
Iron Biofortification of Greenhouse Soilless Lettuce: An Effective Agronomic Tool to Improve the Dietary Mineral Intake

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plant Material
2.2. Treatments
2.3. Lettuce Measurements
2.4. Biochemical Analyses
2.4.1. Leaf Chlorophylls and Carotenoids Concentration
2.4.2. Total Phenol and Flavonoid Concentration
2.4.3. Anthocyanin Concentration
2.4.4. Ascorbic Acid Concentration
2.4.5. Antioxidant Capacity: FRAP and TEAC Assays
2.4.6. Superoxide Anion
2.4.7. Proline
2.4.8. MDA
2.4.9. APX
2.4.10. GPX
2.4.11. CAT
2.5. Mineral Analyses
Forms of Nitrogen
2.6. Statistical Procedures
3. Results
3.1. Lettuce Main Traits and Bioactive Compounds Concentration
3.2. Oxidative Stress Indicators
3.3. Macronutrients and Micronutrients Content
3.4. Nitrogen Forms in Lettuce Leaves
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zoroddu, M.A.; Aaseth, J.; Crisponi, G.; Medici, S.; Peana, M.; Nurchi, V.M. The essential metals for humans: A brief overview. J. Inorg. Biochem. 2019, 195, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Haschka, D.; Hoffmann, A.; Weiss, G. Iron in immune cell function and host defense. Semin. Cell Dev. Biol. 2021, 115, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Camaschella, C. New insights into iron deficiency and iron deficiency anemia. Blood Rev. 2017, 31, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Wishart, K. Increased Micronutrient Requirements during Physiologically Demanding Situations: Review of the Current Evidence. Vitam. Miner. 2017, 6, 166. [Google Scholar] [CrossRef]
- de Valença, A.W.; Bake, A.; Brouwer, I.D.; Giller, K.E. Agronomic biofortification of crops to fight hidden hunger in sub-Saharan Africa. Glob. Food Secur. 2017, 12, 8–14. [Google Scholar] [CrossRef]
- Lillford, P.; Hermansson, A.-M. Global missions and the critical needs of food science and technology. Trends Food Sci. Technol. 2021, 111, 800–811. [Google Scholar] [CrossRef]
- Biesalski, H.K. Sustainable Micronutrients in Europe: Is There Cause for Concern? In Sustainable Nutrition in a Changing World; Springer International Publishing: Cham, Switzerland, 2017; pp. 143–166. ISBN 9783319559421. [Google Scholar]
- Poelman, M.; Strak, M.; Schmitz, O.; Hoek, G.; Karssenberg, D.; Helbich, M.; Ntarladima, A.-M.; Bots, M.; Brunekreef, B.; Grobbee, R.; et al. Relations between the residential fast-food environment and the individual risk of cardiovascular diseases in The Netherlands: A nationwide follow-up study. Eur. J. Prev. Cardiol. 2018, 25, 1397–1405. [Google Scholar] [CrossRef] [Green Version]
- Pasricha, S.R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron deficiency. Lancet 2021, 397, 233–248. [Google Scholar] [CrossRef]
- Cappellini, M.D.; Musallam, K.M.; Taher, A.T. Iron deficiency anaemia revisited. J. Intern. Med. 2020, 287, 153–170. [Google Scholar] [CrossRef]
- Olson, R.; Gavin-Smith, B.; Ferraboschi, C.; Kraemer, K. Food Fortification: The Advantages, Disadvantages and Lessons from Sight and Life Programs. Nutrients 2021, 13, 1118. [Google Scholar] [CrossRef]
- Mazzoni, L.; Ariza Fernández, M.T.; Capocasa, F. Potential health benefits of fruits and vegetables. Appl. Sci. 2021, 11, 8951. [Google Scholar] [CrossRef]
- Mauro, R.P.; Agnello, M.; Rizzo, V.; Graziani, G.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Recovery of eggplant field waste as a source of phytochemicals. Sci. Hortic. 2020, 261, 109023. [Google Scholar] [CrossRef]
- Sabatino, L.; La Bella, S.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C.; Consentino, B.B.; Rouphael, Y. Selenium biofortification and grafting modulate plant performance and functional features of cherry tomato grown in a soilless system. Sci. Hortic. 2021, 285, 110095. [Google Scholar] [CrossRef]
- Ierna, A.; Pellegrino, A.; Mauro, R.P.; Leonardi, C. Micronutrient foliar fertilization for the biofortification of raw and minimally processed early potatoes. Agronomy 2020, 10, 1744. [Google Scholar] [CrossRef]
- Sabatino, L.; Di Gaudio, F.; Consentino, B.B.; Rouphael, Y.; El-Nakhel, C.; La Bella, S.; Vasto, S.; Mauro, R.P.; D’Anna, F.; Iapichino, G.; et al. Iodine biofortification counters micronutrient deficiency and improve functional quality of open field grown curly endive. Horticulture 2021, 7, 58. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C. Enhancing quality of fresh vegetables through salinity eustress and biofortification applications facilitated by soilless cultivation. Front. Plant Sci. 2018, 9, 1254. [Google Scholar] [CrossRef]
- Savvas, D.; Gruda, N. Application of soilless culture technologies in the modern greenhouse industry—A review. Eur. J. Hortic. Sci. 2018, 83, 280–293. [Google Scholar] [CrossRef]
- Sambo, P.; Nicoletto, C.; Giro, A.; Pii, Y.; Valentinuzzi, F.; Mimmo, T.; Lugli, P.; Orzes, G.; Mazzetto, F.; Astolfi, S.; et al. Hydroponic Solutions for Soilless Production Systems: Issues and Opportunities in a Smart Agriculture Perspective. Front. Plant Sci. 2019, 10, 923. [Google Scholar] [CrossRef]
- Buturi, C.V.; Mauro, R.P.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Mineral biofortification of vegetables as a tool to improve human diet. Foods 2021, 10, 223. [Google Scholar] [CrossRef]
- Jones, J.D. Iron Availability and Management Considerations: A 4R Approach. Crops Soils 2020, 53, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Nozoye, T.; Nishizawa, N.K. Iron transport and its regulation in plants. Free Radic. Biol. Med. 2019, 133, 11–20. [Google Scholar] [CrossRef]
- Martens, D.C.; Westermann, D.T. Fertilizer Applications for Correcting Micronutrient Deficiencies. In Micronutrients in Agriculture; Soil Science Society of America: Madison, WI, USA, 2018; pp. 549–592. [Google Scholar]
- Carrasco-Gil, S.; Rios, J.J.; Álvarez-Fernández, A.; Abadía, A.; García-Mina, J.M.; Abadía, J. Effects of individual and combined metal foliar fertilisers on iron- and manganese-deficient Solanum lycopersicum plants. Plant Soil 2016, 402, 27–45. [Google Scholar] [CrossRef] [Green Version]
- Zahra, N.; Hafeez, M.B.; Shaukat, K.; Wahid, A.; Hasanuzzaman, M. Fe toxicity in plants: Impacts and remediation. Physiol. Plant. 2021, 173, 201–222. [Google Scholar] [CrossRef]
- de Souza, A.S.N.; de Oliveira Schmidt, H.; Pagno, C.; Rodrigues, E.; da Silva, M.A.S.; Flôres, S.H.; de Oliveira Rios, A. Influence of cultivar and season on carotenoids and phenolic compounds from red lettuce influence of cultivar and season on lettuce. Int. Food Res. J. 2022, 155, 111110. [Google Scholar] [CrossRef]
- Giordano, M.; El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Iron biofortification of red and green pigmented lettuce in closed soilless cultivation impacts crop performance and modulates mineral and bioactive composition. Agronomy 2019, 9, 290. [Google Scholar] [CrossRef] [Green Version]
- Shatilov, M.V.; Razin, A.F.; Ivanova, M.I. Analysis of the world lettuce market. IOP Conf. Ser. Earth Environ. Sci. 2019, 395, 012053. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Arcila, M.; Cartagena-Valenzuela, J.R.; Franco, G.; Correa-Londoño, G.A.; Quintero-Vásquez, L.M.; Gaviria-Montoya, C.A. Changes in the physico-chemical properties of four lettuce (Lactuca sativa L.) varieties during storage. Corpoica Cienc. Tecnol. Agropecu. 2017, 18, 257–273. [Google Scholar] [CrossRef] [Green Version]
- Mauro, R.P.; Agnello, M.; Distefano, M.; Sabatino, L.; Primo, A.S.B.; Leonardi, C.; Giuffrida, F. Chlorophyll fluorescence, photosynthesis and growth of tomato plants as affected by long-term oxygen root zone deprivation and grafting. Agronomy 2020, 10, 137. [Google Scholar] [CrossRef] [Green Version]
- Giuffrida, F.; Agnello, M.; Mauro, R.P.; Ferrante, A.; Leonardi, C. Cultivation under salt stress conditions influences postharvest quality and glucosinolates content of fresh-cut cauliflower. Sci. Hortic. 2018, 236, 166–174. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Chlorophyll and Carotenoid Determination. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Rivero, R.M.; Ruiz, J.M.; García, P.C.; López-Lefebre, L.R.; Sánchez, E.; Romero, L. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, 1–13. [Google Scholar] [CrossRef]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of Paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef]
- Kubiś, J. Exogenous spermidine differentially alters activities of some scavenging system enzymes, H2O2 and superoxide radical levels in water-stressed cucumber leaves. J. Plant Physiol. 2008, 165, 397–406. [Google Scholar] [CrossRef]
- Bieleski, R.L.; Turner, N.A. Separation and estimation of amino acids in crude plant extracts by thin-layer electrophoresis and chromatography. Anal. Biochem. 1966, 17, 278–293. [Google Scholar] [CrossRef]
- Fu, J.; Huang, B. Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environ. Exp. Bot. 2001, 45, 105–114. [Google Scholar] [CrossRef]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B- and ozone-induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Elia, A.C.; Galarini, R.; Taticchi, M.I.; Dörr, A.J.M.; Mantilacci, L. Antioxidant responses and bioaccumulation in Ictalurus melas under mercury exposure. Ecotoxicol. Environ. Saf. 2003, 55, 162–167. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Martín Peinado, F.J.; Romero-Freire, A.; García Fernández, I.; Sierra Aragón, M.; Ortiz-Bernad, I.; Simón Torres, M. Long-term contamination in a recovered area affected by a mining spill. Sci. Total Environ. 2015, 514, 219–223. [Google Scholar] [CrossRef]
- Krom, M.D. Spectrophotometric determination of ammonia: A study of a modified berthelot reaction using salicylate and dichloroisocyanurate. Analyst 1980, 105, 305–316. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Haroon, M.H.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant. Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Cecílio Filho, A.B.; Mendoza Cortez, J.W.; de Sordi, D.; Urrestarazu, M. Common chicory performance as influenced by iron concentration in the nutrient solution. J. Plant Nutr. 2015, 38, 1489–1494. [Google Scholar] [CrossRef] [Green Version]
- Broschat, T.K.; Moore, K.K. Phytotoxicity of several iron fertilizers and their effects on Fe, Mn, Zn, Cu, and P content of African marigolds and zonal geraniums. HortScience 2004, 39, 595–598. [Google Scholar] [CrossRef] [Green Version]
- Casey Barickman, T.; Sublett, W.L.; Miles, C.; Crow, D.; Scheenstra, E. Lettuce biomass accumulation and phytonutrient concentrations are influenced by genotype, n application rate and location. Horticulturae 2018, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, E.; Timmons, M.B.; Mattson, N.S. Quality, yield, and biomass efficacy of several hydroponic lettuce (Lactuca sativa L.) cultivars in response to high pressure sodium lights or light emitting diodes for greenhouse supplemental lighting. Horticulturae 2020, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Serio, F.; Elia, A.; Santamaria, P.; Rodriguez, G.R. Lettuce growth, yield and nitrate content as affected by electrical conductivity of nutrient solution. Acta Hortic. 2001, 559, 563–568. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Vaštakaitė-Kairienė, V.; Sutulienė, R.; Rasiukevičiūtė, N.; Viršilė, A.; Miliauskienė, J.; Laužikė, K.; Valiuškaitė, A.; Dėnė, L.; Chrapačienė, S.; et al. Phenolic compounds content evaluation of lettuce grown under short-term preharvest daytime or nighttime supplemental LEDs. Plants 2022, 11, 1123. [Google Scholar] [CrossRef]
- Potapovich, A.I.; Kostyuk, V.A. Comparative study of antioxidant properties and cytoprotective activity of flavonoids. Biochemistry 2003, 68, 514–519. [Google Scholar] [CrossRef]
- Kejík, Z.; Kaplánek, R.; Masařík, M.; Babula, P.; Matkowski, A.; Filipenský, P.; Veselá, K.; Gburek, J.; Sýkora, D.; Martásek, P.; et al. Iron complexes of flavonoids-antioxidant capacity and beyond. Int. J. Mol. Sci. 2021, 22, 646. [Google Scholar] [CrossRef]
- Yao, L.H.; Jiang, Y.M.; Shi, J.; Tomás-Barberán, F.A.; Datta, N.; Singanusong, R.; Chen, S.S. Flavonoids in food and their health benefits. Plant Foods Hum. Nutr. 2004, 59, 113–122. [Google Scholar] [CrossRef]
- Assefa, A.D.; Hur, O.-S.; Hahn, B.-S.; Kim, B.; Ro, N.-Y.; Rhee, J.-H. Nutritional metabolites of red pigmented lettuce (Lactuca sativa) germplasm and correlations with selected phenotypic characters. Foods 2021, 10, 2504. [Google Scholar] [CrossRef]
- Sigurdson, G.T.; Robbins, R.J.; Collins, T.M.; Giusti, M.M. Spectral and colorimetric characteristics of metal chelates of acylated cyanidin derivatives. Food Chem. 2017, 222, 1088–1095. [Google Scholar] [CrossRef]
- Shen, Y.; Li, J.; Gu, R.; Yue, L.; Wang, H.; Zhan, X.; Xing, B. Carotenoid and superoxide dismutase are the most effective antioxidants participating in ROS scavenging in phenanthrene accumulated wheat leaf. Chemosphere 2018, 197, 513–525. [Google Scholar] [CrossRef]
- Ems, T.; Lucia, K.S.; Huecker, M.R. Biochemistry, Iron Absorption; StatPearls Publishing LLC: Treasure Island, FL, USA, 2022; pp. 1–3. [Google Scholar]
- Abbaspour, N.; Hurrell, R.; Kelishadi, R. Review on iron and its importance for human health. J. Res. Med. Sci. 2014, 19, 164–174. [Google Scholar]
- Przybysz, A.; Wrochna, M.; Małecka-Przybysz, M.; Gawrońska, H.; Gawroński, S.W. Vegetable sprouts enriched with iron: Effects on yield, ROS generation and antioxidative system. Sci. Hortic. 2016, 203, 110–117. [Google Scholar] [CrossRef]
- Wilson, J.X. The physiological role of dehydroascorbic acid. FEBS Lett. 2002, 527, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Jibril, S.A.; Hassan, S.A.; Ishak, C.F.; Megat Wahab, P.E. Cadmium toxicity affects phytochemicals and nutrient elements composition of lettuce (Lactuca sativa L.). Adv. Agric. 2017, 2017, 1236830. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Szerement, J.; Szatanik-Kloc, A.; Mokrzycki, J.; Mierzwa-Hersztek, M. Agronomic biofortification with se, zn, and fe: An effective strategy to enhance crop nutritional quality and stress defense—A review. Soil Sci. Plant Nutr. 2022, 22, 1129–1159. [Google Scholar] [CrossRef]
- Tavanti, T.R.; de Melo, A.A.R.; Moreira, L.D.K.; Sanchez, D.E.J.; dos Santos Silva, R.; da Silva, R.M.; Dos Reis, A.R. Micronutrient fertilization enhances ROS scavenging system for alleviation of abiotic stresses in plants. Plant Physiol. Biochem. 2021, 160, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Sida-Arreola, J.P.; Sánchez-Chávez, E.; Ávila-Quezada, G.D.; Zamudio-Flores, P.B.; Acosta Muñíz, C.H. Iron biofortification and its impact on antioxidant system, yield and biomass in common bean. Plant Soil Environ. 2015, 61, 573–576. [Google Scholar] [CrossRef]
- Abioye, A.I.; Okuneye, T.A.; Odesanya, A.M.O.; Adisa, O.; Abioye, A.I.; Soipe, A.I.; Ismail, K.A.; Yang, J.W.F.; Fasehun, L.K.; Omotayo, M.O. Calcium intake and iron status in human studies: A systematic review and dose-response meta-analysis of randomized trials and crossover studies. J. Nutr. 2021, 11, 1084–1101. [Google Scholar] [CrossRef]
- Sularz, O.; Smoleń, S.; Koronowicz, A.; Kowalska, I.; Leszczyńska, T. Chemical composition of lettuce (Lactuca sativa L.) biofortified with iodine by KIO3, 5-Iodo-, and 3.5-diiodosalicylic acid in a hydroponic cultivation. Agronomy 2020, 10, 1022. [Google Scholar] [CrossRef]
- Borghesi, E.; Carmassi, G.; Uguccioni, M.C.; Vernieri, P.; Malorgio, F. Effects of calcium and salinity stress on quality of lettuce in soilless culture. J. Plant Nutr. 2013, 36, 677–690. [Google Scholar] [CrossRef]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 500–651. ISBN 9780123849052. [Google Scholar]
- Pretorius, B.; Schönfeldt, H.C.; Hall, N. Total and haem iron content lean meat cuts and the contribution to the diet. Food Chem. 2016, 193, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Jokinen, K.; Salovaara, A.K.; Wasonga, D.O.; Edelmann, M.; Simpura, I.; Mäkelä, P.S.A. Root-applied glycinebetaine decreases nitrate accumulation and improves quality in hydroponically grown lettuce. Food Chem. 2022, 366, 130–558. [Google Scholar] [CrossRef]
- Commission Regulation (EU) No 1258/2011 of 2 December 2011 amending Regulation (EC) No 1881/2006 as regards maximum levels for nitrates in foodstuffs. Off. J. Eur. Union 2011, 320, 15–17.
- Barrameda-Medina, Y.; Lentini, M.; Esposito, S.; Ruiz, J.M.; Blasco, B. Zn-biofortification enhanced nitrogen metabolism and photorespiration process in green leafy vegetable Lactuca sativa L. J. Sci. Food Agric. 2017, 97, 1828–1836. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Plant Biomass (g DW plant−1) | Dry Matter (%) | Total Chls (mg g−1 FW) | Chl a/b Ratio | Total Phenols (μg g−1 FW) | Anthocyanins (mg g−1 FW) | Flavonoids (μg g−1 FW) | Carotenoids (µg g−1 FW) | AsA (μg g−1 FW) | DHA (μg g−1 FW) | FRAP (μM FeSO4 g−1 FW) | TEAC (mg trolox g−1 FW) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fe concentration | |||||||||||||
| Fe0 | 20.0 a | 4.47 b | 2.33 c | 1.53 c | 535 c | 1.50 c | 542 c | 153 c | 100 c | 61.9 c | 6.19 c | 0.637 c | |
| Fe1 | 16.5 b | 5.03 a | 2.64 b | 1.70 b | 781 b | 2.05 b | 901 b | 220 b | 126 b | 79.4 b | 9.17 b | 0.881 b | |
| Fe2 | 16.2 b | 5.17 a | 3.26 a | 1.89 a | 926 a | 2.42 a | 1134 a | 304 a | 143 a | 95.9 a | 12.3 a | 1.173 a | |
| Cultivar | |||||||||||||
| ‘Nauplus’ | 14.6 b | 4.44 b | 2.67 b | 1.69 | 727 | 2.11 a | 881 | 203 b | 116 | 70.0 b | 8.56 b | 0.840 b | |
| ‘Romana’ | 20.5 a | 5.34 a | 2.82 a | 1.72 | 767 | 1.87 b | 838 | 249 a | 130 | 88.2 a | 9.86 a | 0.954 a | |
| Fe × Cv | |||||||||||||
| Fe0 ‘Nauplus’ | 15.1 | 4.09 | 2.21 | 1.52 | 559 | 1.77 c | 686 d | 134 | 97 d | 51.2 c | 6.07 | 0.65 d | |
| Fe1 ‘Nauplus’ | 15 | 4.36 | 2.85 | 1.76 | 766 | 2.11 b | 924 bc | 234 | 131 b | 83.2 b | 9.35 | 0.84 c | |
| Fe2 ‘Nauplus’ | 13.8 | 4.89 | 2.96 | 1.80 | 856 | 2.44 a | 1032 b | 242 | 121 bc | 75.5 b | 10.28 | 1.03 b | |
| Fe0 ‘Romana’ | 24.8 | 4.85 | 2.46 | 1.54 | 511 | 1.22 d | 399 e | 173 | 104 cd | 72.7 b | 9.00 | 0.62 d | |
| Fe1 ‘Romana’ | 18.0 | 5.71 | 2.43 | 1.65 | 796 | 2.00 bc | 879 c | 207 | 120 bc | 75.7 b | 10.28 | 0.92 bc | |
| Fe2 ‘Romana’ | 18.7 | 5.45 | 3.56 | 1.97 | 995 | 2.40 a | 1236 a | 367 | 166 a | 116.4 a | 14.28 | 1.32 a | |
| Significance | |||||||||||||
| Fe concentration | * | * | *** | *** | *** | ** | *** | *** | ** | ** | *** | *** | |
| Cultivar | ** | ** | ** | NS | NS | ** | NS | *** | NS | ** | ** | * | |
| Fe × Cv | NS | NS | NS | NS | NS | * | *** | NS | * | ** | NS | * | |
| O2− (μg g−1 FW) | Proline (μg g−1 FW) | MDA (µM g−1 FW) | APX (Δ Abs mg protein−1 min−1 FW) | GPX (Δ Abs mg protein−1 min−1 FW) | CAT (Δ Abs mg protein−1 min−1 FW) | ||
|---|---|---|---|---|---|---|---|
| Fe concentration | |||||||
| Fe0 | 6.91 c | 15.3 b | 2.92 b | 0.055 b | 0.171 a | 0.011 a | |
| Fe1 | 8.09 b | 18.9 ab | 4.16 a | 0.084 a | 0.149 b | 0.010 b | |
| Fe2 | 9.21 a | 24.7 a | 4.28 a | 0.086 a | 0.155 b | 0.009 c | |
| Cultivar | |||||||
| ‘Nauplus’ | 7.86 | 15.6 b | 3.61 | 0.084 a | 0.153 | 0.011 a | |
| ‘Romana’ | 8.28 | 23.7 a | 3.97 | 0.066 b | 0.163 | 0.008 b | |
| Fe × Cv | |||||||
| Fe0 ‘Nauplus’ | 6.61 d | 15.7 | 2.81 | 0.064 | 0.163 | 0.013 | |
| Fe1 ‘Nauplus’ | 8.61 b | 15.2 | 4.27 | 0.097 | 0.146 | 0.011 | |
| Fe2 ‘Nauplus’ | 8.36 bc | 16.0 | 3.74 | 0.090 | 0.150 | 0.010 | |
| Fe0 ‘Romana’ | 7.21 cd | 14.8 | 3.03 | 0.046 | 0.179 | 0.009 | |
| Fe1 ‘Romana’ | 7.58 bcd | 22.7 | 4.05 | 0.071 | 0.151 | 0.008 | |
| Fe2 ‘Romana’ | 10.06 a | 33.5 | 4.82 | 0.082 | 0.160 | 0.007 | |
| Significance | |||||||
| Fe concentration | ** | * | ** | * | * | ** | |
| Cultivar | NS | * | NS | * | NS | *** | |
| Fe × Cv | * | NS | NS | NS | NS | NS | |
| Macronutrients | Micronutrients | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
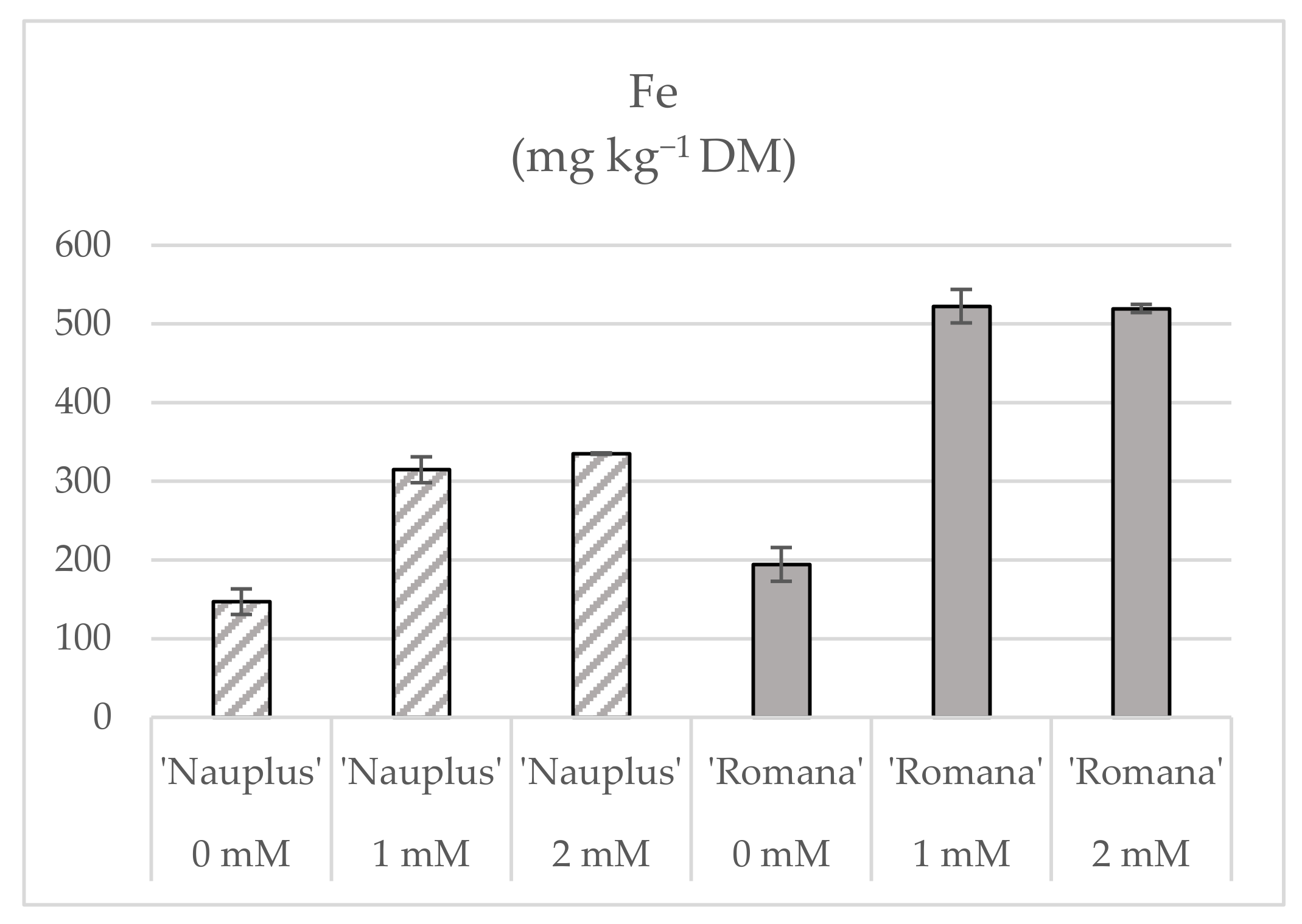
| N | P | K | Ca | Mg | S | Fe | Mn | Zn | B | Cu | ||
| Fe concentration | ||||||||||||
| Fe0 | 4.41 b | 3.88 c | 2.90 c | 0.330 c | 0.143 c | 0.107 c | 7.7 b | 3.49 c | 3.16 c | 1.21 c | 0.689 | |
| Fe1 | 4.68 ab | 4.60 b | 3.07 b | 0.385 a | 0.231 a | 0.135 b | 21.8 a | 6.28 a | 5.74 b | 1.93 a | 0.607 | |
| Fe2 | 4.99 a | 5.06 a | 3.75 a | 0.371 b | 0.215 b | 0.155 a | 22.4 a | 5.64 b | 6.04 a | 1.80 b | 0.695 | |
| Cultivar | ||||||||||||
| ‘Nauplus’ | 4.84 a | 3.96 b | 3.07 b | 0.347 b | 0.166 b | 0.123 b | 12.0 b | 4.33 b | 4.32 b | 1.51 b | 0.603 | |
| ‘Romana’ | 4.54 b | 5.06 a | 3.40 a | 0.377 a | 0.227 a | 0.141 a | 22.5 a | 5.94 a | 5.64 a | 1.78 a | 0.725 | |
| Fe × Cv | ||||||||||||
| Fe0 ‘Nauplus’ | 4.59 | 3.75 | 3.00 | 0.327 c | 0.132 | 0.109 | 6.0 d | 3.20 d | 3.00 d | 1.16 d | 0.602 | |
| Fe1 ‘Nauplus’ | 4.23 | 3.64 | 2.45 | 0.292 c | 0.162 | 0.117 | 13.7 b | 4.09 c | 4.25 c | 1.41 cd | 0.775 | |
| Fe2 ‘Nauplus’ | 5.70 | 4.51 | 3.77 | 0.421 b | 0.204 | 0.144 | 16.4 b | 5.69 b | 5.71 b | 1.96 b | 0.555 | |
| Fe0 ‘Romana’ | 4.22 | 4.02 | 2.80 | 0.332 c | 0.154 | 0.104 | 9.4 c | 3.78 cd | 3.33 cd | 1.25 d | 0.660 | |
| Fe1 ‘Romana’ | 5.12 | 5.56 | 3.69 | 0.478 a | 0.300 | 0.153 | 29.8 a | 8.46 a | 7.23 a | 2.45 a | 0.651 | |
| Fe2 ‘Romana’ | 4.28 | 5.62 | 3.72 | 0.321 c | 0.226 | 0.166 | 28.3 a | 5.59 b | 6.36 ab | 1.64 c | 0.740 | |
| Significance | ||||||||||||
| Fe concentration | * | *** | *** | *** | *** | ** | *** | *** | *** | *** | NS | |
| Cultivar | * | *** | ** | * | *** | ** | *** | *** | ** | ** | NS | |
| Fe × Cv | NS | NS | NS | *** | NS | NS | *** | *** | ** | *** | NS | |
| Organic N (mg g−1 DW) | NH4+ (mg g−1 DW) | NO3− (mg g−1 DW) | Total N (mg g−1 DW) | Mineral N (mg g−1 DW) | Assimilated N (mg g−1 DW) | ||
|---|---|---|---|---|---|---|---|
| Fe concentration | |||||||
| Fe0 | 33.4 | 6.28 c | 66.2 a | 99.6 | 72.5 a | 27.1 b | |
| Fe1 | 40.6 | 8.79 a | 52.8 b | 93.4 | 61.6 b | 31.8 a | |
| Fe2 | 40.7 | 7.62 b | 56.9 b | 97.6 | 64.5 ab | 33.1 a | |
| Cultivar | |||||||
| ‘Nauplus’ | 42.2 a | 7.48 | 66.4 a | 108.7 a | 73.9 a | 34.7 a | |
| ‘Romana’ | 34.3 b | 7.65 | 50.8 b | 85.1 b | 58.5 b | 26.7 b | |
| Fe × Cv | |||||||
| Fe0 ‘Nauplus’ | 37.1 | 7.00 | 75.2 | 112.3 | 82.2 | 30.1 | |
| Fe1 ‘Nauplus’ | 44.5 | 9.13 | 52.6 | 97.1 | 61.8 | 35.4 | |
| Fe2 ‘Nauplus’ | 45.2 | 6.31 | 71.4 | 116.6 | 77.7 | 38.9 | |
| Fe0 ‘Romana’ | 29.8 | 5.55 | 57.2 | 87.0 | 62.7 | 24.3 | |
| Fe1 ‘Romana’ | 36.8 | 8.46 | 52.9 | 89.7 | 61.4 | 28.3 | |
| Fe2 ‘Romana’ | 36.2 | 8.94 | 42.4 | 78.6 | 51.3 | 27.3 | |
| Significance | |||||||
| Fe concentration | NS | ** | * | NS | * | * | |
| Cultivar | ** | NS | *** | *** | *** | ** | |
| Fe × Cv | NS | NS | NS | NS | NS | NS | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buturi, C.V.; Sabatino, L.; Mauro, R.P.; Navarro-León, E.; Blasco, B.; Leonardi, C.; Giuffrida, F. Iron Biofortification of Greenhouse Soilless Lettuce: An Effective Agronomic Tool to Improve the Dietary Mineral Intake. Agronomy 2022, 12, 1793. https://doi.org/10.3390/agronomy12081793
Buturi CV, Sabatino L, Mauro RP, Navarro-León E, Blasco B, Leonardi C, Giuffrida F. Iron Biofortification of Greenhouse Soilless Lettuce: An Effective Agronomic Tool to Improve the Dietary Mineral Intake. Agronomy. 2022; 12(8):1793. https://doi.org/10.3390/agronomy12081793
Chicago/Turabian StyleButuri, Camila Vanessa, Leo Sabatino, Rosario Paolo Mauro, Eloy Navarro-León, Begoña Blasco, Cherubino Leonardi, and Francesco Giuffrida. 2022. "Iron Biofortification of Greenhouse Soilless Lettuce: An Effective Agronomic Tool to Improve the Dietary Mineral Intake" Agronomy 12, no. 8: 1793. https://doi.org/10.3390/agronomy12081793