
The Physiological Role of Abscisic Acid in Regulating Root System Architecture of Alfalfa in Its Adaptation to Water Deficit



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Materials
2.2. Experiment 1
2.2.1. Experimental Description and Design
2.2.2. Soil Water-Holding Capacity
2.2.3. Measurements
Herbage Accumulation
Growing Point Density
Root and Residual Shoot Biomass
2.3. Experiment 2
2.3.1. Experimental Description and Design
2.3.2. Root Sampling and Scanning
2.3.3. Measurements
Root-Sourced ABA Content
Topological Index
Fractal Analysis
2.4. Statistical Analyses
3. Results
3.1. Experiment 1
3.1.1. Herbage Accumulation
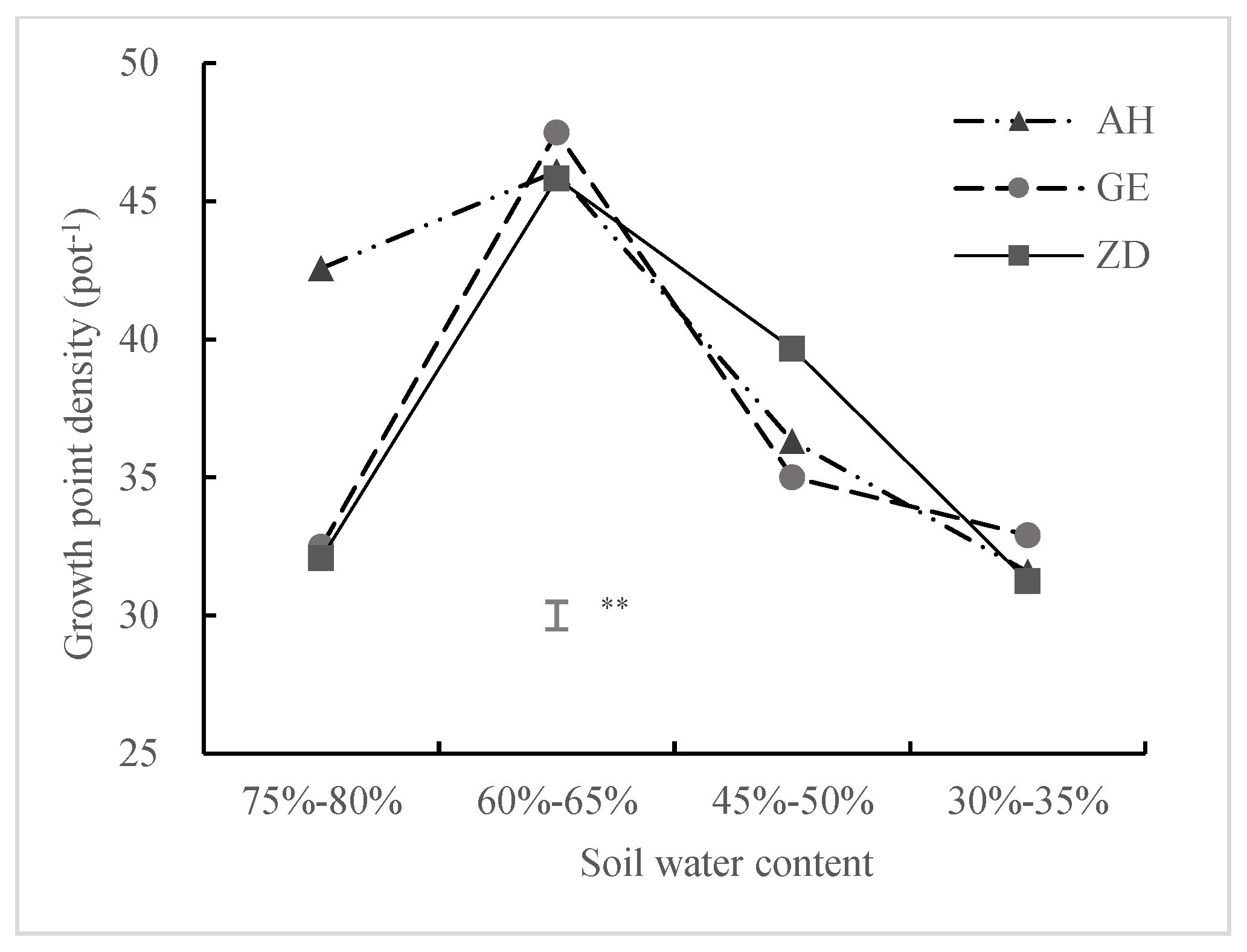
3.1.2. Growing Point Density
3.1.3. Residual Shoot Biomass and Root Biomass
3.2. Experiment 2
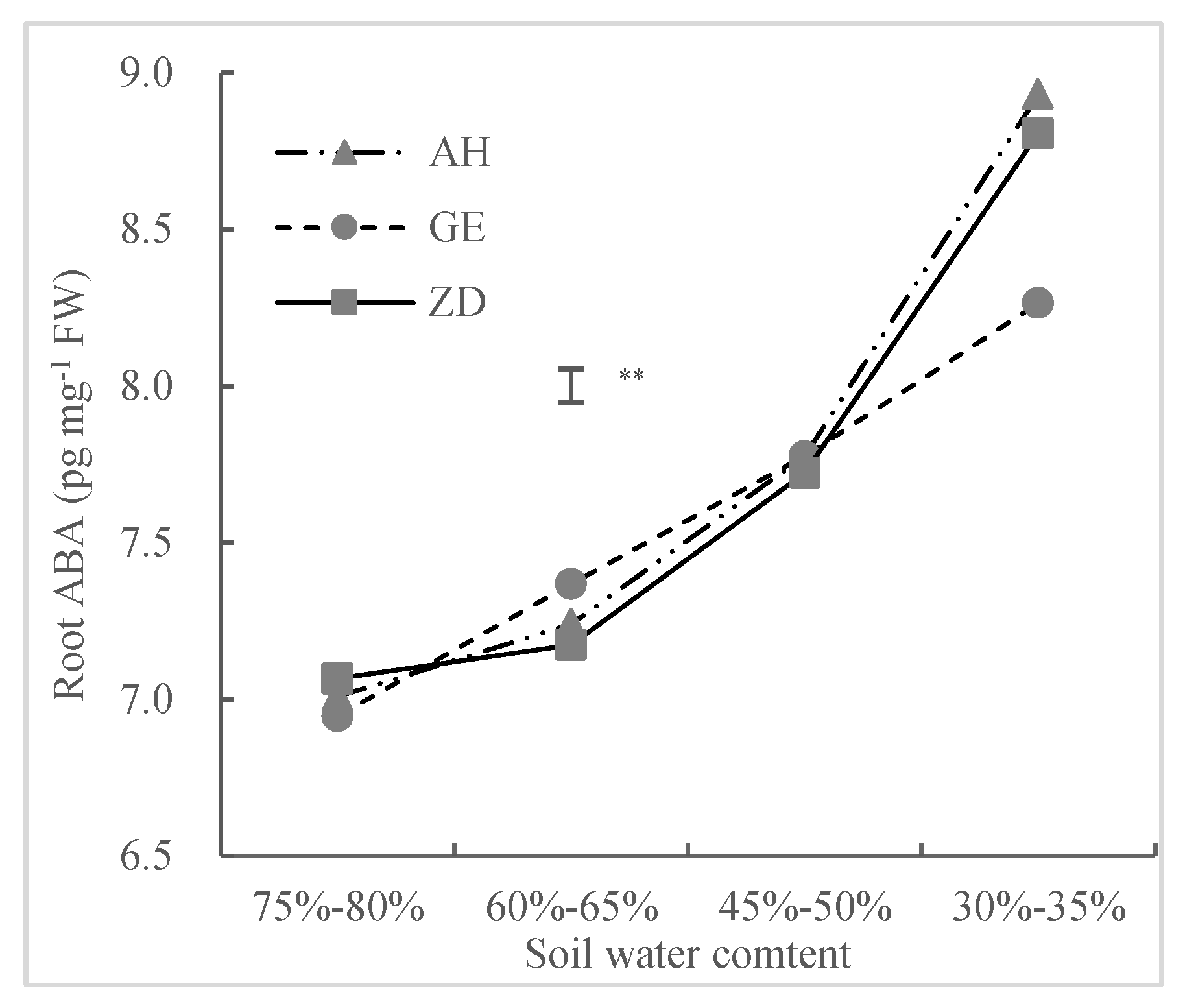
3.2.1. Root-Sourced ABA Content
3.2.2. Root Morphology, Topology, and Root Fractal Analysis
3.2.3. Root Biomass and Herbage Biomass
3.2.4. Modelling Analysis of Root ABA Content, Root System Architecture, and Plant Biomass
4. Discussion
4.1. Effect of Water Stress
4.2. Abscisic Acid in Responses to Water Stress
4.3. Correlation among Root ABA, Root System Architecture, and Plant Biomass
4.4. Practical Implications and Further Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Popp, J.D.; Mccaughey, W.P.; Cohen, R.; Mcallister, T.A.; Majak, W. Enhancing pasture productivity with alfalfa: A review. Rev. Can. Phytotech. 2000, 80, 513–519. [Google Scholar] [CrossRef]
- Deng, L.; Wang, K.-B.; Li, J.-P.; Shangguan, Z.-P.; Sweeney, S. Carbon Storage Dynamics in Alfalfa (Medicago sativa) Fields in the Hilly-Gully Region of the Loess Plateau, China. CLEAN—Soil Air Water 2014, 42, 1253–1262. [Google Scholar] [CrossRef]
- Li, S.; Wan, L.; Nie, Z.; Li, X. Fractal and Topological Analyses and Antioxidant Defense Systems of Alfalfa (Medicago sativa L.) Root System under Drought and Rehydration Regimes. Agronomy 2020, 10, 805. [Google Scholar] [CrossRef]
- Bao, A.-K.; Guo, Z.-G.; Zhang, H.-F.; Wang, S.-M. A procedure for assessing the salt tolerance of lucerne (Medicago sativa L.) cultivar seedlings by combining agronomic and physiological indicators. N. Z. J. Agric. Res. 2009, 52, 435–442. [Google Scholar]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2010, 31, 11–38. [Google Scholar] [CrossRef]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.V.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef] [Green Version]
- Deak, A.; Hall, M.H.; Sanderson, M.A. Grazing Schedule Effect on Forage Production and Nutritive Value of Diverse Forage Mixtures. Agron. J. 2009, 101, 408–414. [Google Scholar] [CrossRef] [Green Version]
- Burke, M.B.; Lobell, D.B.; Guarino, L. Shifts in African crop climates by 2050, and the implications for crop improvement and genetic resources conservation. Glob. Environ. Change 2009, 19, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Saini, H.S.; Westgate, M.E. Reproductive Development in Grain Crops during Drought. Adv. Agron. 1999, 68, 59–96. [Google Scholar]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Li, M.; Li, S.; Li, Y. Studies on Drought in the Past 50 Years in China. Agric. Meteorol. 2003, 24, 7–10. [Google Scholar]
- Young, L.W.; Wilen, R.W.; Bonham-Smith, P.C. High temperature stress of Brassica napus during flowering reduces micro- and megagametophyte fertility, induces fruit abortion, and disrupts seed production. J. Exp. Bot. 2004, 55, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Danjon, F.; Reubens, B. Assessing and analyzing 3D architecture of woody root systems, a review of methods and applications in tree and soil stability, resource acquisition and allocation. Plant Soil 2008, 303, 1–34. [Google Scholar] [CrossRef]
- Wu, W.; Duncan, R.W.; Ma, B.-L. Quantification of canola root morphological traits under heat and drought stresses with electrical measurements. Plant Soil 2017, 415, 229–244. [Google Scholar] [CrossRef]
- Colombi, T.; Walter, A. Root responses of triticale and soybean to soil compaction in the field are reproducible under controlled conditions. Funct. Plant Biol. 2015, 43, 114–128. [Google Scholar] [CrossRef]
- Atkinson, J.A.; Pound, M.P.; Bennett, M.J.; Wells, D.M. Uncovering the hidden half of plants using new advances in root phenotyping. Curr. Opin. Biotechnol. 2018, 55, 1–8. [Google Scholar] [CrossRef]
- Grzesiak, M.T.; Hura, K.; Jurczyk, B.; Hura, T.; Rut, G.; Szczyrek, P.; Grzesiak, S.A. Physiological markers of stress susceptibility in maize and triticale under different soil compactions and/or soil water contents. J. Plant Interact. 2017, 12, 355–372. [Google Scholar] [CrossRef] [Green Version]
- Colombi, T.; Kirchgessner, N.; Walter, A.; Keller, T. Root Tip Shape Governs Root Elongation Rate under Increased Soil Strength. Plant Physiol. 2017, 174, 2289–2301. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.; Craig, B.; James, S. Whole root system water conductance responds to both axial and radial traits and network topology over natural range of trait variation. J. Theor. Biol. 2018, 456, 49–61. [Google Scholar]
- Berntson, G.M. Topological scaling and plant root system architecture: Developmental and functional hierarchies. New Phytol. 1997, 135, 621–634. [Google Scholar] [CrossRef]
- Berntson, G.M. The Characterization of Topology: A Comparison of Four topological Indices for Rooted Binary Trees. J. Theor. Biol. 1995, 177, 271–281. [Google Scholar] [CrossRef]
- Kai, L.N.; Lynch, J.P.; Weiss, H.N. Fractal Geometry of Bean Root Systems: Correlations between Spatial and Fractal Dimension. Am. J. Bot. 1997, 84, 26–33. [Google Scholar]
- Ketipearachchi, K.W.; Tatsumi, J. Local Fractal Dimensions and Multifractal Analysis of the Root System of Legumes. Plant Prod. Sci. 2000, 3, 289–295. [Google Scholar] [CrossRef]
- Giehl, R.; Wiren, N.V. Root Nutrient Foraging. Plant Physiol. 2014, 166, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, J.A.; Rasmussen, A.; Traini, R.; Voß, U.; Sturrock, C. Branching Out in Roots: Uncovering Form, Function, and Regulation1. Plant Physiol. 2014, 166, 538–550. [Google Scholar] [CrossRef] [Green Version]
- Ljung, K.; Nemhauser, J.L.; Perata, P. New mechanistic links between sugar and hormone signaling networks. Curr. Opin. Plant Biol. 2015, 25, 130–137. [Google Scholar] [CrossRef]
- Yu, P.; Gutjahr, C.; Li, C.; Hochholdinger, F. Genetic Control of Lateral Root Formation in Cereals. Trends Plant Sci. 2016, 21, 951–961. [Google Scholar] [CrossRef]
- Dodd, I.C. Root-To-Shoot Signalling: Assessing the Roles of ‘Up’ In the Up and Down World of Long-Distance Signaling in Planta. Plant Soil 2005, 274, 251–270. [Google Scholar] [CrossRef]
- Ikegami, K.; Okamoto, M.; Seo, M.; Koshiba, T. Activation of abscisic acid biosynthesis in the leaves of Arabidopsis thaliana in response to water deficit. J. Plant Res. 2009, 122, 235–243. [Google Scholar] [CrossRef]
- Thole, J.M.; Beisner, E.R.; Liu, J.; Venkova, S.V.; Strader, L.C. Abscisic Acid Regulates Root Elongation Through the Activities of Auxin and Ethylene in Arabidopsis thaliana. G3 2014, 4, 1259–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowe, J.H.; Topping, J.F.; Liu, J.; Lindsey, K. Abscisic acid regulates root growth under osmotic stress conditions via an interacting hormonal network with cytokinin, ethylene and auxin. New Phytol. 2016, 211, 225–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.-P.; Sussmilch, F.; Nichols, D.S.; Cardoso, A.A.; Brodribb, T.J.; McAdam, S.A.M. Leaves, not roots or floral tissue, are the main site of rapid, external pressure-induced ABA biosynthesis in angiosperms. J. Exp. Bot. 2018, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Jeon, E.; Kang, N.Y.; Cho, C.; Seo, P.J.; Suh, M.C.; Kim, J. LBD14/ASL17 Positively Regulates Lateral Root Formation and is Involved in ABA Response for Root Architecture in Arabidopsis. Plant Cell Physiol. 2017, 58, 2190–2201. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2014, 72, 673–689. [Google Scholar] [CrossRef]
- Li, S.; Nie, Z.; Zhang, D. Competition between cocksfoot (Dactylis glomerata L.) and companion species: Evidence for allelopathy. Field Crops Res. 2016, 196, 452–462. [Google Scholar] [CrossRef]
- Li, Z.S.; Wan, L.; Li, S.; Li, X.; He, F.; Tong, Z. Plastic Response of Medicago Sativa L. Root System Traits and Cold Resistance to Simulated Rainfall Events. PeerJ 2021, 9, 11962. [Google Scholar] [CrossRef]
- Nie, Z.N.; Zollinger, R.P.; Jacobs, J.L. Performance of 7 Australian native grasses from the temperate zone under a range of cutting and fertiliser regimes. Crop Pasture Sci. 2009, 60, 943–953. [Google Scholar] [CrossRef]
- Cao, X.; Zhu, C.; Zhong, C.; Hussain, S.; Zhu, L.; Wu, L.; Jin, Q. Mixed-nitrogen nutrition-mediated enhancement of drought tolerance of rice seedlings associated with photosynthesis, hormone balance and carbohydrate partitioning. Plant Growth Regul. 2018, 20, 451–465. [Google Scholar] [CrossRef]
- Fitter, A.H. An architectural approach to the comparative ecology of plant root systems. New Phytol. 1987, 106, 61–77. [Google Scholar]
- Tanaka, M.; Kayama, A.; Kato, R.; Ito, Y. Estimation of the fractal dimension of feature surface patterns by box counting method. Fractals 2003, 7, 335–340. [Google Scholar] [CrossRef]
- Fitter, A.H.; Stickland, T.R. Fractal Characterization of Root System Architecture. Funct. Ecol. 1992, 6, 632–635. [Google Scholar] [CrossRef]
- Gramshaw, D.; Bishop, H.G.; Ludke, D.H. Performance of grazed creeping-rooted lucernes on two soils in central Queensland. Aust. J. Exp. Agric. 1982, 22, 177–181. [Google Scholar] [CrossRef]
- Shan, L.S.; Zhang, X.M.; Wang, Y.K.; Wang, H.; Yan, H.N.; Wei, J.; Xu, H. Influence of moisture on the growth and biomass allocation in Haloxylon ammodendron and Tamarix ramosissima seedlings in the shelterbelt along the Tarim Desert Highway, Xinjiang, China. Chin. Sci. Bull. 2008, 53, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Nie, Z.N.; Miller, S.; Moore, G.A.; Hackney, B.F.; Boschma, S.P.; Reed, K.F.M.; Mitchell, M.; Albertsen, T.O.; Clark, S.; Craig, A.D.; et al. Field evaluation of perennial grasses and herbs in southern Australia. 2. Persistence, root characteristics and summer activity. Aust. J. Exp. Agric. 2008, 48, 424–435. [Google Scholar] [CrossRef]
- Passioura, J.B.; Angus, J.F. Improving Productivity of Crops in Water-limited Environments. Adv. Agron. 2010, 106, 37–75. [Google Scholar]
- Wasson, A.P.; Richards, R.A.; Chatrath, R.; Misra, S.C.; Prasad, S.V.S.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef] [Green Version]
- Grzesiak, M.T.; Hordyńska, N.; Maksymowicz, A.; Grzesiak, S.; Szechyńska-Hebda, M. Variation Among Spring Wheat (Triticum aestivum L.) Genotypes in Response to the Drought Stress. II—Root System Structure. Plants 2019, 8, 584. [Google Scholar] [CrossRef] [Green Version]
- Fitter, A.H. The topology and geometry of plant-root systems—Influence of watering rate on root-system topology in trifolium-pratense. Ann. Bot. 1986, 58, 91–101. [Google Scholar] [CrossRef]
- Fitter, A.H.; Stickland, T.R.; Harvey, M.L.; Wilson, G.W. Architectural analysis of plant root systems 1. Architectural correlates of exploitation efficiency. New Phytol. 1991, 118, 375–382. [Google Scholar] [CrossRef]
- Oppelt, A.L.; Winfried, K.; Godbold, D.L. Topology, scaling relations and Leonardo’s rule in root systems from African tree species. Tree Physiol. 2001, 21, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Day, S.D.; Wiseman, P.E.; Dickinson, S.B.; Harris, J.R. Tree Root Ecology in the Urban Environment and Implications for a Sustainable Rhizosphere. Arboric. Urban For. 2010, 36, 193–205. [Google Scholar] [CrossRef]
- Berntson, G.M.; Stoll, P. Correcting for Finite Spatial Scales of Self–similarity When Calculating Fractal Dimensions of Real–world Structures. Proc. R. Soc. B Biol. Sci. 1997, 264, 1531–1537. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Zhao, X.; Fang, Y.; Xu, W.; Gao, F.; Zhao, W.; Fu, Q.; Xia, J. Root Growth and Architecture of Tamarix Chinensis in Response to the Groundwater Level in the Yellow River Delta. Mar. Pollut. Bull. 2022, 179, 113717. [Google Scholar] [CrossRef]
- Quijano-Guerta, C.; Kirk, G.J.D.; Portugal, A.M.; Bartolome, V.I.; Mclaren, G.C. Tolerance of rice germplasm to zinc deficiency. Field Crops Res. 2002, 76, 123–130. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Huang, J.J.; Liu, Z.M.; Wang, P.; Han, W.D. Study on Spatial Connection and Fractal Characteristics of Prop Roots of Natural Rhizophora Stylosa. For. Res. 2013, 26, 359–365. [Google Scholar]
- Savaldi-Goldstein, S.; Peto, C.; Chory, J. The epidermis both drives and restricts plant shoot growth. Nature 2007, 446, 199–202. [Google Scholar] [CrossRef]
- Gonzalez-Garcia, M.P.; Vilarrasa-Blasi, J.; Zhiponova, M.; Divol, F.; Mora-Garcia, S.; Russinova, E.; Cano-Delgado, A.I. Brassinosteroids control meristem size by promoting cell cycle progression in Arabidopsis roots. Development 2011, 138, 849–859. [Google Scholar] [CrossRef] [Green Version]
- Hacham, Y.; Holland, N.; Butterfield, C.; Ubeda-Tomas, S.; Bennett, M.J.; Chory, J.; Savaldi-Goldstein, S. Brassinosteroid perception in the epidermis controls root meristem size. Development 2011, 138, 839–848. [Google Scholar] [CrossRef] [Green Version]
- Wolfram, H.; Angela, S.; Eleonore, H. Abscisic acid in the xylem: Where does it come from, where does it go to. J. Exp. Bot. 2002, 53, 27–32. [Google Scholar]
- Li, Y.; Li, Z.-S.; Miao, L.-H.; Wang, Y.-F.; Tong, Z.-Y.; He, F.; Wan, L.-Q.; Li, X.-L. Effects of Drought Stress at Different Growth Stages on Growth and Root ABA Content of Alfalfa. Acta Agrestia Sin. 2017, 25, 1245–1250. [Google Scholar]
- Leung, J.; Giraudat, J. Abscisic acid signal transduction. Annu. Rev. Plant Biol. 1998, 49, 199–222. [Google Scholar] [CrossRef] [Green Version]
- Smet, I.D.; Signora, L.; Beeckman, T.; Inzé, D.; Foyer, C.H.; Zhang, H. An abscisic acid-sensitive checkpoint in lateral root development of Arabidopsis. Plant J. 2010, 33, 543–555. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Li, C.; Wu, Z.; Jia, Y.; Wang, H.; Sun, S.; Mao, C.; Wang, X. Abscisic Acid Regulates Auxin Homeostasis in Rice Root Tips to Promote Root Hair Elongation. Front. Plant Sci. 2017, 8, 1121. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Inukai, Y.; Yamauchi, A. Root Development and Nutrient Uptake. Crit. Rev. Plant Sci. 2006, 25, 279–301. [Google Scholar] [CrossRef]
- Tatsumi, J. Fractal Geometry in Root Systems: Quantitative Evaluation of Distribution Pattern. Jpn. J. Crop Sci. 1995, 64, 50–57. [Google Scholar] [CrossRef]
- Glimskr, A. Estimates of root system topology of five plant species grown at steady-state nutrition. Plant Soil 2000, 227, 249–256. [Google Scholar] [CrossRef]
- Agurla, S.; Gahir, S.; Munemasa, S.; Murata, Y.; Raghavendra, A.S. Mechanism of Stomatal Closure in Plants Exposed to Drought and Cold Stress: Adaptation Mechanisms and Their Applications. In Survival Strategies in Extreme Cold and Desiccation; Springer: Singapore, 2018; Volume 1081. [Google Scholar]
- Chae, L.; Woonhee, B.; Jangho, J.; Jung-Hyun, K.; Sung, L. Function of ABA in Stomatal Defense against Biotic and Drought Stresses. Int. J. Mol. Sci. 2015, 16, 15251–15270. [Google Scholar]
- Takuya, Y.; Junro, M.; Kazuko, Y.S. Omics Approaches Toward Defining the Comprehensive Abscisic Acid Signaling Network in Plants. Plant Cell Physiol. 2015, 56, 1043–1052. [Google Scholar]
- Lodge, G.M. Management practices and other factors contributing to the decline in persistence of grazed lucerne in temperate Australia: A review. Aust. J. Exp. Agric. 1991, 31, 713–724. [Google Scholar] [CrossRef]
- Schuppler, U.; He, P.H.; Peter, C.L.J.; Munns, R. Effect of water stress on cell division and cell-division-cycle 2-like cell-cycle kinase activity in wheat leaves. Plant Physiol. 1998, 117, 667–678. [Google Scholar] [CrossRef] [Green Version]
- Kavar, T.; Maras, M.; Kidrič, M.; Šuštar-Vozlič, J.; Meglič, V. Identification of genes involved in the response of leaves of Phaseolus vulgaris to drought stress. Mol. Breed. 2008, 21, 159–172. [Google Scholar] [CrossRef]
- Malamy, J.E. Intrinsic and environmental response pathways that regulate root system architecture. Plant Cell Environ. 2005, 28, 67–77. [Google Scholar] [CrossRef]
- Duvick, D.N.; Smith, J.S.C.; Cooper, M. Long-Term Selection in a Commercial Hybrid Maize Breeding Program; John Wiley & Sons, Ltd.: Oxford, UK, 2004. [Google Scholar]
- Decuseara, C.; Nedelea, G. The effect of genotype and water stress on root growth in some alfalfa varieties. J. Hortic. For. Biotechnol. 2012, 16, 149–152. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Nie, Z.; Sun, J.; Li, X.; Yang, G. The Physiological Role of Abscisic Acid in Regulating Root System Architecture of Alfalfa in Its Adaptation to Water Deficit. Agronomy 2022, 12, 1882. https://doi.org/10.3390/agronomy12081882
Li S, Nie Z, Sun J, Li X, Yang G. The Physiological Role of Abscisic Acid in Regulating Root System Architecture of Alfalfa in Its Adaptation to Water Deficit. Agronomy. 2022; 12(8):1882. https://doi.org/10.3390/agronomy12081882
Chicago/Turabian StyleLi, Shuo, Zhongnan Nie, Juan Sun, Xianglin Li, and Guofeng Yang. 2022. "The Physiological Role of Abscisic Acid in Regulating Root System Architecture of Alfalfa in Its Adaptation to Water Deficit" Agronomy 12, no. 8: 1882. https://doi.org/10.3390/agronomy12081882