
Exogenous Application of GABA Alleviates Alkali Damage in Alfalfa by Increasing the Activities of Antioxidant Enzymes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Germination and Treatments
2.2. Alkali Stress Treatments
2.3. Physio-Biochemical Analysis of Plant Materials
2.4. Determination of GAD Enzyme Activity by a Colorimetric Method
2.5. Determination of Organic Acid Content by High-Performance Liquid Chromatography
2.6. Determination of GABA-T Activity, Glu and GABA Content by the Microplate Method
2.7. RNA Extraction and qRT–PCR Analysis
2.8. Statistical Analysis
3. Results
3.1. Determination of the Optimal Concentration of GABA for Exogenous Application
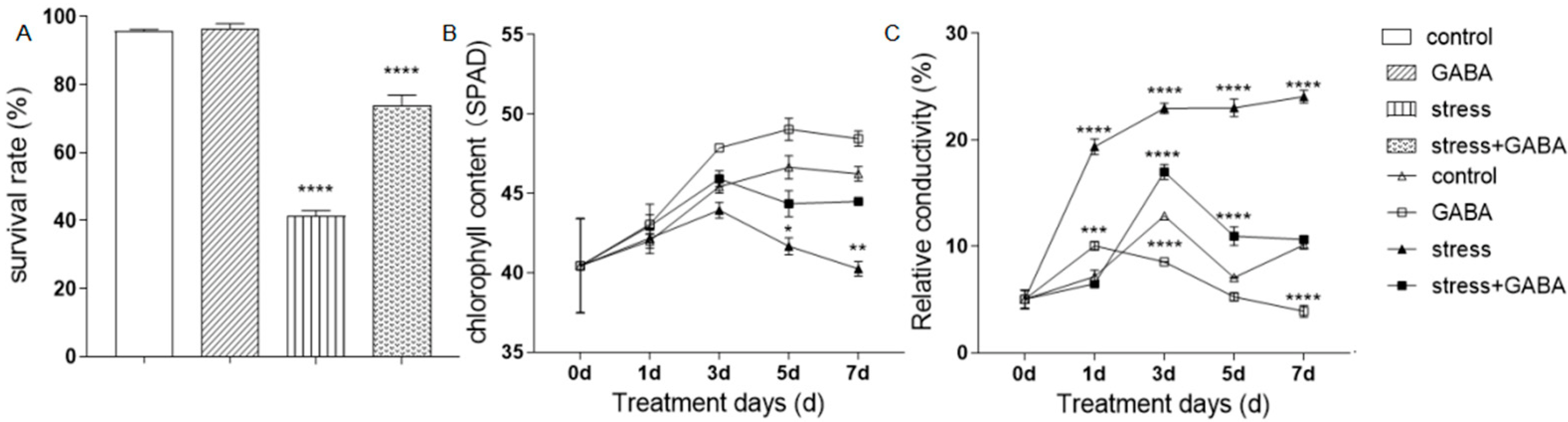
3.2. Effect of Exogenous GABA on Alkali Tolerance in Alfalfa
3.3. Effect of Exogenous GABA on the Levels of Osmotically Active Substances
3.4. Effects of Exogenous GABA on the Activity of Antioxidant Enzymes and the Production of Reactive Oxygen Species under Alkali Stress Conditions
3.5. Effects of Exogenous GABA on the Content of Metabolites and the Activity of Enzymes in the GABA Pathway under Alkali Stress
3.6. Effects of Exogenous GABA on the Levels of Expression of Four Genes Related to GAD and Those of Other Related Genes in Leaves under Alkali Stress
3.7. Effect of GABA Application on the Growth of Alfalfa under Alkali Stress Conditions
4. Discussion
4.1. Exogenous GABA Improved Alkali Stress Tolerance by Alleviating the Phenotypic Symptoms of Alkali Damage
4.2. Exogenous GABA Regulated the Content of Osmotically Active Substances in the Leaves of Plants Exposed to Alkali Stress
4.3. Exogenous GABA Improved Alkali Stress Tolerance by Regulating the Antioxidant System
4.4. Regulation of Exogenous GABA via the GABA Metabolic Pathway in Alfalfa Seedlings under Alkali Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hendry, G.W. Alfalfa in History. Agron. J. 1923, 5, 171–176. [Google Scholar] [CrossRef]
- Bouton, J.H. An overview of the role of lucerne (Medicago sativa L.) in pastoral agriculture. Crop Pasture Sci. 2012, 63, 734–738. [Google Scholar] [CrossRef]
- Peng, Y.L.; Gao, Z.W.; Gao, Y.; Liu, G.F.; Sheng, L.X.; Wang, D.L. Eco-physiological Characteristics of Alfalfa Seedlings in Response to Various Mixed Salt-alkaline Stresses. Chin. Bot. Engl. Version 2008, 50, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.I.; Jalil, S.U.; Ansari, S.A.; Hasanuzzaman, M. GABA shunt: A key-player in mitigation of ROS during stress. Plant Growth Regul. 2021, 94, 129–139. [Google Scholar] [CrossRef]
- Tang, M.; Li, Z.; Luo, L.; Cheng, B.; Zhang, Y.; Zeng, W.; Peng, Y. Nitric Oxide Signal, Nitrogen Metabolism, and Water Balance Affected by γ-Aminobutyric Acid (GABA) in Relation to Enhanced Tolerance to Water Stress in Creeping Bentgrass. Int. J. Mol. Sci. 2020, 21, 7460. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Jia, Q.; Ji, S.; Gong, B.; Li, J.; Lü, G.; Gao, H. Gamma-aminobutyric acid (GABA) alleviates salt damage in tomato by modulating Na+ uptake, the GAD gene, amino acid synthesis and reactive oxygen species metabolism. BMC Plant Biol. 2020, 20, 465. [Google Scholar] [CrossRef]
- Li, Z.; Yu, J.; Peng, Y.; Huang, B. Metabolic pathways regulated by γ-aminobutyric acid (GABA) contributing to heat tolerance in creeping bentgrass (Agrostis stolonifera). Sci. Rep. 2016, 6, 30338. [Google Scholar] [CrossRef] [Green Version]
- Palma, F.; Carvajal, F.; Jiménez-Muñoz, R.; Pulido, A.; Jamilena, M.; Garrido, D. Exogenous γ-aminobutyric acid treatment improves the cold tolerance of zucchini fruit during postharvest storage. Plant Physiol. Biochem. 2019, 136, 188–195. [Google Scholar] [CrossRef]
- Qing-ping, X.I.; Hong-bo, G.A.O.; Jing-rui, L.I. Effects of γ-aminobutyric acid (GABA) on photosynthesis and chlorophyll fluorescence parameters of melon seedlings under hypoxic stress. Chin. J. Appl. Ecol. 2011, 22, 8. [Google Scholar]
- Qu, Z.; Jia, Y.; Duan, Y.; Chen, H.; Wang, X.; Zheng, H.; Zhao, H. Research progress on metabolism and function of γ-aminobutyric acid in crops under stress. J. Crops 2014, 7, 12. [Google Scholar]
- Yang, L. Effects of γ-aminobutyric acid on the metabolism of reactive oxygen species and the expression of key enzymes in melon seedlings under hypoxic stress. Hebei Agric. Univ. 2012, 23, 6. [Google Scholar]
- Kabir, A.H.; Hossain, H.H.; Khatun, M.A.; Mandal, A.; Haider, S.A. Role of Silicon Counteracting Cadmium Toxicity in Alfalfa (Medicago sativa L.). Front. Plant Sci. 2016, 7, 1117. [Google Scholar]
- Cheng, B.; Li, Z.; Liang, L.; Cao, Y.; Zeng, W.; Zhang, X.; Peng, Y. The γ-Aminobutyric Acid (GABA) Alleviates Salt Stress Damage during Seeds Germination of White Clover Associated with Na+/K+ Transportation, Dehydrins Accumulation, and Stress-Related Genes Expression in White Clover. Int. J. Mol. Sci. 2018, 19, 2520. [Google Scholar]
- Kalhor, M.S.; Aliniaeifard, S.; Seif, M.; Asayesh, E.J.; Bernard, F.; Hassani, B.; Li, T. Title: Enhanced salt tolerance and photosynthetic performance: Implication of -amino butyric acid application in salt-exposed lettuce (Lactuca sativa L.) plants. Plant Physiol. Biochem. PPB 2018, 130, 157. [Google Scholar] [CrossRef] [PubMed]
- Sheteiwy, M.S.; Shao, H.; Qi, W.; Hamoud, Y.A.; Shaghaleh, H.; Khan, N.U.; Tang, B. GABA-Alleviated Oxidative Injury Induced by Salinity, Osmotic Stress and their Combination by Regulating Cellular and Molecular Signals in Rice. Int. J. Mol. Sci. 2019, 20, 5709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Li, M.; Mao, P.; Jiang, Y. Effects of Gamma-Aminobutyric Acid on Seed Germination, Ion Balance, and Metabolic Activity in Perennial Ryegrass Under Salinity Stress. J. Plant Growth Regul. 2022, 41, 1835–1844. [Google Scholar] [CrossRef]
- Brikis, C.J.; Adel, Z.; Trobacher, C.P.; DeEll, J.R.; Akama, K.; Mullen, R.T.; Bozzo, G.G.; Shelp, B.J. Ancient Plant Glyoxylate/Succinic Semialdehyde Reductases: GLYR1s Are Cytosolic, Whereas GLYR2s Are Localized to Both Mitochondria and Plastids. Front. Plant Sci. 2017, 8, 601. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gu, W.; Meng, Y.; Xie, T.; Li, L.; Li, J.; Wei, S. γ-Aminobutyric Acid Imparts Partial Protection from Salt Stress Injury to Maize Seedlings by Improving Photosynthesis and Upregulating Osmoprotectants and Antioxidants. Sci. Rep. 2017, 7, 43609. [Google Scholar] [CrossRef]
- Shi, S.Q.; Shi, Z.; Jiang, Z.P.; Qi, L.W.; Sun, X.M.; Li, C.X.; Zhang, S. Regulation of exogenous GABA on ethylene production in Caragana intermedia seedlings under NaCl stress. For. Sci. 2008, 9, 29–33. [Google Scholar]
- Nayyar, H.; Kaur, R.; Kaur, S.; Singh, R. γ-Aminobutyric Acid (GABA) Imparts Partial Protection from Heat Stress Injury to Rice Seedlings by Improving Leaf Turgor and Upregulating Osmoprotectants and Antioxidants. J. Plant Growth Regul. 2014, 33, 408–419. [Google Scholar] [CrossRef]
- Lu, X.; Li, L.; Hu, X.; Ma, H.E. Effects of exogenous GABA on the antioxidant function of the roots of Stator rhizome under mild low temperature stress in the root zone. Northwest Bot. 2019, 39, 9. [Google Scholar]
- Zhao, H.; Hu, W.; Sha, H.; Jia, Y.; Tian, X. Regulatory effects of proline and γ-aminobutyric acid on the antioxidant system of rice under salt stress. J. Northeast. Agric. Univ. 2017, 48, 9. [Google Scholar]
- Hu, W. Regulatory effects of salicylic acid, proline and γ-aminobutyric acid on the antioxidant system of rice under salt stress. Northeast. Agric. Univ. 2018, 4, 1–64. [Google Scholar]
- Wang, X.; Xie, B.; Li, J.; Duan, L. Effects of exogenous γ-aminobutyric acid (GABA) on waterlogging tolerance of wheat seedlings. North China Agric. J. 2010, 25, 155–160. [Google Scholar]
- Xu, J.; Liu, T.; Yang, S.; Jin, X.; Qu, F.; Huang, N.; Hu, X. Polyamines are involved in GABA-regulated salinity-alkalinity stress tolerance in muskmelon. Environ. Exp. Bot. 2019, 164, 181–189. [Google Scholar] [CrossRef]
- Hong, S.; Xiaolong, Y.; Shaoling, Z. Simultaneous Analysis of Organic Acids and Inorganic Anions in Common Bean by Ion Chromatography. J. South China Agric. Univ. 2001, 2, 8–10. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.Y.; Guo, Y.J.; Zhang, X.Q.; Gao, H.B.; Gao, X.M.; Hao, L.Y.; Wd YA, N.G. Effect of γ-aminobutyric Acid on Growth and Mineral Elements Contents in Cucumber Seedlings Under Different NaCl Concentration. North. Hortic. 2014, 3, 5–8. [Google Scholar]
- Li, W.; Liu, J.; Ashraf, U.; Li, G.; Li, Y.; Lu, W.; Hu, J. Exogenous γ-aminobutyric Acid (GABA) Application Improved Early Growth, Net Photosynthesis, and Associated Physio-Biochemical Events in Maize. Front. Plant Sci. 2016, 7, 919. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Yuan, F.; Guo, J.; Han, G.; Wang, C.; Chen, M.; Wang, B. Current Understanding of Role of Vesicular Transport in Salt Secretion by Salt Glands in Recretohalophytes. Int. J. Mol. Sci. 2021, 22, 2203. [Google Scholar] [CrossRef]
- Daridon, A.; Sequeira, M.; Pennarun-Thomas, G.; Dirac, H.; Krog, J.P.; Gravesen, P.; de Rooij, N.F. Chemical sensing using an integrated microfluidic system based on the Berthelot reaction. Sens. Actuators B Chem. 2001, 76, 235–243. [Google Scholar] [CrossRef]
- Rezaei-Chiyaneh, E.; Seyyedi, S.M.; Ebrahimian, E.; Moghaddam, S.S.; Damalas, C.A. Exogenous application of gamma-aminobutyric acid (GABA) alleviates the effect of water deficit stress in black cumin (Nigella sativa L.). Ind. Crops Prod. 2018, 112, 741–748. [Google Scholar] [CrossRef]
- Gong, B.; Wen, D.; VandenLangenberg, K.; Wei, M.; Yang, F.; Shi, Q.; Wang, X. Comparative effects of NaCl and NaHCO3 stress on photosynthetic parameters, nutrient metabolism, and the antioxidant system in tomato leaves. Entia Hortic. 2013, 157, 1–12. [Google Scholar] [CrossRef]
- Adolf, V.I.; Jacobsen, S.E.; Shabala, S. Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Fang, S.; Hou, X.; Liang, X. Response Mechanisms of Plants Under Saline-Alkali Stress. Front. Plant Sci. 2021, 12, 667458. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Liu, T.; Xu, J.; Gao, Z.; Hu, X. Exogenous GABA enhances muskmelon tolerance to salinity-alkalinity stress by regulating redox balance and chlorophyll biosynthesis. BMC Plant Biol. 2019, 19, 48. [Google Scholar] [CrossRef] [PubMed]
- Kinnersley, A.M.; Turano, F.J. Gamma Aminobutyric Acid (GABA) and Plant Responses to Stress. Crit. Rev. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Zhang, D.; Zhang, Z.; Xing, Y.; Sun, N.; Wang, S.; Cai, H. Exogenous Application of GABA Alleviates Alkali Damage in Alfalfa by Increasing the Activities of Antioxidant Enzymes. Agronomy 2022, 12, 1577. https://doi.org/10.3390/agronomy12071577
Li D, Zhang D, Zhang Z, Xing Y, Sun N, Wang S, Cai H. Exogenous Application of GABA Alleviates Alkali Damage in Alfalfa by Increasing the Activities of Antioxidant Enzymes. Agronomy. 2022; 12(7):1577. https://doi.org/10.3390/agronomy12071577
Chicago/Turabian StyleLi, Donghuan, Depeng Zhang, Zizhao Zhang, Yimei Xing, Na Sun, Shuo Wang, and Hua Cai. 2022. "Exogenous Application of GABA Alleviates Alkali Damage in Alfalfa by Increasing the Activities of Antioxidant Enzymes" Agronomy 12, no. 7: 1577. https://doi.org/10.3390/agronomy12071577