
Are Traditional Lima Bean (Phaseolus lunatus L.) Landraces Valuable to Cope with Climate Change? Effects of Drought on Growth and Biochemical Stress Markers

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Plant Growth and Water Stress Treatments
2.3. Plant Sampling and Growth Parameters
2.4. Photosynthetic Pigments
2.5. Ion Concentration Measurements
2.6. Osmolyte Quantification
2.7. Oxidative Stress Markers and Non-Enzymatic Antioxidants
2.8. Statistical Analysis
2.9. Electrochemical Experiments
3. Results
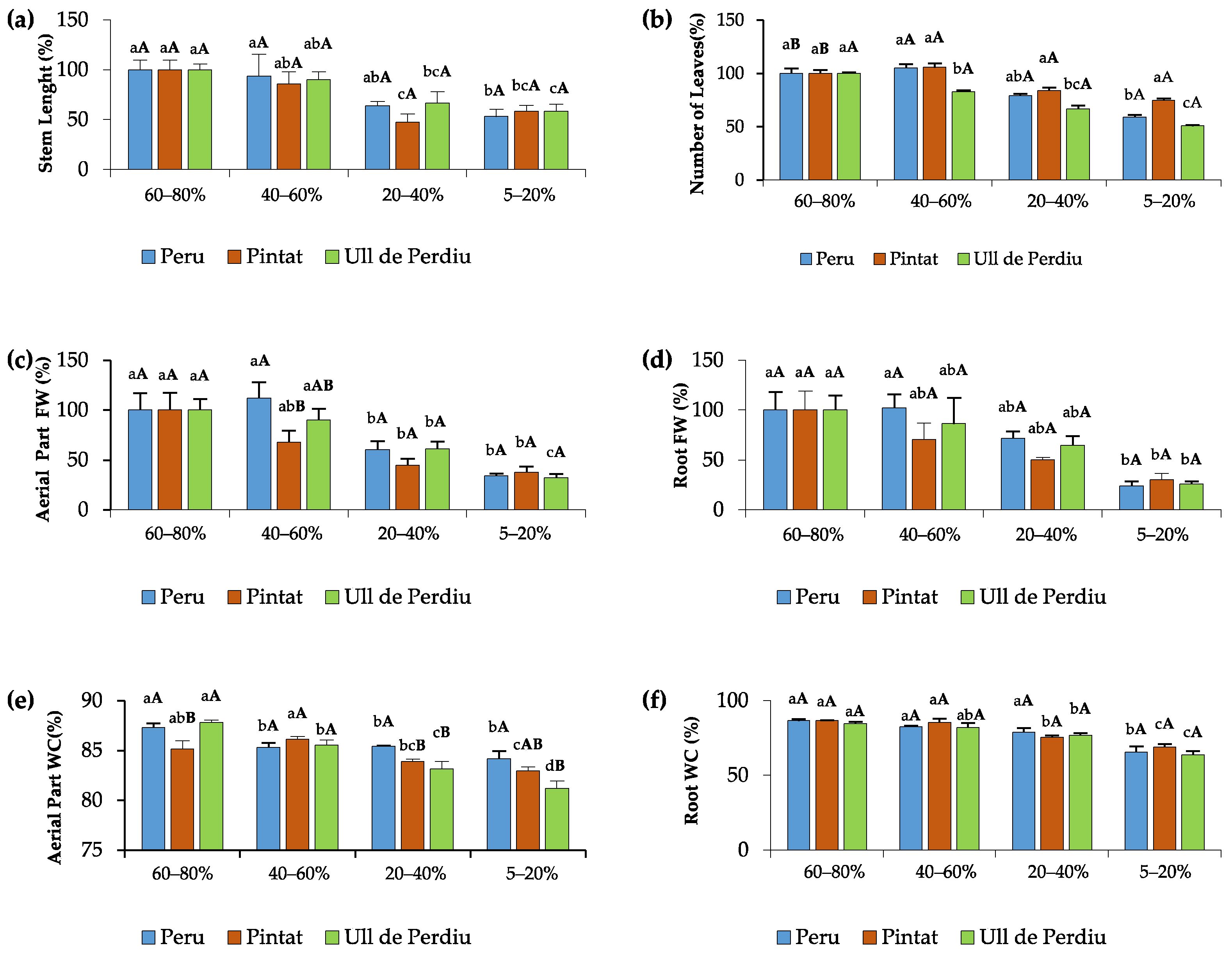
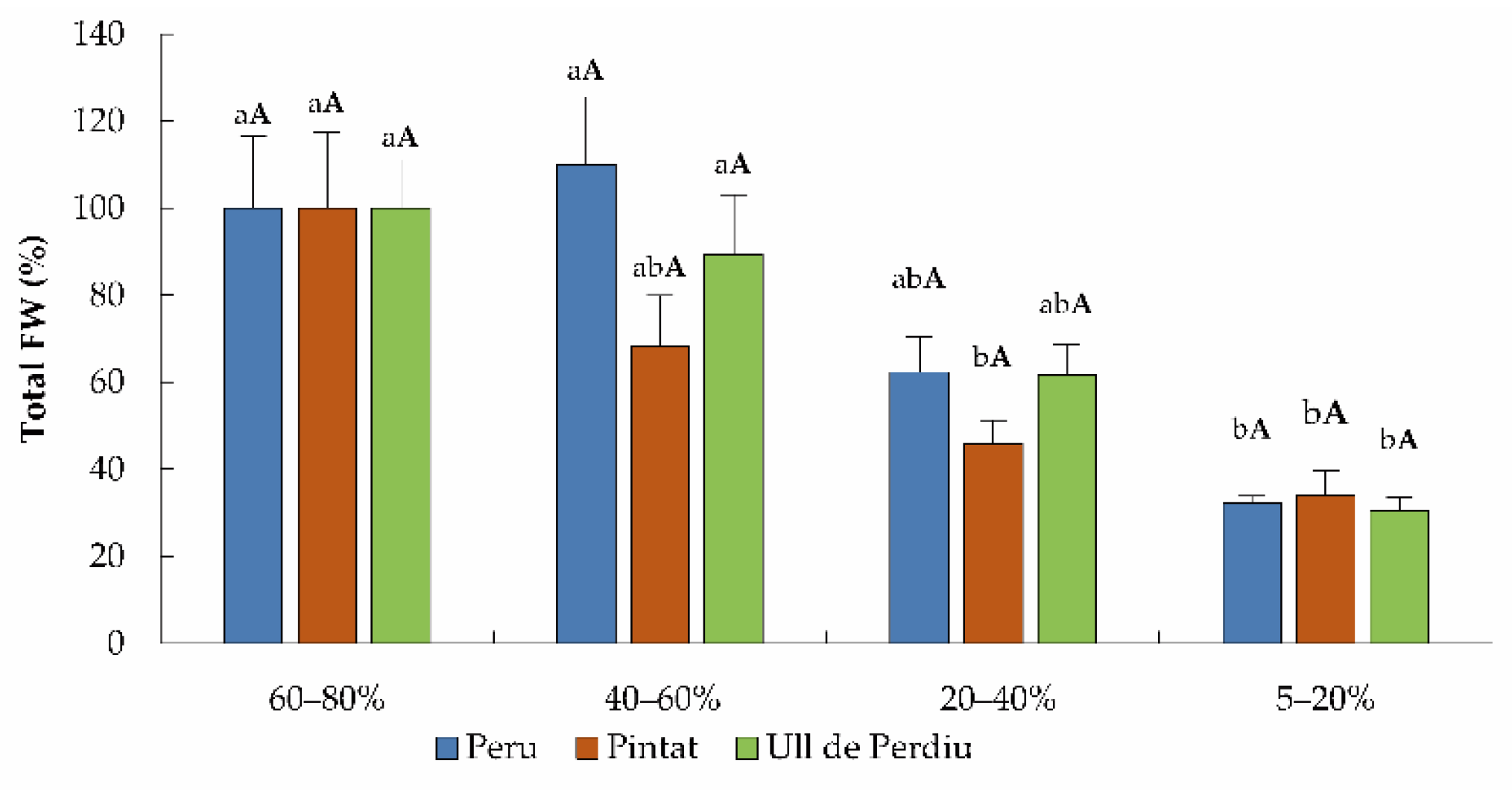
3.1. Plant Growth Analyses
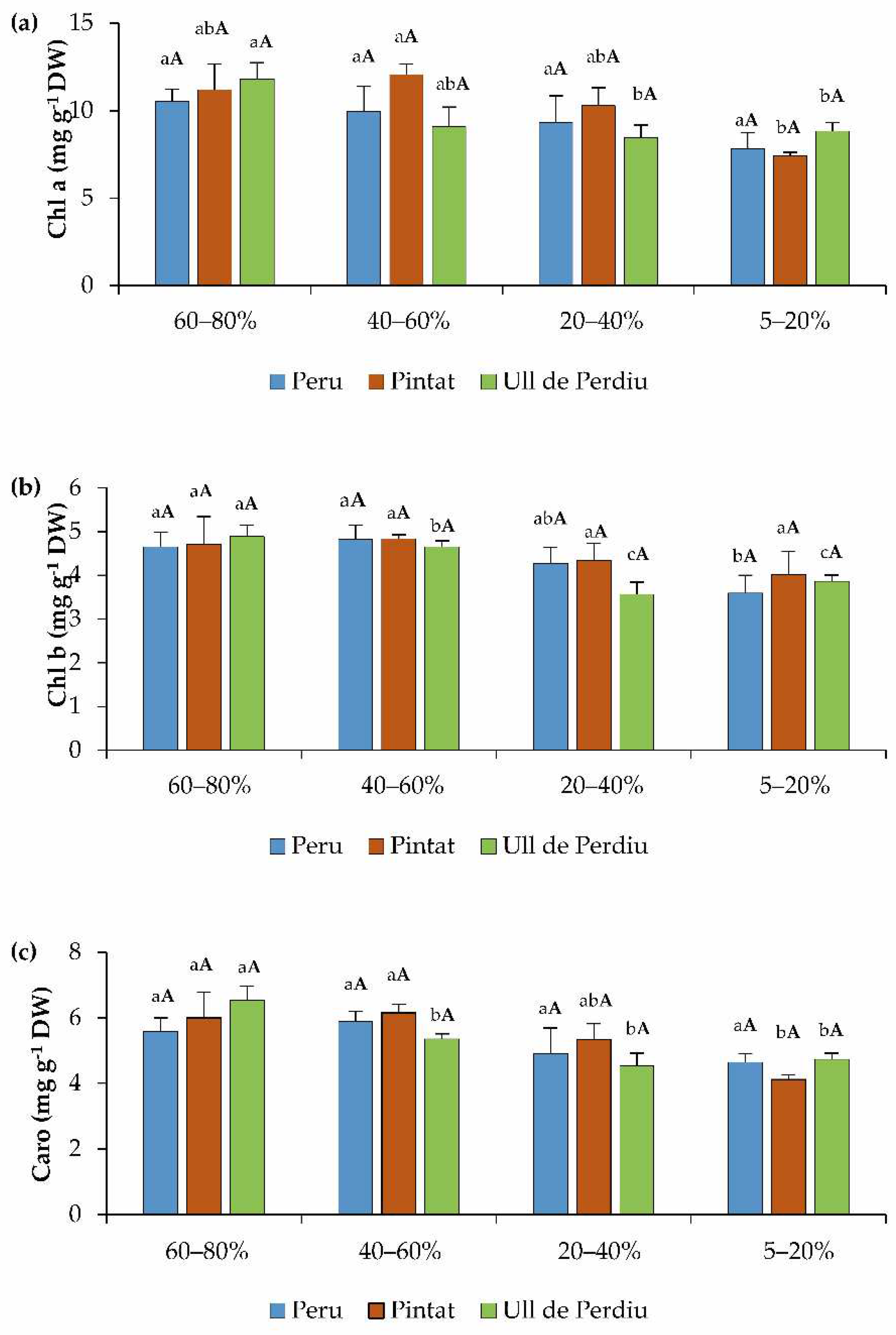
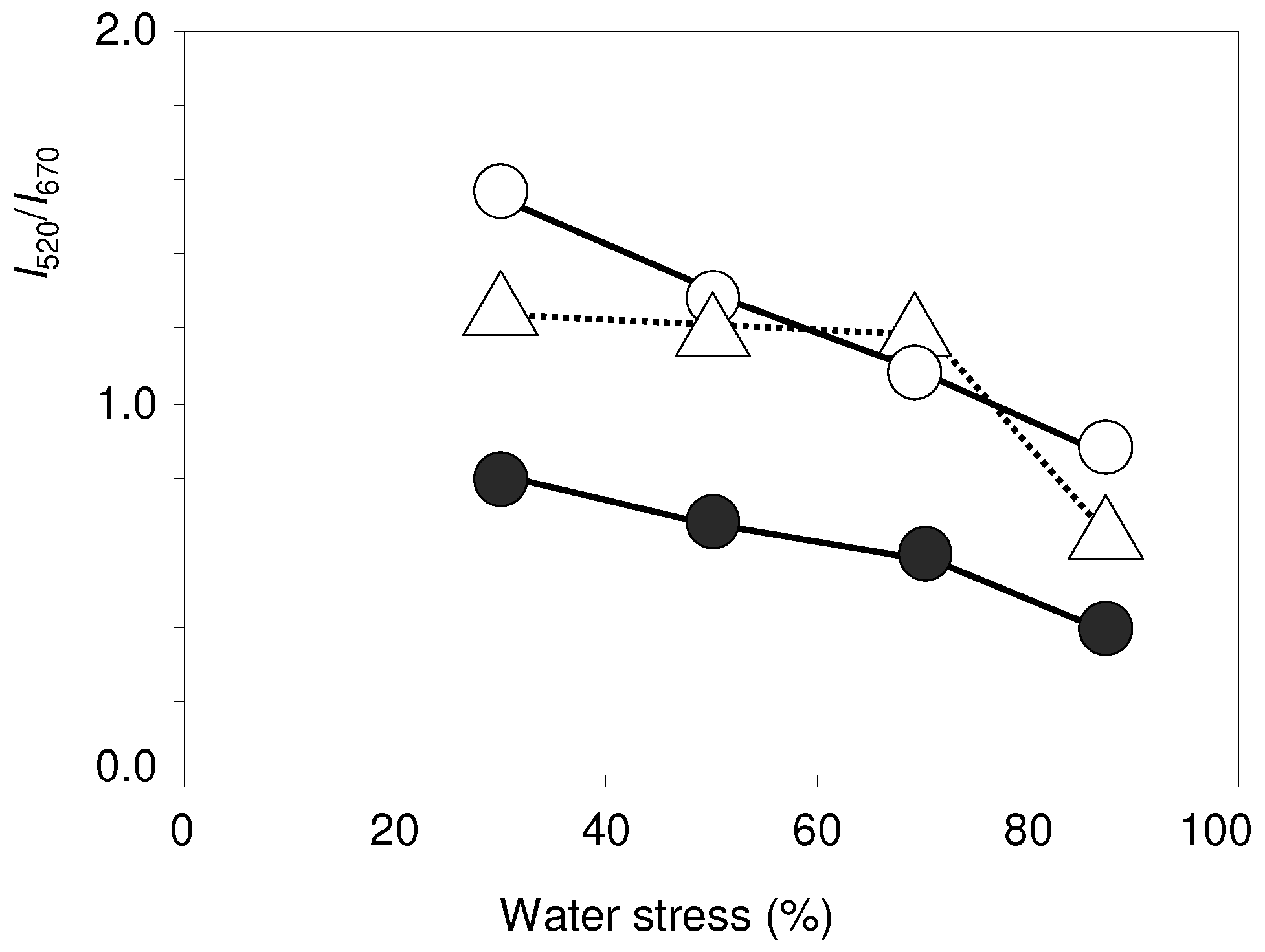
3.2. Photosynthetic Pigments
3.3. Ions Accumulation
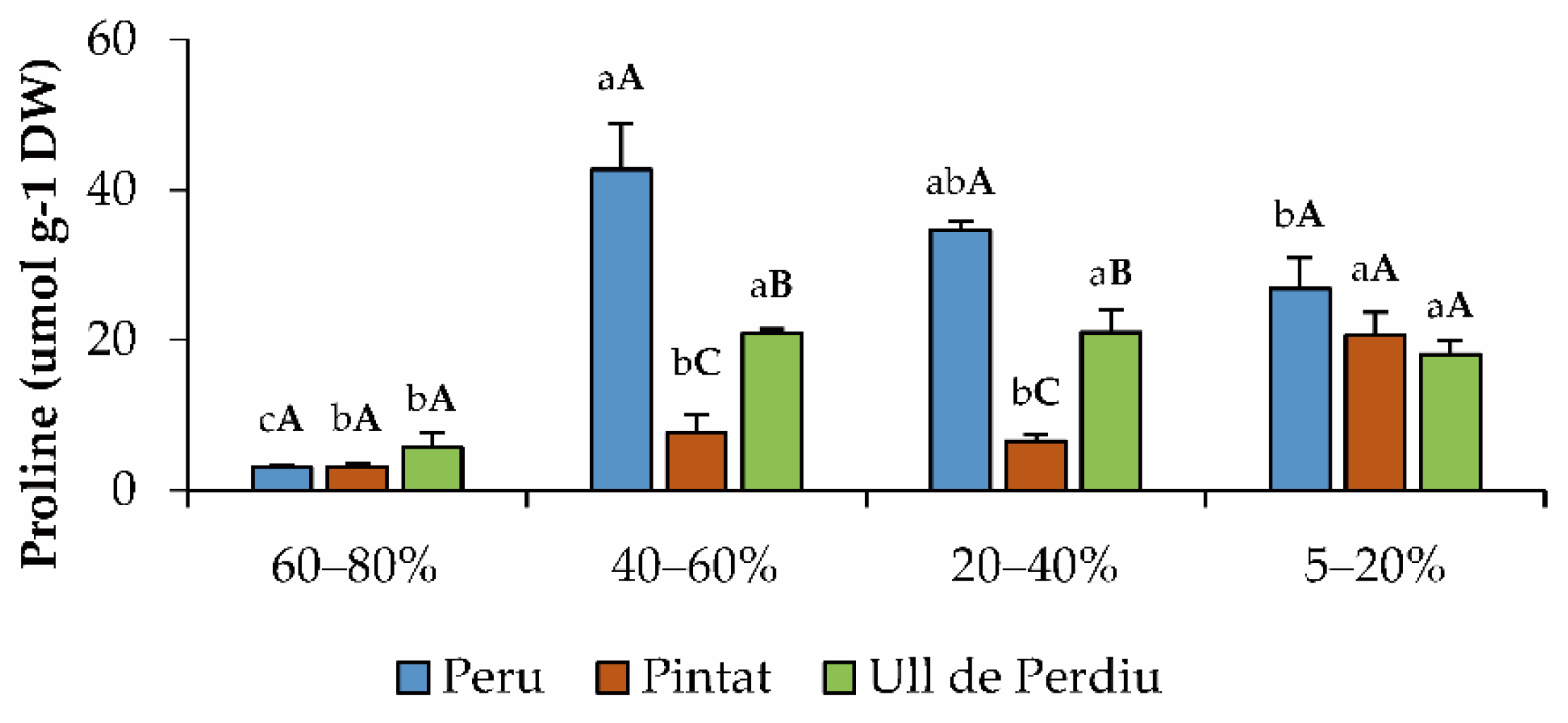
3.4. Osmolyte Quantification
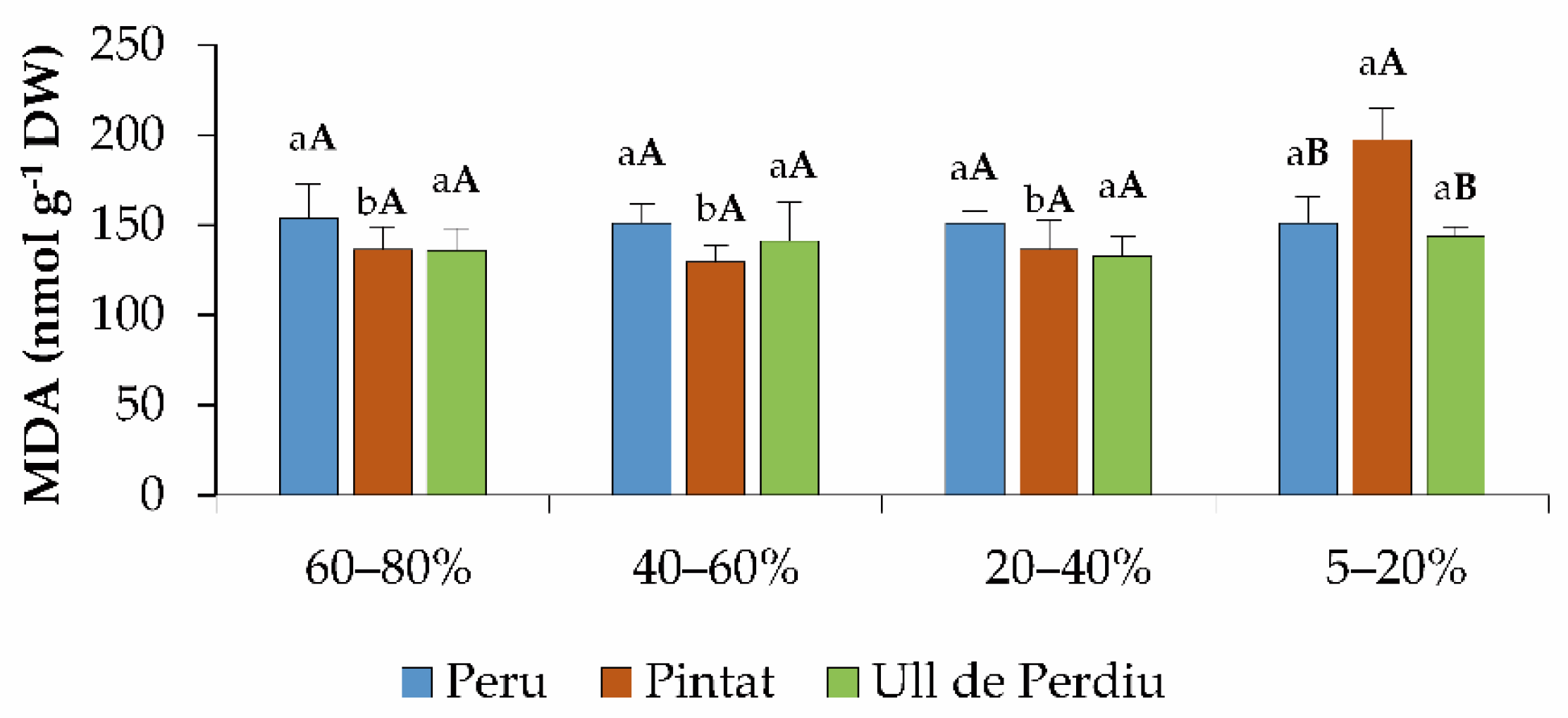
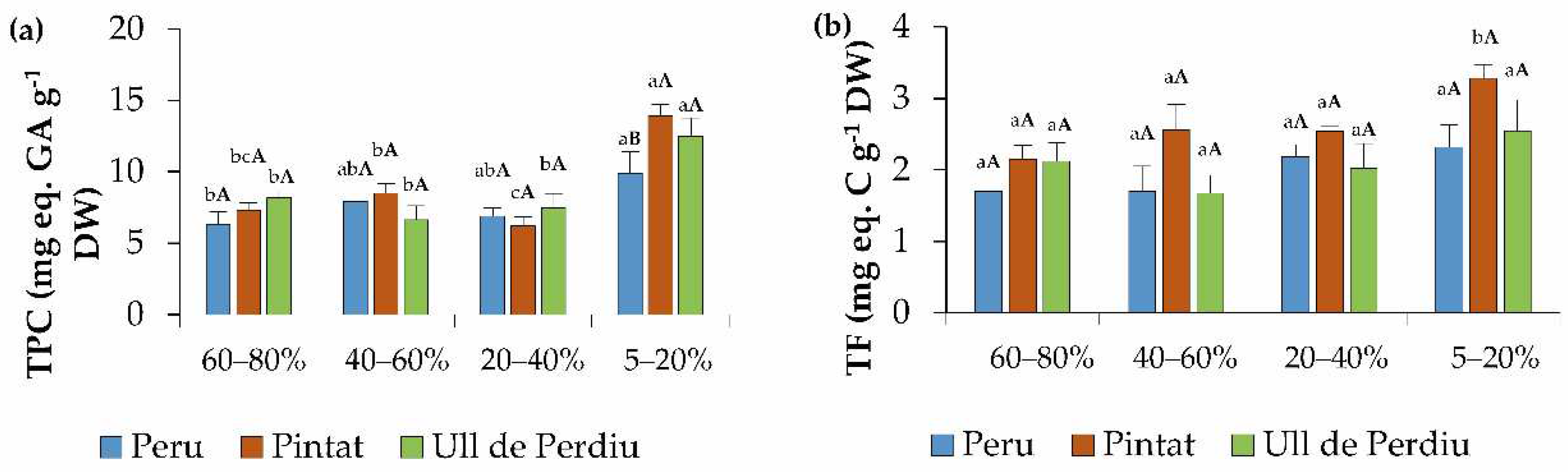
3.5. Oxidative Stress Markers and Non-Enzymatic Antioxidants
3.6. Effects of the Studied Parameters in Water Stress Response of P. lunatus
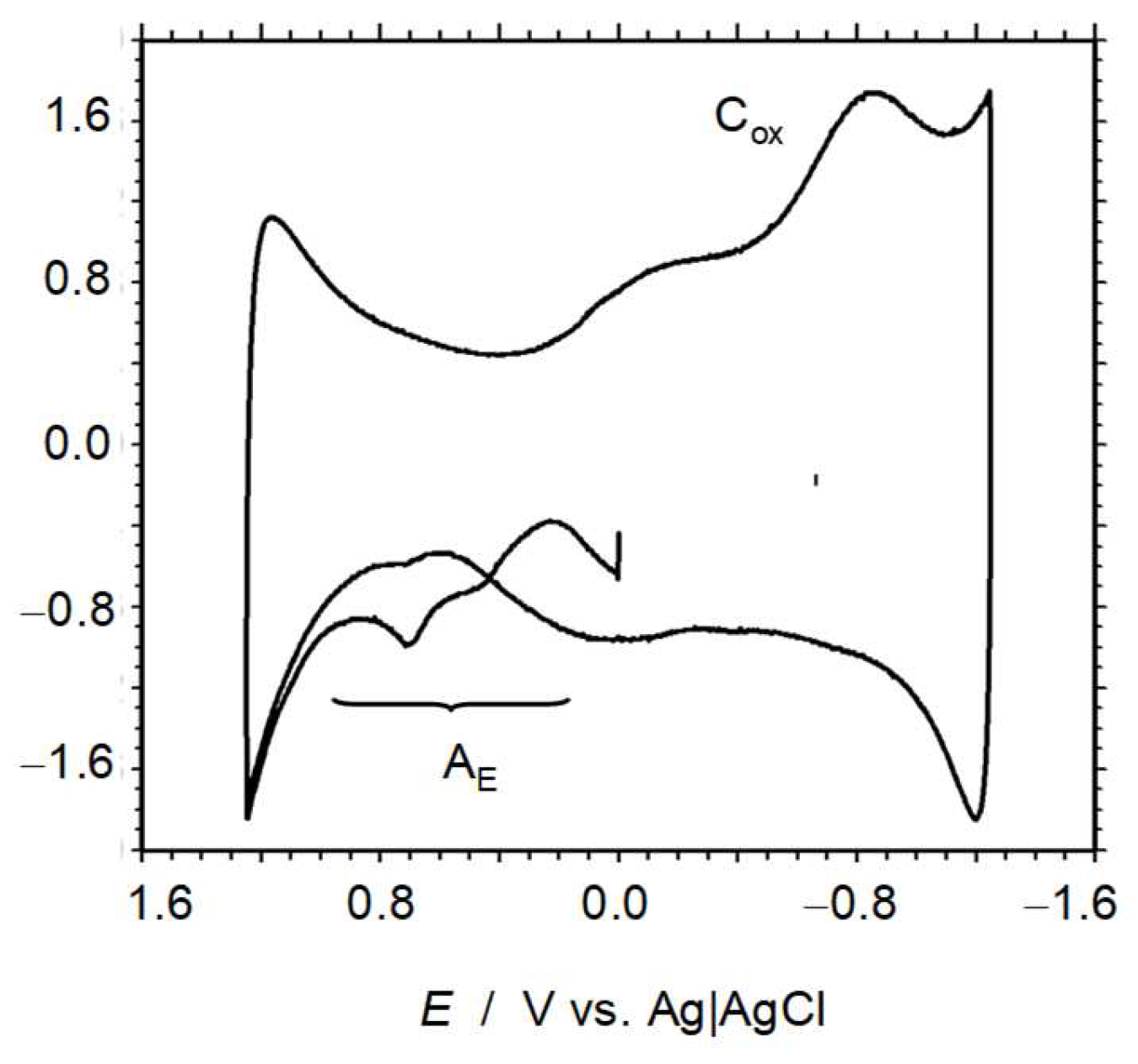
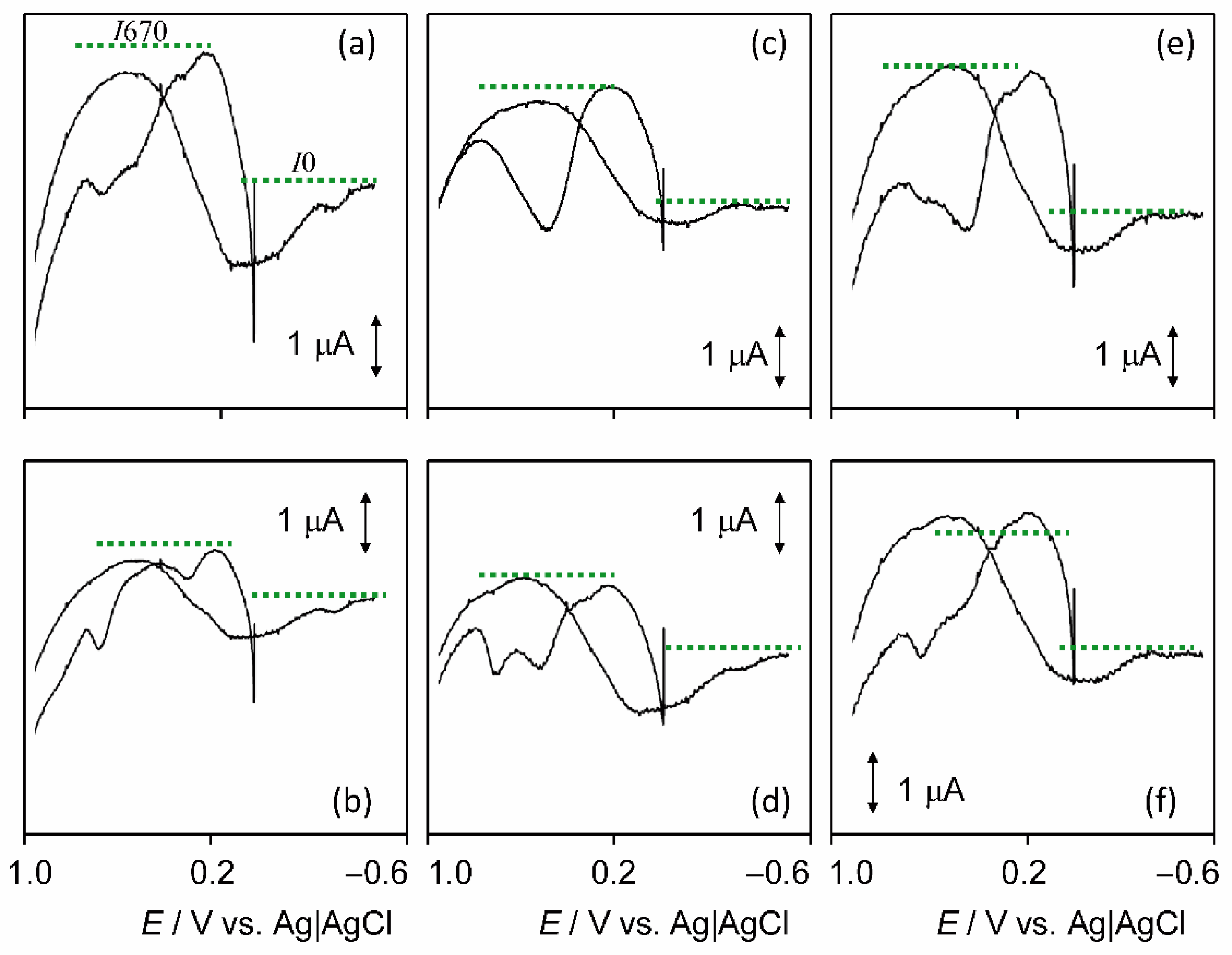
3.7. Electrochemical Experiments
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Food and Agriculture: Key to Achieving the 2030 Agenda for Sustainable Development; Job No. I5499; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016; p. 23. [Google Scholar]
- FAO. Transforming Our World: The 2030 Agenda for Sustainable Development. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/ (accessed on 27 May 2022).
- Brundtland, G.H. What is sustainable development. In Our Common Future; Oxford University Press: New York, NY, USA, 1987. [Google Scholar]
- UN. Transforming Our World: The 2030 Agenda for Sustainable Development. UN General Assembly, 25 September 2015. Available online: http://www.un.org/ga/search/view_doc.asp?symbol=A/70/L.1&referer=https://www.google.es/&Lang=E.2015 (accessed on 1 May 2022).
- Omilola, B.; Robele, S. The Central Position of Agriculture within the 2030 Agenda for Sustainable Development; International Food Policy Research Institute: Washington, DC, USA, 2017; Volume 1683. [Google Scholar]
- Fita, A.; Rodríguez-Burruezo, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Breeding and domesticating crops adapted to drought and salinity: A new paradigm for increasing food production. Front. Plant Sci. 2015, 6, 978. [Google Scholar] [CrossRef] [Green Version]
- Morton, J.F. The impact of climate change on smallholder and subsistence agriculture. Proc. Natl. Acad. Sci. USA 2007, 104, 19680–19685. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Chen, F.; Lin, X.; Liu, Z.; Zhang, H.; Zhao, J.; Kenan, L.; Ye, Q.; Li, Y.; Lv, S.; et al. Potential benefits of climate change for crop productivity in China. Agric. For. Meteorol. 2015, 208, 76–84. [Google Scholar] [CrossRef]
- Singh, S.; Vikram, P.; Sehgal, D.; Burgueño, J.; Sharma, A.; Singh, S.K.; Sansaloni, C.P.; Joynson, R.; Brabbs, T.; Ortiz, C.; et al. Harnessing genetic potential of wheat germplasm banks through impact-oriented-prebreeding for future food and nutritional security. Sci. Rep. 2018, 8, 12527. [Google Scholar] [CrossRef] [Green Version]
- Gharib, M.A.A.H.; Qabil, N.; Salem, A.H.; Ali, M.M.A.; Awaad, H.A.; Mansour, E. Characterization of wheat landraces and commercial cultivars based on morpho-phenological and agronomic traits. Cereal Res. Commun. 2021, 49, 149–159. [Google Scholar] [CrossRef]
- Tapan, K.; Bharadwaj, C.; Rizvi, A.H.; Ashutosh, S.; Shailesh, T.; Afroz, A.; Chauhan, S.K. Chickpea landraces: A valuable and divergent source for drought tolerance. Int. J. Trop. Agric. 2015, 33, 633–638. [Google Scholar]
- Nelimor, C.; Badu-Apraku, B.; Tetteh, A.Y.; Garcia-Oliveira, A.L.; N’guetta, A.S.P. Assessing the potential of extra-early maturing landraces for improving tolerance to drought, heat, and both combined stresses in maize. Agronomy 2020, 10, 318. [Google Scholar] [CrossRef] [Green Version]
- Ochieng, G.; Ngugi, K.; Wamalwa, L.N.; Manyasa, E.; Muchira, N.; Nyamongo, D.; Odeny, D.A. Novel sources of drought tolerance from landraces and wild sorghum relatives. Crop Sci. 2021, 61, 104–118. [Google Scholar] [CrossRef]
- Li, X.; Liu, F. Drought stress memory and drought stress tolerance in plants: Biochemical and molecular basis. In Drought Stress Tolerance in Plants; Hossain, M., Wani, S., Bhattacharjee, S., Burritt, D., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2016; Volume 1, pp. 17–44. [Google Scholar]
- Mantri, N.; Patade, V.; Penna, S.; Ford, R.; Pang, E. Abiotic stress responses in plants: Present and future. In Abiotic Stress Responses in Plants: Metabolism, Productivity and Sustainability; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 1–19. [Google Scholar]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [Green Version]
- Kawamitsu, Y.; Driscoll, T.; Boyer, J.S. Photosynthesis during desiccation in an intertidal alga and a land plant. Plant. Cell Physiol. 2000, 41, 344–353. [Google Scholar] [CrossRef] [Green Version]
- Boyer, J.S. Differing sensitivity of photosynthesis to low leaf water potentials in corn and soybean. Plant physiol. 1970, 46, 236–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iturbe-Ormaetxe, I.; Escuredo, P.R.; Arrese-Igor, C.; Becana, M. Oxidative damage in pea plants exposed to water deficit or paraquat. Plant physiol. 1998, 116, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Manivannan, P.; Jaleel, C.A.; Sankar, B.; Kishorekumar, A.; Somasundaram, R.; Lakshmanan, G.A.; Panneerselvam, R. Growth, biochemical modifications and proline metabolism in Helianthus annuus L. as induced by drought stress. Colloids Surf. B Biointerfaces 2007, 59, 141–149. [Google Scholar] [CrossRef]
- Smirnoff, N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef] [PubMed]
- Del Río, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef]
- Mansori, M.; Chernane, H.; Latique, S.; Benaliat, A.; Hsissou, D.; El Kaoua, M. Seaweed extract effect on water deficit and antioxidative mechanisms in bean plants (Phaseolus vulgaris L.). J. Appl. Phycol. 2015, 27, 1689–1698. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharma, S. Proline metabolism and its implications for plant-environment interaction. In Arabidopsis Book; American Society of Plant Biologists: Washington, DC, USA, 2010; p. 8e0140. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Iram, A. Drought stress induced changes in some organic substances in nodules and other plant parts of two potential legumes differing in salt tolerance. Flora 2005, 200, 535–546. [Google Scholar] [CrossRef]
- Svetleva, D.; Krastev, V.; Dimova, D.; Mitrovska, Z.; Miteva, D.; Parvanova, P.; Chankova, S. Drought tolerance of Bulgarian common bean genotypes, characterised by some biochemical markers for oxidative stress. Cent. Eur. Agric. J. 2012, 13, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Rosales, M.A.; Ocampo, O.; Rodríguez-Valentín, R.; Olvera-Carrillo, Y.; Acosta-Gallegos, J.; Covarrubias, A. Physiological analysis of common bean (Phaseolus vulgaris L.) cultivars uncovers characteristics related to terminal drought resistance. Plant Physiol. Biochem. 2012, 56, 24–34. [Google Scholar] [CrossRef]
- Morosan, M.; Al Hassan, M.; Naranjo, M.; Lopez-Gresa, M.P.; Vicente, O. Comparative analysis of drough stress responses in Phaseolus vulgaris (common bean) and P. coccineus (runner bean) cultivar. EuroBiotech. J. 2017, 1, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Scholz, F.; Schröder, U.; Gulaboski, R.; Doménech-Carbó, A. Electrochemistry of Immobilized Particles and Droplets, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Doménech-Carbó, A.; Montoya, N.; Soriano, P.; Estrelles, E. An electrochemical analysis suggests role of gynodioecy in adaptation to stress in Cortaderia selloana. Curr. Plant Biol. 2018, 16, 9–14. [Google Scholar] [CrossRef]
- Marselli, B.; Garcia-Gomez, J.; Michaud, P.A.; Rodrigo, M.A.; Comninellis, C. Electrogeneration of hydroxyl radicals on boron-doped diamond electrodes. J. Electrochem. Soc. 2003, 150, D79. [Google Scholar] [CrossRef]
- Enache, T.A.; Chiorcea-Paquim, A.M.; Fatibello-Filho, O.; Oliveira-Brett, A.M. Hydroxyl radicals electrochemically generated in situ on a boron-doped diamond electrode. Electrochem. Commun. 2009, 11, 1342–1345. [Google Scholar] [CrossRef]
- Kapałka, A.; Fóti, G.; Comninellis, C. The importance of electrode material in environmental electrochemistry: Formation and reactivity of free hydroxyl radicals on boron-doped diamond electrodes. Electrochim. Acta 2009, 54, 2018–2023. [Google Scholar] [CrossRef]
- Bellucci, E.; Bitocchi, E.; Rau, D.; Rodriguez, M.; Biagetti, E.; Giardini, A.; Attene, G.; Nanni, L.; Papa, R. Genomics of origin, domestication and evolution of Phaseolus vulgaris. In Genomics of Plant Genetic Resources; Tuberosa, R., Graner, A., Frison, E., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 483–507. [Google Scholar]
- Martínez-Nieto, M.I.; Estrelles, E.; Prieto-Mossi, J.; Roselló, J.; Soriano, P. Resilience capacity assessment of the traditional Lima Bean (Phaseolus lunatus L.) landraces facing climate change. Agronomy 2020, 10, 758. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Weimberg, R. Solute adjustments in leaves of two species of wheat at two different stages of growth in response to salinity. Physiol. Plant. 1987, 70, 381–388. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Gil, R.; Bautista, I.; Boscaiu, M.; Lidón, A.; Wankhade, S.; Sánchez, H.; Lilinares, J.; Vicente, O. Responses of five Mediterranean halophytes to seasonal changes in environmental conditions. AoB Plants 2014, 6, plu049. [Google Scholar] [CrossRef] [PubMed]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Taulavuori, E.; Hellström, E.K.; Taulavuori, K.; Laine, K. Comparison of two methods used to analyse lipid peroxidation from Vaccinium myrtillus (L.) during snow removal, reacclimation and cold acclimation. J. Exp. Bot. 2001, 52, 2375–2380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blainski, A.; Lopes, G.C.; De Mello, J.C.P. Application and analysis of the folin ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Grothmann, T.; Patt, A. Adaptive capacity and human cognition: The process of individual adaptation to climate change. Glob. Environ. Change. 2005, 15, 199–213. [Google Scholar] [CrossRef]
- Aguilar-Benítez, G.; Peña-Valdivia, C.B.; Vega, J.R.; Castro-Rivera, R.; Ramírez-Tobías, H.M. Seed germination and early root growth in common bean and maize landraces and improved cultivars at different water stress levels. Int. J. Appl. Sci. Technol. 2014, 4, 118–127. [Google Scholar]
- Azeez, M.A.; Adubi, A.O.; Durodola, F.A. Landraces and crop genetic improvement. In Rediscovery of Landraces as a Resource for the Future; Grillo, O., Ed.; IntechOpen: London, UK, 2018; pp. 1–20. [Google Scholar]
- Petropoulos, S.A.; Barros, L.; Ferreira, I.C. Rediscovering local landraces: Shaping horticulture for the future. Front. Plant Sci. 2019, 10, 126. [Google Scholar] [CrossRef] [Green Version]
- Mabhaudhi, T. Responses of Maize (Zea mays L.) Landraces to Water Stress Compared with Commercial Hybrids. Ph.D. Dissertation, University of KwaZulu-Natal, Pietermaritzburg, South Africa, January 2009. [Google Scholar]
- Sithole, N.; Modi, A.T. Responses of selected bottle gourd [Lagenaria siceraria (Molina Standly)] landraces to water stress. Acta Agric. Scand. Sect. B Soil Plant Sci. 2015, 65, 350–356. [Google Scholar] [CrossRef]
- Widuri, L.I.; Lakitan, B.; Sodikin, E.; Hasmeda, M.; Meihana, M.; Kartika, K.; Siaga, E. Shoot and root growth in common bean (Phaseolus vulgaris L.) exposed to gradual drought stress. AGRIVITA J. Agric. Sci. 2018, 40, 442–452. [Google Scholar] [CrossRef]
- IIaque, M.E.; Chang, T.T.; Tepora, N.; Loresto, G.C. Root characters as selection indices for drought resistance in rice. Philipp. J. Crop. Sci. 1989, 14, 7–9. [Google Scholar]
- Midaoui, E.M.; Serieys, H.; Griveau, Y.; Benbella, M.; Talouizte, A.; Bervillé, A.; Kaan, F. Effects of osmotic and water stresses on root and shoot morphology and seed yield in sunflower (Helianthus annuus L.) genotypes bred for Morocco or issued from introgression with H. argophyllus T. & G. Helia 2003, 26, 1–15. [Google Scholar]
- Khalil, A.M.; Murchie, E.H.; Mooney, S.J. Quantifying the influence of water deficit on root and shoot growth in wheat using X-ray Computed Tomography. AoB Plants 2020, 12, plaa036. [Google Scholar] [CrossRef] [PubMed]
- Arteaga, S.; Yabor, L.; Díez, M.J.; Prohens, J.; Boscaiu, M.; Vicente, O. The use of proline in screening for tolerance to drought and salinity in common bean (Phaseolus vulgaris L.) genotypes. Agronomy 2020, 10, 817. [Google Scholar] [CrossRef]
- Thinley, J.; Dorji, C. Screening of beans (Phaseolus vulgaris L.) genotypes for drought tolerance. BioRxiv 2021, 1–30. [Google Scholar] [CrossRef]
- Emam, Y.; Shekoofa, A.; Salehi, F.; Jalali, A.H. Water stress effects on two common bean cultivars with contrasting growth habits. Am. Eurasian J. Agric. Environ Sci. 2010, 9, 495–499. [Google Scholar]
- Boshkovski, B.; Tzerakis, C.; Doupis, G.; Zapolska, A.; Kalaitzidis, C.; Koubouris, G. Relationships of spectral reflectance with plant tissue mineral elements of common bean (Phaseolus vulgaris L.) under drought and salinity stresses. Commun. Soil Sci. Plant Anal. 2020, 51, 675–686. [Google Scholar] [CrossRef]
- Doğan, N.; Akinci, Ş. Effects of water stress on the uptake of nutrients by bean seedlings (Phaseolus vulgaris L.). Fresenius Environ. Bull. 2011, 20, 2163–2173. [Google Scholar]
- Pacheco, J.; Plazas, M.; Pettinari, I.; Landa-Faz, A.; González-Orenga, S.; Boscaiu, M.; Soler, S.; Prohens, J.; Vicente, O.; Gramazio, P. Moderate and severe water stress effects on morphological and biochemical traits in a set of pepino (Solanum muricatum) cultivars. Sci. Hortic. 2021, 284, 110143. [Google Scholar] [CrossRef]
- Xi, J.J.; Chen, H.Y.; Bai, W.P.; Yang, R.C.; Yang, P.Z.; Chen, R.J.; Tian-Ming Hu & Wang, S.M. Sodium-related adaptations to drought: New insights from the xerophyte plant Zygophyllum xanthoxylum. Front. Plant Sci. 2018, 9, 1678. [Google Scholar] [CrossRef]
- Arteaga, S.; Al Hassan, M.; Chaminda Bandara, W.M.; Yabor, L.; Llinares, J.V.; Boscaiu, M.; Vicente, O. Screening for salt tolerance in four local varieties of Phaseolus lunatus from Spain. Agriculture 2018, 8, 201. [Google Scholar] [CrossRef] [Green Version]
- Tuteja, N.; Mahajan, S. Calcium signaling network in plants: An overview. Plant Signal. Behav. 2007, 2, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Bose, J.; Pottosin, I.I.; Shabala, S.S.; Palmgren, M.G.; Shabala, S. Calcium efflux systems in stress signaling and adaptation in plants. Front. Plant Sci. 2011, 2, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darkwa, K.; Ambachew, D.; Mohammed, H.; Asfaw, A.; Blair, M.W. Evaluation of common bean (Phaseolus vulgaris L.) genotypes for drought stress adaptation in Ethiopia. Crop J. 2016, 4, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Kusvuran, S.; Dasgan, H.Y. Effects of drought stress on physiological and biochemical changes in Phaseolus vulgaris L. Legume Res. 2017, 40, 55–62. [Google Scholar]
- Mafakheri, A.; Siosemardeh, A.F.; Bahramnejad, B.; Struik, P.C.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 8, 580–585. [Google Scholar]
- Singer, S.M.; Helmy, Y.I.; Karas, A.N.; Abou-Hadid, A.F. Influences of different water-stress treatments on growth, development and production of snap bean (Phaseolus vulgaris. L.). Acta Hortic. 2003, 614, 605–611. [Google Scholar] [CrossRef]
- Dolatabadian, A.; Sanavy, S.M.; Chashmi, N.A. The effects of foliar application of ascorbic acid (vitamin C) on antioxidant enzymes activities, lipid peroxidation and proline accumulation of canola (Brassica napus L.) under conditions of salt stress. J. Agron. Crop Sci. 2008, 194, 206–213. [Google Scholar] [CrossRef]
- Kusvuran, A. The effects of salt stress on the germination and antioxidative enzyme activity of Hungarian vetch (Vicia pannonica Crantz.) varieties. Legume Res. 2015, 38, 51–59. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Y.; Ma, X. Different response on drought tolerance and post-drought recovery between the small-leafed and the large-leafed white clover (Trifolium repens L.) associated with antioxidative enzyme protection and lignin metabolism. Acta Physiol. Plant. 2013, 35, 213–222. [Google Scholar] [CrossRef]
- Doménech-Carbó, A.; Cervelló-Bulls, P.; González, J.M.; Soriano, P.; Estrelles, E.; Montoya, N. Electrochemical monitoring of ROS influence on seedlings and germination response to salinity stress of three species of the tribe Inuleae. RSC Adv. 2019, 9, 17856–17867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Varia, F.; Macaluso, D.; Vaccaro, A.; Caruso, P.; Guccione, G.D. The Adoption of Landraces of Durum Wheat in Sicilian Organic Cereal Farming Analysed Using a System Dynamics Approach. Agronomy 2021, 11, 319. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Moisture | Peru | Pintat | Ull de Perdiu | |
|---|---|---|---|---|
| Na+ Root (μmol g−1 DW) | 60–80% | 295.91 ± 34.86 bB | 321.65 ± 36.32 bB | 503.79 ± 34.02 aA |
| 40–60% | 499.80 ± 38.87 aA | 381.07 ± 74.26 abA | 487.52 ± 73.23 abA | |
| 20–40% | 393.37 ± 58.19 abA | 364.51 ± 17.94 abA | 390.07 ± 34.04 abA | |
| 5–20% | 380.12 ± 53.63 abA | 540.28 ± 90.49 aB | 343.88 ± 48.53 bA | |
| Na+ Leaves (μmol g−1 DW) | 60–80% | 308.23 ± 34.86 aA | 250.67 ± 41.36 bA | 269.13 ± 50.06 aA |
| 40–60% | 302.71 ± 33.89 aA | 326.87 ± 15.10 aA | 269.02 ± 24.02 aA | |
| 20–40% | 301.17 ± 32.76 aA | 313.45 ± 6.95 abA | 276.01 ± 17.49 aA | |
| 5–20% | 263.03 ± 09.65 aB | 330.62 ± 11.14 aA | 356.82 ± 30.56 aA | |
| Cl− Root (μmol g−1 DW) | 60–80% | 227.38 ± 25.72 bcAB | 177.69 ± 19.95 aB | 301.86 ± 33.22 aA |
| 40–60% | 338.73 ± 48.57 abA * | 239.55 ± 38.95 aA | 281.08 ± 40.04 aA | |
| 20–40% | 439.60 ± 56.82 aA * | 287.40 ± 57.14 aA | 305.70 ± 25.62 aA | |
| 5–20% | 178.87 ± 19.91 cA | 178.45 ± 48.00 aA | 245.98 ± 31.55 aA | |
| Cl− Leaves (μmol g−1 DW) | 60–80% | 181.65 ± 41.12 aB | 356.60 ± 68.63 aA | 204.14 ± 25.19 aAB |
| 40–60% | 152.74 ± 18.00 aA | 180.40 ± 37.73 bA | 217.15 ± 58.82 aA | |
| 20–40% | 204.76 ± 22.70 aA | 222.01 ± 42.04 abA | 262.92 ± 29.17 aA | |
| 5–20% | 186.88 ± 21.27 aA | 230.95 ± 56.43 abA | 257.46 ± 4.31 aA | |
| K+ Root (μmol g−1 DW) | 60–80% | 467.11 ± 67.33 bA | 353.13 ± 95.19 cA | 424.67 ± 94.20 bA |
| 40–60 % | 618.32 ± 58.03 abA | 460.17 ± 42.41 bcA | 488.54 ± 76.55 bA | |
| 20–40% | 707.85 ± 58.46 aA | 572.51 ± 47.04 bAB | 473.84 ± 29.83 bB | |
| 5–20% | 791.67 ± 26.97 aA | 798.49 ± 33.52 aA | 774.64 ± 66.95 aA* | |
| K+ Leaves (μmol g−1 DW) | 60–80% | 288.55 ± 67.07 bC | 640.44 ± 63.90 aB* | 957.82 ± 91.66 aA* |
| 40–60% | 662.42 ± 109.57 abA | 561.75 ± 82.32 aA | 615.66 ± 106.76 bA | |
| 20–40% | 799.27 ± 193.30 aA | 493.82 ± 47.53 aA | 513.25 ± 53.76 abA | |
| 5–20% | 671.87 ± 132.63 abA | 679.04 ± 155.30 aA | 294.61 ± 51.19 cB | |
| Ca2+ Root (μmol g−1 DW) | 60–80% | 9.44 ± 2.58 bA | 21.36 ± 8.41 bA | 22.07 ± 12.36 aA |
| 40–60% | 21.09 ± 7.25 bA | 17.88 ± 0.77 bA | 19.22 ± 5.71 aA | |
| 20–40% | 69.45 ± 9.18 aA | 42.24 ± 3.36 aA | 16.45 ± 2.47 aB | |
| 5–20% | 27.28 ± 6.69 bA | 11.49 ± 2.48 bB | 8.77 ± 0.23 aB | |
| Ca2+ Leaves (μmol g−1 DW) | 60–80% | 145.22 ± 25.83 bB * | 309.39 ± 24.20 aA * | 178.85 ± 9.84 bcB * |
| 40–60% | 258.61 ± 18.42 aAB * | 272.98 ± 31.37 aA * | 149.92 ± 47.92 cB * | |
| 20–40% | 323.95 ± 40.42 aA * | 301.53 ± 23.55 aA * | 278.91 ± 38.99 aA * | |
| 5–20% | 139.14 ± 20.58 bB * | 178.59 ± 29.76 bB * | 259.60 ± 15.33 abA * |
| Trait | Cultivar | Treatment | Interaction | Residual |
|---|---|---|---|---|
| Stem Length (cm) | 9.48 * | 42.09 *** | 2.27 n.s. | 46.16 |
| Leaves Fresh Weight | 0.30 n.s. | 59.54 *** | 3.64 n.s. | 36.51 |
| Leaves: % Water | 0.03 n.s. | 12.67 n.s. | 3.55 n.s. | 83.79 |
| Root Fresh Weight | 2.18 n.s. | 50.75 *** | 2.93 n.s. | 44.15 |
| Root: % water | 1.13 n.s. | 69.03 *** | 2.15 n.s. | 25.23 |
| Leaves_Na+ | 0.77 n.s. | 4.92 n.s. | 16.47 n.s. | 77.84 |
| Leaves_K+ | 0.04 n.s. | 1.18 n.s. | 41.24 *** | 57.55 |
| Leaves_Ca2+ | 6.74 * | 22.62 *** | 28.98 *** | 43.63 |
| Leaves_Cl− | 8.51 n.s. | 6.26 n.s. | 14.11 n.s. | 69.38 |
| Root_Na+ | 1.60 n.s. | 6.25 n.s. | 25.73 * | 66.43 |
| Root_K+ | 6.19 n.s. | 48.76 *** | 4.61 n.s. | 40.08 |
| Root_Ca2+ | 9.44 * | 28.76 *** | 25.93 ** | 36.70 |
| Root_Cl− | 9.49 * | 25.83 ** | 12.22 n.s. | 53.62 |
| Proline | 17.37 *** | 23.28 *** | 30.71 *** | 30.29 |
| Chl a | 0.97 n.s. | 17.64 * | 10.10 n.s. | 71.94 |
| Chl b | 0.60 n.s. | 31.55 *** | 8.75 n.s. | 59.09 |
| Carotenoids | 1.98 n.s. | 22.38 * | 5.33 n.s. | 70.95 |
| Flavonoids | 16.43 ** | 17.99 ** | 4.97 n.s. | 60.62 |
| Phenols | 2.99 n.s. | 48.92 *** | 8.56 n.s. | 39.53 |
| MDA | 0.01 n.s. | 16.56 * | 12.03 n.s. | 71.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Nieto, M.I.; González-Orenga, S.; Soriano, P.; Prieto-Mossi, J.; Larrea, E.; Doménech-Carbó, A.; Tofei, A.M.; Vicente, O.; Mayoral, O. Are Traditional Lima Bean (Phaseolus lunatus L.) Landraces Valuable to Cope with Climate Change? Effects of Drought on Growth and Biochemical Stress Markers. Agronomy 2022, 12, 1715. https://doi.org/10.3390/agronomy12071715
Martínez-Nieto MI, González-Orenga S, Soriano P, Prieto-Mossi J, Larrea E, Doménech-Carbó A, Tofei AM, Vicente O, Mayoral O. Are Traditional Lima Bean (Phaseolus lunatus L.) Landraces Valuable to Cope with Climate Change? Effects of Drought on Growth and Biochemical Stress Markers. Agronomy. 2022; 12(7):1715. https://doi.org/10.3390/agronomy12071715
Chicago/Turabian StyleMartínez-Nieto, M. Isabel, Sara González-Orenga, Pilar Soriano, Josefa Prieto-Mossi, Elena Larrea, Antonio Doménech-Carbó, Ana Maria Tofei, Oscar Vicente, and Olga Mayoral. 2022. "Are Traditional Lima Bean (Phaseolus lunatus L.) Landraces Valuable to Cope with Climate Change? Effects of Drought on Growth and Biochemical Stress Markers" Agronomy 12, no. 7: 1715. https://doi.org/10.3390/agronomy12071715