
Genetic Mapping and Identification of the Gibberellin 3-Oxidase Gene GA3ox Leading to a GA-Deficient Dwarf Phenotype in Pumpkin (Cucurbita moschata D.)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
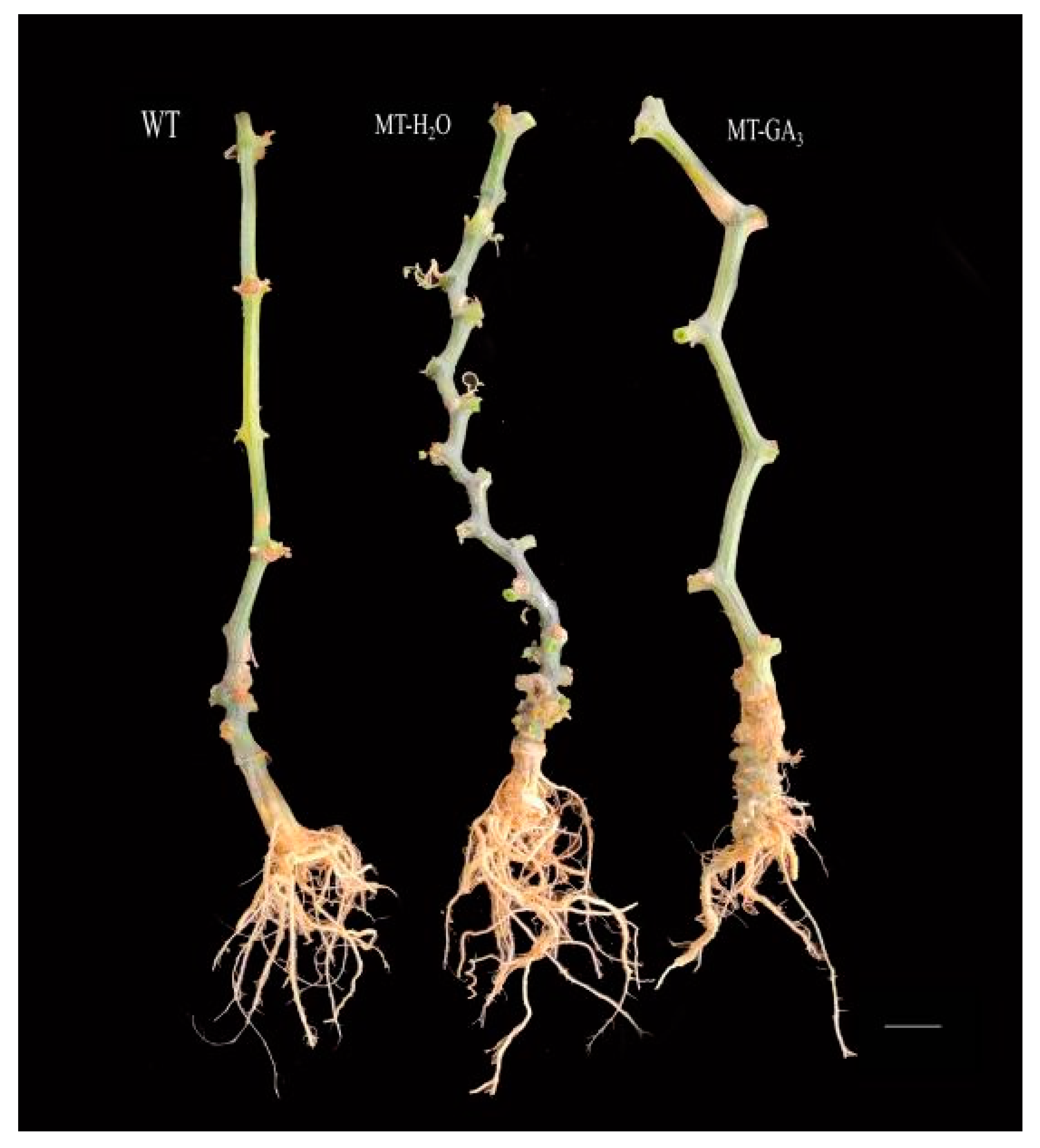
2.2. Plant Phenotyping and Morphological Characterization
2.3. Bulked Segregant Analysis (BSA)
2.4. Marker Development and Fine Mapping of the CmaSI1 Gene
2.5. Candidate Gene Analysis
2.6. RNA-Sequencing and Data Analysis
2.7. Treatment with Exogenous GA3
3. Results
3.1. Distinct Phenotypes of the Dwarf Mutant
3.2. Genetic Analysis of the Dwarf Mutant
3.3. Fine Mapping of the CmaSI1 Locus by BSA-Seq and Linkage Analyses
3.4. Identification of Candidate Genes for the CmaSI1 Gene
3.5. Analysis of the Expression of GAs Biosynthetic Pathways and Regulatory Genes
3.6. Si1 Is a GA Biosynthetic-Deficient Mutant
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiao, Z.; Fu, R.; Li, J.; Fan, Z.; Yin, H. Overexpression of the gibberellin 2-Oxidase gene from Camellia lipoensis induces dwarfism and smaller flowers in Nicotiana tabacum. Plant Mol. Biol. Rep. 2015, 34, 182–191. [Google Scholar] [CrossRef]
- Dong, W.; Wu, D.; Li, G.; Wu, D.; Wang, Z. Next-generation sequencing from bulked segregant analysis identifies a dwarfism gene in watermelon. Sci. Rep. 2018, 8, 2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S. Green revolution: A mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.; Yao, Q.; Zhang, X.; Xiao, X.; Wang, Y. The semi-dwarfing gene Rht-dp from dwarf Polish wheat (Triticum polonicum L.) is the “Green Revolution” gene Rht-B1b. BMC Genom. 2020, 22, 63. [Google Scholar] [CrossRef] [PubMed]
- Meland, M.; Frynes, O.; Maas, F. Performance of dwarfing and semi-dwarfing plum rootstocks on three european plum scion cultivars in a nordic climate. Acta Hortic. 2019, 1260, 181–186. [Google Scholar] [CrossRef]
- Velde, K.; Thomas, S.G.; Heyse, F.; Kaspar, R.; Rohde, A. N-terminal truncated RHT-1 proteins generated by translational reinitiation cause semi-dwarfing of wheat green revolution alleles. Mol. Plant 2021, 14, 9. [Google Scholar] [CrossRef] [PubMed]
- Bhuvaneswari, S.; Krishnan, S.; Ellur, R.; Vinod, K.; Singh, A. Discovery of a novel induced polymorphism in SD1 gene governing semi-dwarfism in rice and development of a functional marker for marker-assisted selection. Plants 2020, 9, 1198. [Google Scholar] [CrossRef]
- Monna, L.; Kitazawa, N.; Yoshino, R.; Suzuki, J.; Masuda, H.; Maehara, Y.; Tanji, M.; Sato, M.; Nasu, S.; Minobe, Y. Positional cloning of rice semi-dwarfing gene, sd-1: ‘Rice green revolution’ gene encodes a mutant enzyme involved in gibberellin synthesis. DNA Res. 2002, 9, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zheng, S.; Gui, J.; Fu, C.; Yu, H.; Song, D.; Shen, J.; Qin, P.; Liu, X.; Han, B.; et al. Shortened basal internodes encodes a gibberellin 2-oxidase and contributes to lodging resistance in rice. Mol. Plant 2018, 11, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Acua, T.; Spielmeyer, W.; Richards, R. Effect of ga sensitive rht18 and GA insensitive rht-d1b dwarfing genes on vegetative and reproductive growth in bread wheat. J. Exp. Bot. 2020, 72, 445–458. [Google Scholar] [CrossRef]
- Ford, B.; Foo, E.; Sharwood, R.; Karafifiatova, M.; Vrana, J.; MacMillan, C.; Nichols, D.; Steuernagel, B.; Uauy, C.; Dolezel, J.; et al. Rht18 semidwarfifism in wheat is due to increased GA2-oxidaseA9 expression and reduced GA content. Plant Physiol. 2018, 177, 168–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, A.; Fujioka, S.; Sunohara, H.; Kamiya, N.; Hong, Z.; Inukai, Y.; Miura, K.; Takatsuto, S.; Yoshida, S.; Ueguchi-Tanaka, M.; et al. The role of OsBRI1 and its homologous genes, OsBRL1 and OsBRL3, in rice. Plant Physiol. 2006, 140, 580–590. [Google Scholar] [CrossRef] [Green Version]
- Multani, D.; Briggs, S.; Chamberlin, M.; Blakeslee, J.; Murphy, A.; Johal, G. Loss of an MDR transporter in compact stalks of maize br2 and sorghum dw3 mutants. Science 2003, 302, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Tamiru, M.; Undan, J.; Takagi, H.; Abe, A.; Yoshida, K.; Undan, J.; Satoshi, N.; Aiko, U.; Hiromasa, S.; Hideo, M.; et al. A cytochrome P450, OsDSS1, is involved in growth and drought stress responses in rice (Oryza sativa L.). Plant Mol. Biol. 2015, 88, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zhang, M.; Sun, S.; Yang, S.; Yang, L. A single nucleotide deletion in an ABC transporter gene leads to a dwarf phenotype in watermelon. Front. Plant Sci. 2019, 10, 1399. [Google Scholar] [CrossRef]
- Hong, Z.; Ueguchi-Tanaka, M.; Matsuoka, M. Brassinosteroids and rice architecture. J. Pestic. Sci. 2004, 29, 184–188. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Zhou, S.; Ou, L.; Liu, F.; Yang, L.; Zheng, J.; Chen, W.; Zhang, Z.; Yang, S.; Ma, Y.; et al. A novel single-base mutation in CaBRI1 confers dwarf phenotype and brassinosteroid accumulation in pepper. Mol. Genet. Genom. 2020, 295, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Zhu, C.; Yang, L.; Zhao, W.; Ma, R.; Li, H.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X. A point mutation resulting in a 13 bp deletion in the coding sequence of Cldf leads to a GA-deficient dwarf phenotype in watermelon. Hortic. Res. 2019, 6, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.; Wang, S.; Zhong, Y.; Gao, D.; Cui, Q.; Chen, H. A truncated F-box protein confers the dwarfism in cucumber. J. Genet. Genom. 2016, 43, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Chao, W.; Wen, C.; Zhou, S.; Tao, W. Clavata1-type receptor-like kinase csclavata1 is a putative candidate gene for dwarf mutation in cucumber. Mol. Genet. Genom. 2018, 293, 1393–1405. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Yang, L.; Pathak, M.; Li, D.; He, X.; Weng, Y. Fine genetic mapping of cp: A recessive gene for compact (dwarf) plant architecture in cucumber, Cucumis sativus L. Theor. Appl. Genet. 2011, 123, 973. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Jiang, Z.; Zhang, E. Pumpkin function properties and application in food industry. Sci. Technol. Food Ind. 2000, 2, 62–64. [Google Scholar]
- Yadav, M.; Jain, S.; Tomar, R.; Prasad, G.; Yadav, H. Medicinal and biological potential of pumpkin: An updated review. Nutr. Res. Rev. 2010, 23, 184–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun-Xin, W.; Chang, Z. A construction of aflp analysis system for dwarf Cucurbita moschata research. Acta Agric. Boreali-Sin. 2011, 26, 124–127. [Google Scholar]
- Xiang, C.; Duan, Y.; Li, H.; Ma, W.; Huang, S.; Sui, X.; Zhang, Z.; Wang, C. A high-density EST-SSR-based genetic map and QTL analysis of dwarf trait in Cucurbita pepo L. Int. J. Mol. 2018, 19, 3140. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Huang, H.; Lin, Y.E.; Chen, Q.; Liang, Z.; Wu, T. Genetic and gene expression analysis of dm1, a dwarf mutant from Cucurbita maxima Duch. ex Lam, based on the AFLP method. Can. J. Plant Sci. 2014, 94, 293–302. [Google Scholar] [CrossRef]
- Zhang, G.; Ren, Y.; Sun, H. A high- density genetic map for anchoring genome sequences and identifying QTLs associated with dwarf vine in pumpkin (Cucurbita maxima Duch.). BMC Genom. 2015, 16, 1101–1110. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Cao, J. Differential gene expression of tropical pumpkin (Cucurbita moschata Duchesne) bush mutant during internode development. Sci. Hortic. 2008, 117, 219–224. [Google Scholar] [CrossRef]
- Min, Z.; Han, X.; Li, Y.; Hu, X.; Wang, D.; Sun, X. Construction and preliminary screening of EMS induced mutant library of pumpkin (Cucurbita moschata D.). J. Nucl. Agr. Sci. 2021, 35, 761–768. [Google Scholar]
- Hu, X.; Su, J.; Yuan, Z.; Li, Y.; Han, X. Breeding of xingshudaguomiben, a pumpkin cultivar with high yield and high quality. J. Chang. Veg. 2015, 12, 6–7. [Google Scholar]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Li, Z.; Liu, Z.; Guo, Y.; Qiu, L. Next-generation sequencing from bulked-segregant analysis accelerates the simultaneous identification of two qualitative genes in soybean. Front. Plant Sci. 2017, 8, 919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasheed, A.; He, Z.; Gao, F.; Zhai, S.; Jin, H.; Liu, J.; Guo, Q.; Zhang, Y.; Dreisigacker, S.; Xia, X.; et al. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat. Theor. Appl. Genet. 2016, 129, 1843–1860. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ding, W.; Wang, Y.; Qi, C.; Luo, Y.; Qu, S. Fine mapping identified the gibberellin 2-oxidase gene cpdw leading to a dwarf phenotype in squash (Cucurbita pepo L.). Plant Sci. 2021, 306, 110857. [Google Scholar] [CrossRef]
- Ling-Duo, B.; Si-Gui, J.; Jiang-Zhou, L. The effects of gibberellin (GA3) concentration on the growth, yield and quality of tobacco upper leaves. J. Anhui Agric. Sci. 2018, 46, 46–51. [Google Scholar]
- Hwang, J.; Song, W.; Hong, D.; Ko, D.; Yamaoka, Y.; Jang, S. Plant ABC transporters enable many unique aspects of a terrestrial plant’s lifestyle. Mol. Plant 2016, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lester, D.; Ross, J.; Davies, P.; Reid, J. Mendel’s stem length gene (Le) encodes a gibberellin 3β-hydroxylase. Plant Cell 1997, 9, 1435–1443. [Google Scholar]
- Wang, Y.; Zhao, J.; Lu, W.; Deng, D. Gibberellin in plant height control: Old player, new story. Plant Cell Rep. 2017, 36, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Cao, J.; Qin, Z.; Du, Y. Identification of a novel GA-related bush mutant in pumpkin (Cucurbita moschata Duchesne). Pak. J. Bot. 2015, 47, 1359–1366. [Google Scholar]
- Wang, D.; Wang, S.; Chao, J.; Wu, X.; Sun, Y.; Li, F.; Gao, X.; Liu, G.; Wang, Y. Morphological phenotyping and genetic analyses of a new chemical-mutagenized population of tobacco (Nicotiana tabacum L.). Planta 2017, 246, 149–163. [Google Scholar] [CrossRef]
- Hao, N.; Du, Y.; Li, H.; Wang, C.; Wang, C.; Gong, S. Csmyb36 is involved in the formation of yellow green peel in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2018, 131, 1659–1669. [Google Scholar] [CrossRef]
- Li, X.; Xiang, F.; Zhang, W.; Yan, J.; Li, X.; Zhong, M.; Yang, P.; Chen, C.; Liu, X.; Mao, D.; et al. Characterization and fine mapping of a new dwarf mutant in Brassica napus. BMC Plant Biol. 2021, 21, 117. [Google Scholar] [CrossRef]
- Ashikari, M.; Itoh, H.; Miyako, U. Gibberell in signal transduction in rice. Plant Growth Regul. 2003, 22, 141–151. [Google Scholar] [CrossRef]
- Sun, T. The molecular mechanism and evolution of the GA-GID1-DELLA signaling module in plants. Curr. Biol. 2011, 21, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Binenbaum, J.; Weinstain, R.; Shani, E.; Kamiya, Y. Gibberellin localization and transport in plants. Trends Plant Sci. 2018, 23, 410–421. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, S.; Sun, T.; Kawaide, H.; Kamiya, Y. The GA2 locus of Arabidopsis thaliana encodes ent-kaurene synthase of gibberellin biosynthesis. Plant Physiol. 1998, 116, 1271–1278. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Tang, D.; Liu, K.; Miao, C.; Zhuo, X.; Li, Y.; Tan, X.; Sun, M.; Luo, Q.; Cheng, Z. Characterization of a new semi-dominant dwarf allele of SLR1 and its potential application in hybrid rice breeding. J. Exp. Bot. 2018, 69, 4703–4713. [Google Scholar] [CrossRef]
- Mitchum, M.; Yamaguchi, S.; Hanada, A.; Kuwahara, A.; Yoshioka, Y.; Kato, T.; Tabata, S.; Kamiya, Y.; Sun, T. Distinct and overlapping roles of two gibberellin 3-oxidases in Arabidopsis development. Plant J. 2006, 45, 804–818. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, L.; Knop, N.; Theo, L. Stamen-derived bioactive gibberellin is essential for male flower development of Cucurbita maxima L. J. Exp. Bot. 2012, 63, 2681–2691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimenta, L.; Liebrandt, A.; Arnold, L.; Chmielewska, S.; Felsberger, A.; Freier, E.; Heuer, M.; Zur, D.; Lange, T. Functional characterization of gibberellin oxidases from cucumber, Cucumis sativus L. Phytochemistry 2013, 90, 62–69. [Google Scholar] [CrossRef]
- Chen, Y.; Hou, M.; Liu, L.; Wu, S.; Shen, Y.; Ishiyama, K.; Kobayashi, M.; McCarty, D.; Tan, B. The maize DWARF1 encodes a gibberellin 3-oxidase and is dual localized to the nucleus and cytosol. Plant Physiol. 2014, 166, 2028. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Generations | No. Normal Individuals | No. Dwarf Individuals | No. Total Individuals | Expected Ratio | χ2 |
|---|---|---|---|---|---|
| F1 (P1 × P2) | 40 | 0 | 40 | ||
| F1 (P2 × P1) | 48 | 0 | 48 | ||
| BC1P1 (F1 × N87) | 80 | 0 | 80 | ||
| BC1P2 (F1 × Si1) | 63 | 61 | 124 | 1:1 | 0.03 |
| F2 (1) | 188 | 56 | 244 | 3:1 | 0.55 |
| F2 (2) | 1488 | 512 | 2000 | 3:1 | 0.38 |
| Chromosome | Position | Gene_ID | WT | MT | Type |
|---|---|---|---|---|---|
| Cmo_Chr08 | 3688362 | CmoCh08G006070 | C | T | Intron |
| Cmo_Chr08 | 3747353 | CmoCh08G006150 | G | A | Intron |
| Cmo_Chr08 | 3779377 | CmoCh08G006170 | GGA:G | AGA:R | CDS_nonsyn |
| Cmo_Chr08 | 3815907 | CmoCh08G006250 | CTA:L | TTA:L | CDS_syn |
| Cmo_Chr08 | 3828051 | CmoCh08G006300 | G | A | Intron |
| Cmo_Chr08 | 4123114 | CmoCh08G006320 | G | A | Intron |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, Z.; Hu, X.; Han, X.; Li, Y.; Li, J.; Wang, D.; Sun, L.; Sun, X. Genetic Mapping and Identification of the Gibberellin 3-Oxidase Gene GA3ox Leading to a GA-Deficient Dwarf Phenotype in Pumpkin (Cucurbita moschata D.). Agronomy 2022, 12, 1779. https://doi.org/10.3390/agronomy12081779
Min Z, Hu X, Han X, Li Y, Li J, Wang D, Sun L, Sun X. Genetic Mapping and Identification of the Gibberellin 3-Oxidase Gene GA3ox Leading to a GA-Deficient Dwarf Phenotype in Pumpkin (Cucurbita moschata D.). Agronomy. 2022; 12(8):1779. https://doi.org/10.3390/agronomy12081779
Chicago/Turabian StyleMin, Ziyang, Xinjun Hu, Xiaoxia Han, Yongqi Li, Jiajia Li, Duanhua Wang, Longjun Sun, and Xiaowu Sun. 2022. "Genetic Mapping and Identification of the Gibberellin 3-Oxidase Gene GA3ox Leading to a GA-Deficient Dwarf Phenotype in Pumpkin (Cucurbita moschata D.)" Agronomy 12, no. 8: 1779. https://doi.org/10.3390/agronomy12081779