.; He, F.; Zhu, B.; Ren, M.J.; Tang, H. NAC transcription factors from Aegilops markgrafii reduce cadmium concentration in transgenic wheat. Plant Soil 2020, 449, 39–50. [Google Scholar] [CrossRef]
- Hu, J.; Cai, J.; Park, S.J.; Lee, K.; Li, Y.X.; Chen, Y.; Yun, J.Y.; Xu, T.; Kang, H. N-6-Methyladenosine mRNA methylation is important for salt stress tolerance in Arabidopsis. Plant J. 2021, 106, 1759–1775. [Google Scholar] [CrossRef]
- Finnegan, E.J.; Peacock, W.J.; Dennis, E.S. Reduced DNA methylation in Arabidopsis thaliana results in abnormal plant de-velopment. Proc. Natl. Acad. Sci. USA 1996, 93, 8449–8454. [Google Scholar] [CrossRef]
- Sun, D.-D.; Sun, J.-W.; Huang, L.-Y.; Chen, N.; Wang, Q.-W. Effects of cadmium stress on DNA methylation in soybean. Biotechnol. Biotechnol. Equip. 2021, 35, 1696–1705. [Google Scholar] [CrossRef]
- Shafiq, S.; Zeb, Q.; Ali, A.; Sajjad, Y.; Nazir, R.; Widemann, E.; Liu, L.Y. Lead, cadmium and zinc phytotoxicity alter DNA methylation levels to confer heavy metal tolerance in wheat. Int. J. Mol. Sci. 2019, 20, 4676. [Google Scholar] [CrossRef]
- Fan, S.K.; Ye, J.Y.; Zhang, L.L.; Chen, H.S.; Zhang, H.H.; Zhu, Y.X.; Liu, X.X.; Jin, C.W. Inhibition of DNA demethylation enhances plant tolerance to cadmium toxicity by improving iron nutrition. Plant Cell Environ. 2020, 43, 275–291. [Google Scholar] [CrossRef]
- Su, T.T.; Fu, L.B.; Kuang, L.H.; Chen, D.Y.; Zhang, G.P.; Shen, Q.F.; Wu, D.Z. Transcriptome-wide m6A methylation profile reveals regulatory networks in roots of barley under cadmium stress. J. Hazard. Mater. 2022, 423, 127140. [Google Scholar] [CrossRef]
- Cheng, Q.; Wang, P.; Wu, G.L.; Wang, Y.N.; Tan, J.G.; Li, C.J.; Zhang, X.Y.; Liu, S.L.; Huang, S.Y.; Huang, T.; et al. Coordination of m6A mRNA methylation and gene transcriptome in rice response to cadmium stress. Rice 2021, 14, 62. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.F.; Gong, S.H.; Wang, Y.; Wang, F.J.; Bao, H.X.G.D.L.; Sun, J.W.; Cai, C.; Yi, K.K.; Chen, Z.X.; Zhu, C. MicroRNA166 modulates cadmium tolerance and accumulation in rice. Plant Physiol. 2018, 177, 1691–1703. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Kumar, V.; Khare, T.; Tripathi, P.; Shah, T.; Ramakrishna, C.; Aglawe, S.; Mangrauthia, S.K. miRNA applications for engineering abiotic stress tolerance in plants. Biologia 2020, 75, 1063–1081. [Google Scholar] [CrossRef]
- Zhou, M.; Zheng, S.G.; Liu, R.; Lu, L.; Zhang, C.H.; Zhang, L.; Yant, L.; Wu, Y. The genome-wide impact of cadmium on microRNA and mRNA expression in contrasting Cd responsive wheat genotypes. BMC Genom. 2019, 20, 615. [Google Scholar] [CrossRef]
- Tang, Z.; Cai, H.; Li, J.; Lv, Y.; Zhang, W.; Zhao, F.-J. Allelic variation of NtNramp5 associated with cultivar variation in cadmium accumulation in tobacco. Plant Cell Physiol. 2017, 58, 1583–1593. [Google Scholar] [CrossRef]
- Liedschulte, V.; Laparra, H.; Battey, J.N.D.; Schwaar, J.D.; Broye, H.; Mark, R.; Klein, M.; Goepfert, S.; Bovet, L. Impairing both HMA4 homeologs is required for cadmium reduction in tobacco. Plant Cell Environ. 2021, 40, 364–377. [Google Scholar] [CrossRef]
- Liu, W.; Huo, C.; He, L.; Ji, X.; Yu, T.; Yuan, J.; Zhou, Z.; Song, L.; Yu, Q.; Chen, N.; et al. The NtNRAMP1 transporter is involved in cadmium and iron transport in tobacco (Nicotiana tabacum). Plant Physiol. Biochem. 2022, 173, 59–67. [Google Scholar] [CrossRef]
- Jia, H.F.; Yin, Z.R.; Xuan, D.D.; Lian, W.L.; Han, D.; Zhu, Z.T.; Li, C.Y.; Li, C.; Song, Z.P. Mutation of NtNRAMP3 improves cadmium tolerance and its accumulation in tobacco leaves by regulating the subcellular distribution of cadmium. J. Hazard. Mater. 2022, 432, 128701. [Google Scholar] [CrossRef]
- Zhang, H.A.; Xu, Z.A.; Huo, Y.A.; Guo, K.A.; Wang, Y.B.; He, G.C.; Sun, H.C.; Li, M.A.; Li, X.A.; Xu, N.D. Overexpression of TrxCDSP32 gene promotes chlorophyll synthesis and photosynthetic electron transfer and alleviates cadmium-induced photoinhibition of PSII and PSI in tobacco leaves. J. Hazard. Mater. 2020, 398, 122899. [Google Scholar] [CrossRef]
- Gong, X.; Yin, L.; Chen, J.; Guo, C. Overexpression of the iron transporter NtPIC1 in tobacco mediates tolerance to cadmium. Plant Cell Rep. 2015, 34, 1963–1973. [Google Scholar] [CrossRef]
- Shimo, H.; Ishimaru, Y.; An, G.; Yamakawa, T.; Nakanishi, H.; Nishizawa, N.K. Low cadmium (LCD), a novel gene related to cadmium tolerance and accumulation in rice. J. Exp. Bot. 2011, 62, 5727–5734. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Kakei, Y.; Shimo, H.; Bashir, K.; Sato, Y.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. A rice phenolic efflux transporter is essential for solubilizing precipitated apoplasmic iron in the plant stele. J. Biol. Chem. 2011, 286, 24649–24655. [Google Scholar] [CrossRef]
- Tang, L.; Mao, B.G.; Li, Y.K.; Lv, Q.M.; Zhang, L.P.; Chen, C.Y.; He, H.J.; Wang, W.P.; Zeng, X.F.; Shao, Y.; et al. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H.; et al. OsHMA3, a P-1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef]
- Ding, Y.F.; Ye, Y.Y.; Jiang, Z.H.; Wang, Y.; Zhu, C. MicroRNA390 is involved in cadmium tolerance and accumulation in rice. Front. Plant Sci. 2016, 7, 235. [Google Scholar] [CrossRef]
- Lee, B.R.; Hwang, S. Over-expression of NtHb1 encoding a non-symbiotic class 1 hemoglobin of tobacco enhances a tolerance to cadmium by decreasing NO (nitric oxide) and Cd levels in Nicotiana tabacum. Environ. Exp. Bot. 2015, 113, 18–27. [Google Scholar] [CrossRef]
- Bahmani, R.; Kim, D.; Lee, B.D.; Hwang, S. Over-expression of tobacco UBC1 encoding a ubiquitin-conjugating enzyme increases cadmium tolerance by activating the 20S/26S proteasome and by decreasing Cd accumulation and oxidative stress in tobacco (Nicotiana tabacum). Plant Mol. Biol. 2017, 94, 433–451. [Google Scholar] [CrossRef]
- Bahmani, R.; Modareszadeh, M.; Kim, D.; Hwang, S. Overexpression of tobacco UBQ2 increases Cd tolerance by decreasing Cd accumulation and oxidative stress in tobacco and Arabidopsis. Environ. Exp. Bot. 2019, 166, 103805. [Google Scholar] [CrossRef]
- Li, C.; Zhang, T.; Peng, P.; Li, D.; Brestic, M.; Liu, Y.; Yang, X. Genetic engineering of glycinebetaine synthesis enhances cadmium tolerance in BADH-transgenic tobacco plants via reducing cadmium uptake and alleviating cadmium stress damage. Environ. Exp. Bot. 2021, 191, 104602. [Google Scholar] [CrossRef]
- Kobayashi, E.; Suwazono, Y.; Honda, R.; Dochi, M.; Nishijo, M.; Kido, T.; Nakagawa, H. Changes in renal tubular and glomerular functions and biological acid-base balance after soil replacement in Cd-polluted rice paddies calculated with a general linear mixed model. Biol. Trace Elem. Res. 2008, 124, 164–172. [Google Scholar] [CrossRef]
- Hseu, Z.-Y.; Su, S.-W.; Lai, H.-Y.; Guo, H.-Y.; Chen, T.-C.; Chen, Z.-S. Remediation techniques and heavy metal uptake by different rice varieties in metal-contaminated soils of Taiwan: New aspects for food safety regulation and sustainable agriculture. Soil Sci. Plant Nutr. 2010, 56, 31–52. [Google Scholar] [CrossRef]
- Ren, Y.; Abbood, H.A.; He, F.B.; Peng, H.; Huang, K.X. Magnetic EDTA-modified chitosan/SiO2/Fe3O4 adsorbent: Preparation, characterization, and application in heavy metal adsorption. Chem. Eng. J. 2013, 226, 300–311. [Google Scholar] [CrossRef]
- Dong, J.; Sun, Q.; Zhang, X.; Zhou, Y.; Xia, L.C.; Yuan, J. Effect of soil washing with ferric chloride on cadmium removal and soil structure. Appl. Sci. 2021, 11, 10956. [Google Scholar] [CrossRef]
- Hussain, F.; Eom, H.; Toor, U.A.; Lee, C.S.; Oh, S.-E. Rapid assessment of heavy metal-induced toxicity in water using micro-algal bioassay based on photosynthetic oxygen evolution. Environ. Eng. Res. 2021, 26, 200391. [Google Scholar] [CrossRef]
- Honma, T.; Ohba, H.; Kaneko-Kadokura, A.; Makino, T.; Nakamura, K.; Katou, H. Optimal soil Eh, pH, and water management for simultaneously minimizing arsenic and cadmium concentrations in rice grains. Environ. Sci. Technol. 2016, 50, 4178–4185. [Google Scholar] [CrossRef]
- Jalloh, M.A.; Chen, J.H.; Zhen, F.R.; Zhang, G.P. Effect of different N fertilizer forms on antioxidant capacity and grain yield of rice growing under Cd stress. J. Hazard. Mater. 2009, 162, 1081–1085. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, W.; Xu, S.; Shi, H.; Wen, D.; Huang, Y.; Peng, L.; Deng, T.; Du, R.; Li, F.; et al. Increasing ammonium nutrition as a strategy for inhibition of cadmium uptake and xylem transport in rice (Oryza sativa L.) exposed to cadmium stress. Environ. Exp. Bot. 2018, 155, 734–741. [Google Scholar] [CrossRef]
- Chen, F.; Dong, J.; Wang, F.; Wu, F.B.; Zhang, G.P.; Li, G.M.; Chen, Z.F.; Chen, J.X.; Wei, K. Identification of barley genotypes with low grain Cd accumulation and its interaction with four microelements. Chemosphere 2007, 67, 2082–2088. [Google Scholar] [CrossRef]
- de Oliveira, V.F.; Busanello, C.; Viana, V.E.; Stafen, C.F.; Pedrolo, A.M.; Paniz, F.P.; Pedron, T.; Pereira, R.M.; Rosa, S.A.; Junior, A.M.D.M.; et al. Assessing mineral and toxic elements content in rice grains grown in southern Brazil. J. Food Compos. Anal. 2021, 100, 103914. [Google Scholar] [CrossRef]
- Sabella, E.; Luvisi, A.; Genga, A.; De Bellis, L.; Aprile, A. Molecular responses to cadmium exposure in two contrasting durum wheat genotypes. Int. J. Mol. Sci. 2021, 22, 7343. [Google Scholar] [CrossRef]
- Ayachi, I.; Ghabriche, R.; Kourouma, Y.; Ben Naceur, M.; Abdelly, C.; Thomine, S.; Ghnaya, T. Cd tolerance and accumulation in barley: Screening of 36 North African cultivars on Cd-contaminated soil. Environ. Sci. Pollut. Res. 2021, 28, 42722–42736. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, R.G.; Tang, W.B.; Chen, Y.C.; Zhou, J.Q.; Ma, H.R.; Li, S.; Deng, H.B.; Han, L.; Chen, Y.B.; et al. Robust identification of low-Cd rice varieties by boosting the genotypic effect of grain Cd accumulation in combination with marker-assisted selection. J. Hazard. Mater. 2022, 424, 127703. [Google Scholar] [CrossRef] [PubMed]
- Kong, Z.Y.; Glick, B.R. The role of plant growth-promoting bacteria in metal phytoremediation. Adv. Microb. Physiol. 2017, 71, 97–132. [Google Scholar] [PubMed]
- Jabeen, R.; Ahmad, A.; Iqbal, M.F. Phytoremediation of heavy metals: Physiological and molecular mechanisms. Bot. Rev. 2009, 75, 339–364. [Google Scholar] [CrossRef]
- Schwartz, C.; Echevarria, G.; Morel, J.L. Phytoextraction of cadmium with Thlaspi caerulescens. Plant Soil 2003, 249, 27–35. [Google Scholar] [CrossRef]
- Milner, M.J.; Mitani–Ueno, N.; Yamaji, N.; Yokosho, K.; Craft, E.; Fei, Z.J.; Ebbs, S.; Zambrano, M.C.; Ma, J.F.; Kochian, L.V. Root and shoot transcriptome analysis of two ecotypes of Noccaea caerulescens uncovers the role of NcNramp1 in Cd hyperaccumulation. Plant J. 2014, 78, 398–410. [Google Scholar] [CrossRef]
- Li, C.F.; Zhou, K.H.; Qin, W.Q.; Tian, C.J.; Qi, M.; Yan, X.M.; Han, W.B. A review on heavy metals contamination in soil: Effects, sources, and remediation techniques. Soil Sediment Contam. 2019, 28, 380–394. [Google Scholar] [CrossRef]
- Hou, D.D.; Wang, R.Z.; Gao, X.Y.; Wang, K.; Lin, Z.; Ge, J.; Liu, T.; Wei, S.; Chen, W.K.; Xie, R.H.; et al. Cultivar-specific response of bacterial community to cadmium contamination in the rhizosphere of rice (Oryza sativa L.). Environ. Pollut. 2018, 241, 63–73. [Google Scholar] [CrossRef]
- Sun, L.N.; Wu, B.B.; Xu, Z.H.; Li, Z.L.; Zhang, S.Q.; Yang, E.D.; Zhou, J.H. Rhizosphere microbial community structure and functional diversity in two Cd-accumulating wheat cultivars. Acta Ecol. Sin. 2022, 42, 3366–3376. [Google Scholar]
- Huang, H.L.; Li, M.; Rizwan, M.; Dai, Z.H.; Yuan, Y.; Hossain, M.M.; Cao, M.H.; Xiong, S.L.; Tu, S.X. Synergistic effect of silicon and selenium on the alleviation of cadmium toxicity in rice plants. J. Hazard. Mater. 2021, 401, 123393. [Google Scholar] [CrossRef]
- Wang, T.F.; Zhang, K.; Lu, L.; Xiao, X.; Chen, B.L. Novel insights into effects of silicon-rich biochar (Sichar) amendment on cadmium uptake, translocation and accumulation in rice plants. Environ. Pollut. 2020, 265, 114772. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.W.; Wise, B.E. Mitigating cadmium accumulation in spinach and onions by the application of silicon fertilizer to soil. Soil Sediment Contam. 2020, 29, 532–544. [Google Scholar] [CrossRef]
- Rodriguez-Serrano, M.; Romero-Puertas, M.C.; Zabalza, A.; Corpas, F.J.; Gomez, M.; Del Rio, L.A.; Sandalio, L.M. Cadmium effect on oxidative metabolism of pea (Pisum sativum L.) roots. Imaging of reactive oxygen species and nitric oxide accumulation in vivo. Plant Cell Environ. 2006, 29, 1532–1544. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.T.; Chen, H.P.; Zhu, K.K.; Jin, Q.J.; Xie, Y.J.; Cui, J.; Xia, Y.; Zhang, J.; Shen, W.B. Cadmium-induced hydrogen sulfide synthesis is involved in cadmium tolerance in medicago sativa by reestablishment of reduced (Homo) glutathione and reactive oxygen species homeostasis. PLoS ONE 2014, 9, e109669. [Google Scholar]
- Kapoor, B.; Kumar, P.; Gill, N.S.; Sharma, R.; Thakur, N.; Irfan, M. Molecular mechanisms underpinning the silicon-selenium (Si-Se) interactome and cross-talk in stress-induced plant responses. Plant Soil 2022, 1–24. [Google Scholar] [CrossRef]

| Gene Name | Materials | Environment | Treatment | Reference |
|---|---|---|---|---|
| NtHMA4.1, NtHMA4.2 | tobacco RNAi lines, EMS mutant | glasshouse, field trial | 1 μM Cd | [90] |
| NtNramp1 | heterologous expression in yeast, transgenic tobacco | glasshouse, synthetic medium | 20, 50 μM Cd | [91] |
| NtNramp3 | Knock-out lines | glasshouse | 5, 20 and 50 μM Cd | [92] |
| NtNramp5 | heterologous expression in yeast | glasshouse, synthetic medium | 0.1, 5 μM Cd | [89] |
| NtHb1 | transgenic tobacco KO Arabidopsis | 1/2 MS agar | 50 μM | [100] |
| NtUBC1 | transgenic tobacco | 1/2 MS agar | 50 μM | [101] |
| NtUBQ2 | transgenic tobacco, Arabidopsis mutant | 1/2 MS agar | 50 μM | [102] |
| NtBADH | transgenic tobacco | MS medium, greenhouse | 0.2, 0.5 mM | [103] |
| TrxCDSP32 | transgenic tobacco | artificial culture chamber | 100 μM | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, S.; Lin, K.; Williams, D.V.; Liu, Y.; Dai, H.; Cao, F. Cadmium Accumulation in Cereal Crops and Tobacco: A Review. Agronomy 2022, 12, 1952. https://doi.org/10.3390/agronomy12081952
Mei S, Lin K, Williams DV, Liu Y, Dai H, Cao F. Cadmium Accumulation in Cereal Crops and Tobacco: A Review. Agronomy. 2022; 12(8):1952. https://doi.org/10.3390/agronomy12081952
Chicago/Turabian StyleMei, Shineng, Kaina Lin, Darron V. Williams, Yang Liu, Huaxin Dai, and Fangbin Cao. 2022. "Cadmium Accumulation in Cereal Crops and Tobacco: A Review" Agronomy 12, no. 8: 1952. https://doi.org/10.3390/agronomy12081952