
Hybrid Vigour and Hybrid Mimics in Japonica Rice

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
- Selection for hybrid mimics
- Field evaluation
- Real-time qPCR
- Q2 gas-exchange photosynthetic measurement
3. Results
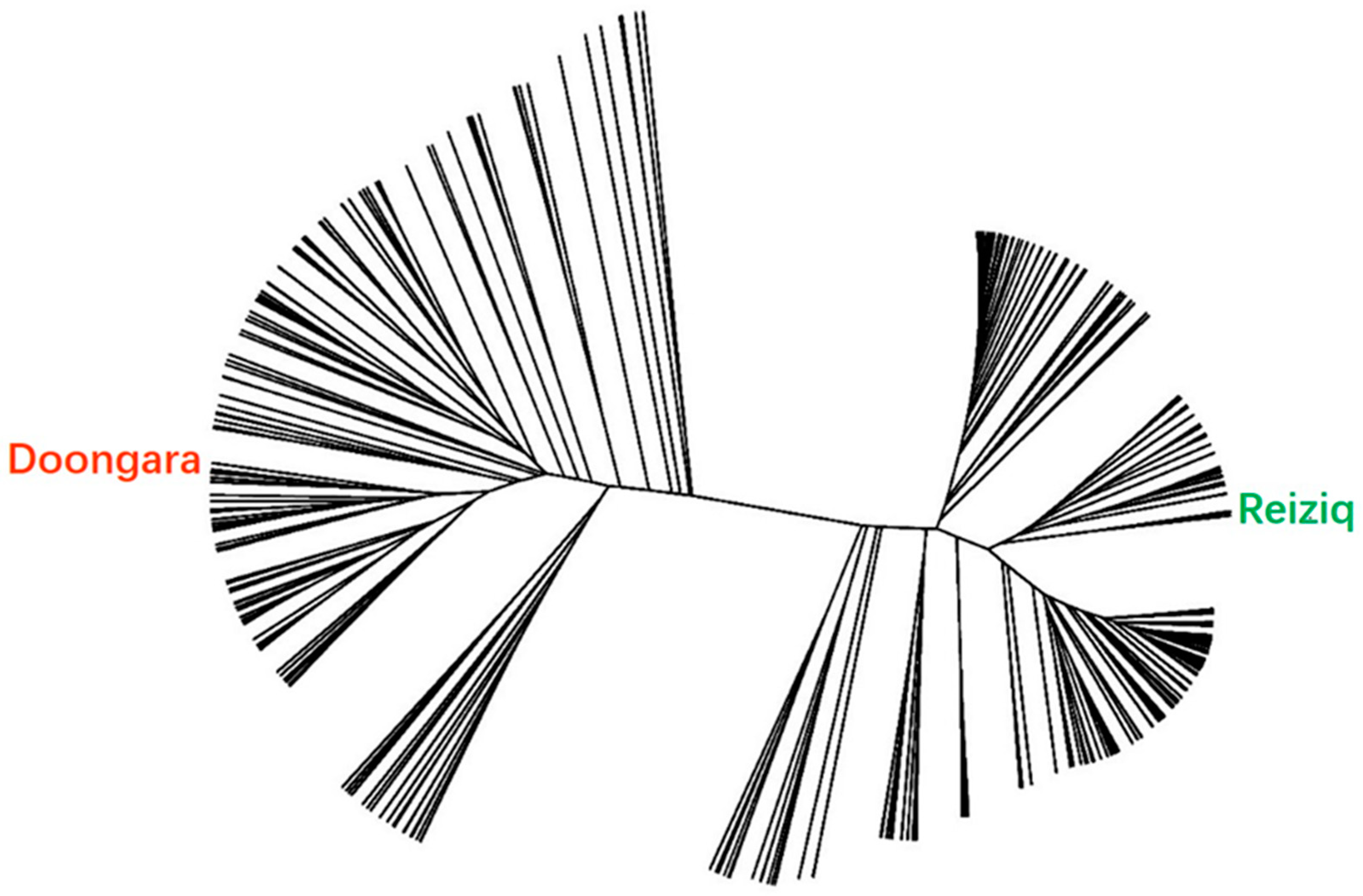
3.1. Selection of Japonica Rice Hybrids for Development of Hybrid Mimics
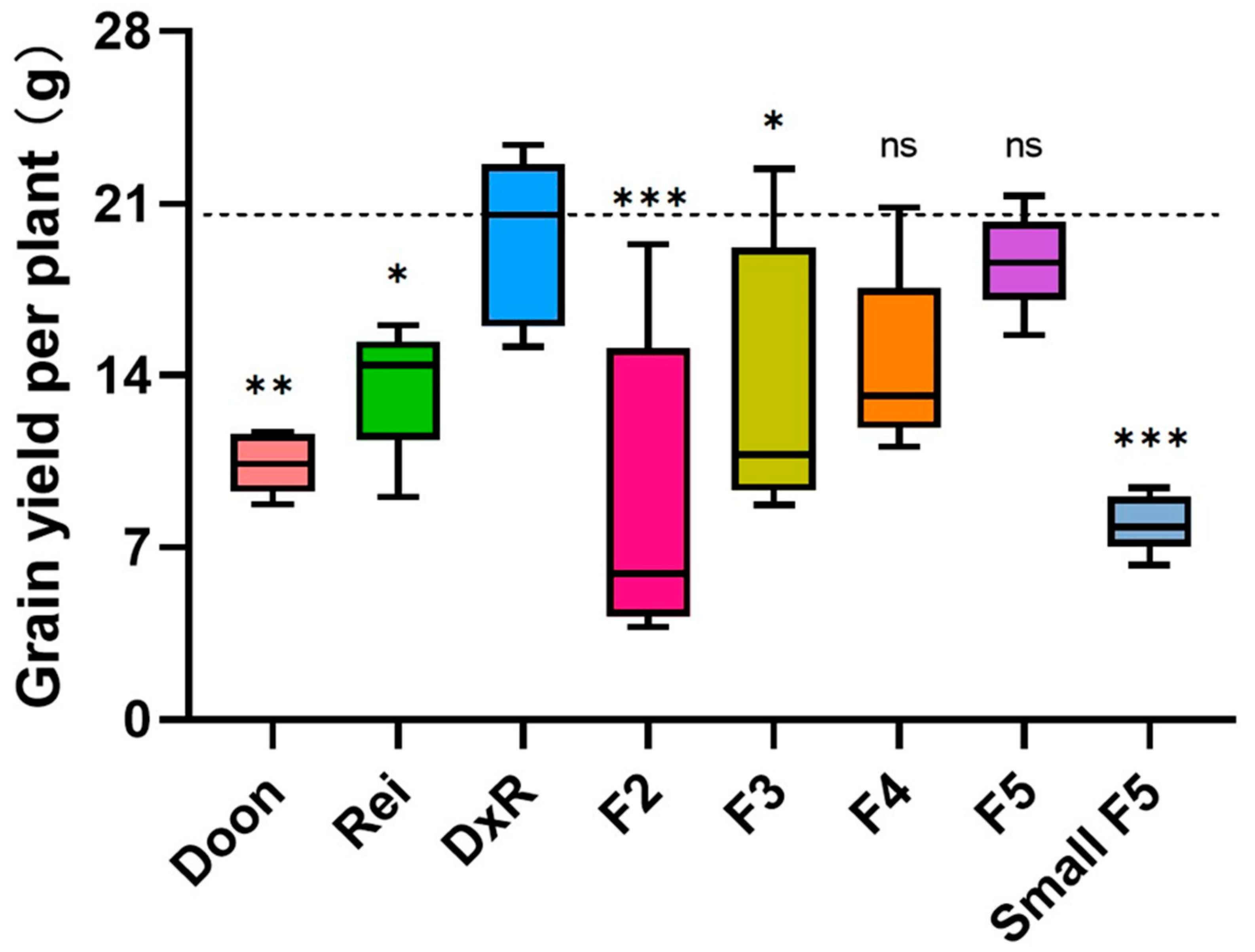
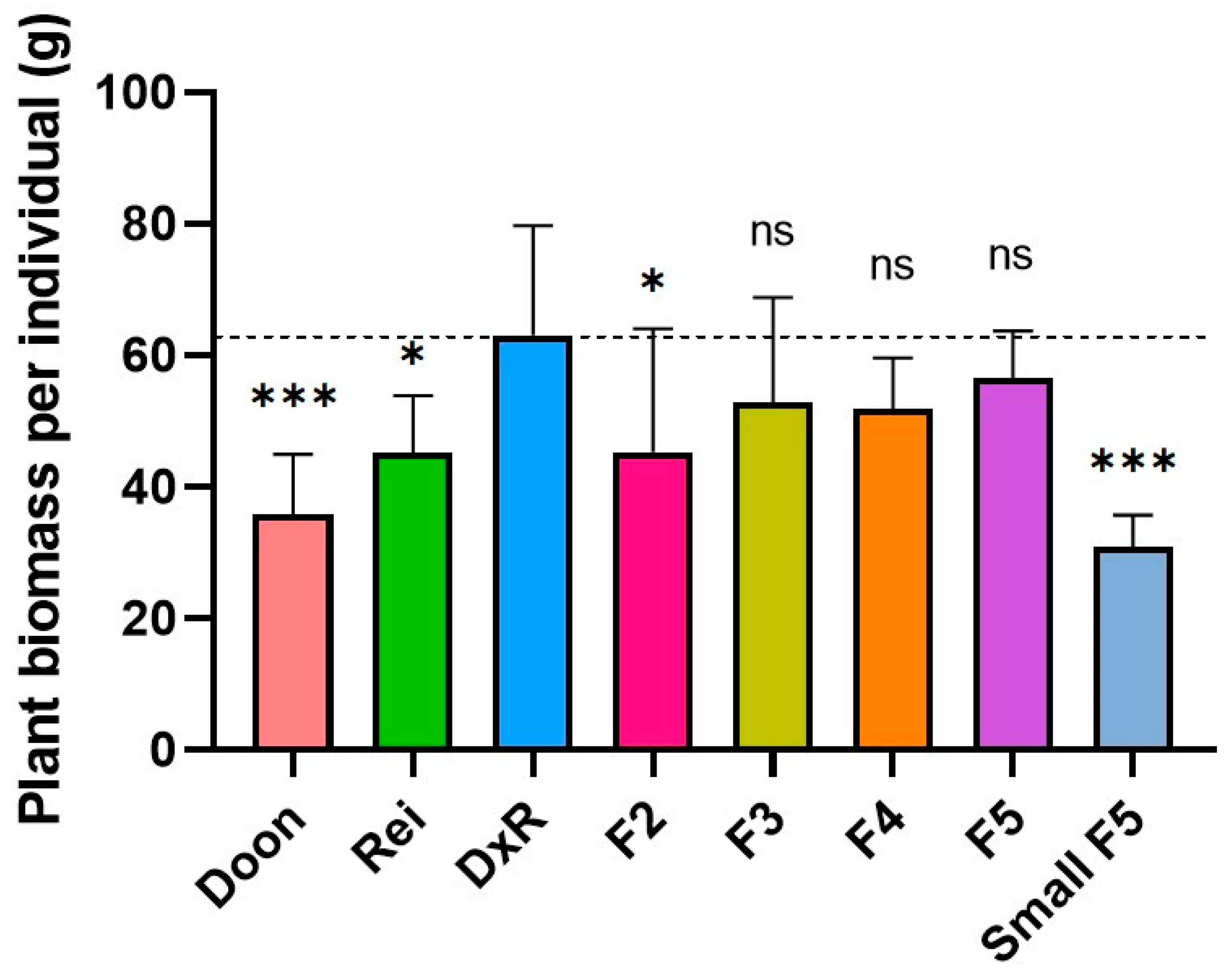
3.2. Hybrid Mimic Selection from a Japonica Rice Hybrid
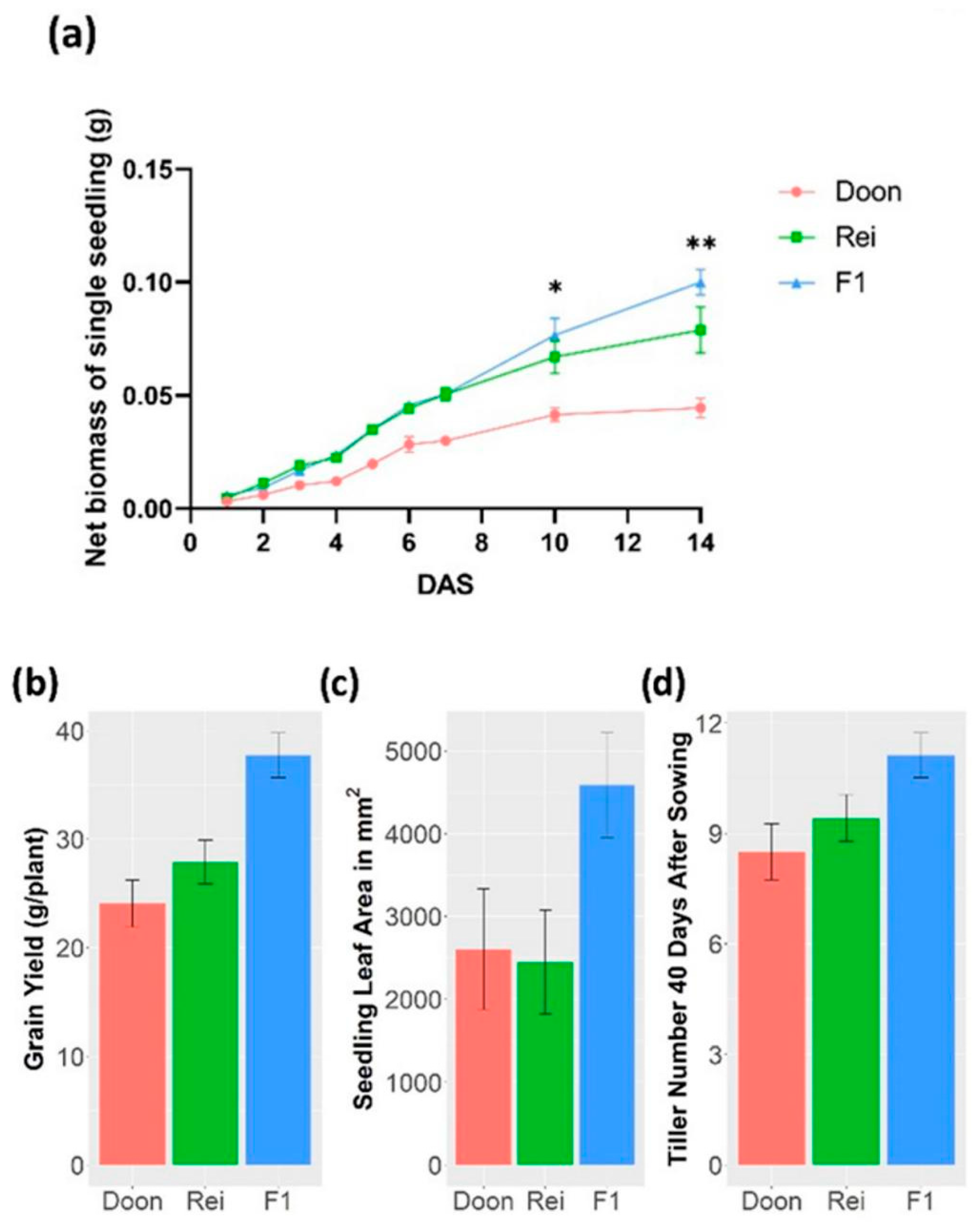
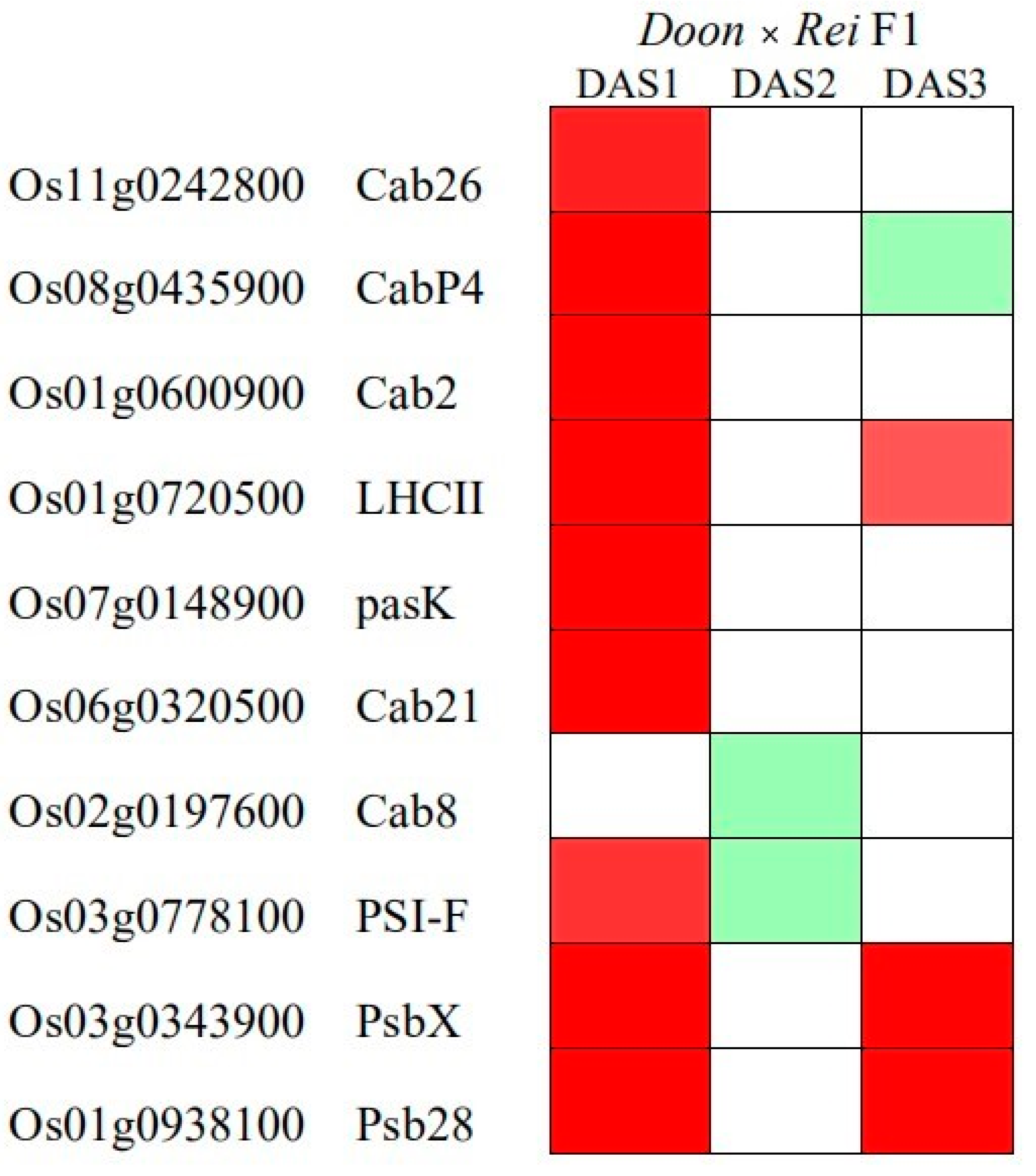
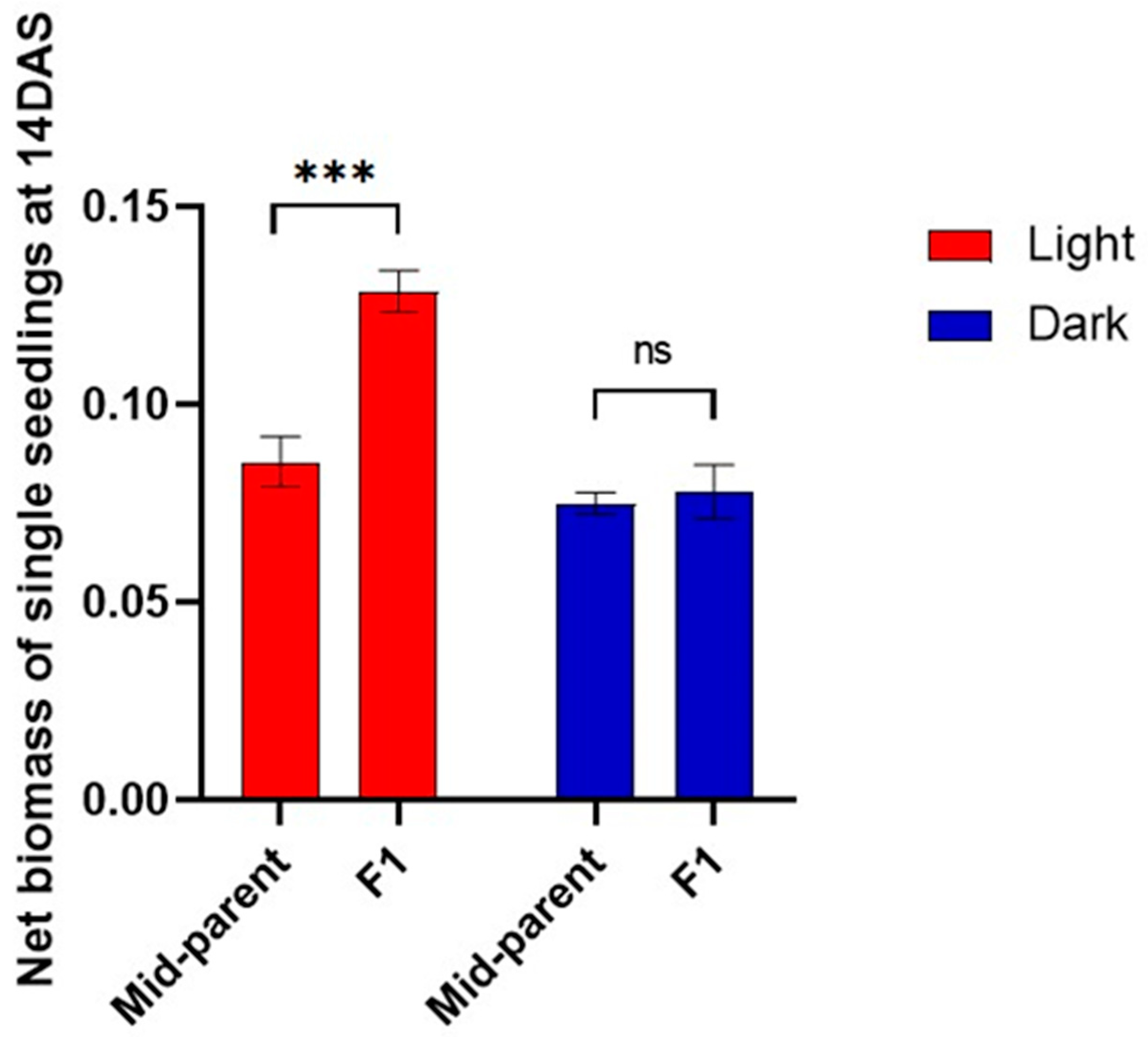
3.3. Heterosis of the F1 Hybrid Started in Early Development
3.4. Dark Germination Eliminated Hybrid Vigour
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shull, G.H. What Is “Heterosis”? Genetics 1948, 33, 439–446. [Google Scholar] [CrossRef]
- Fischer, R.; Byerlee, D.; Edmeades, G. Crop Yields and Global Food Security; ACT ACIAR: Canberra, Australia, 2014.
- Chen, Z. Genomic and epigenetic insights into the molecular bases of heterosis. Nat. Rev. Genet. 2013, 14, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Davenport, C.B. Degeneration, Albinism and Inbreeding. Science 1908, 28, 454–455. [Google Scholar] [CrossRef] [PubMed]
- East, E.M. Inbreeding in Corn. In Reports of the Connecticut Agricultural Experiments Station for 1907; University of Illinois Urbana-Champaign: Urbana, IL, USA; Champaign, IL, USA, 1908; pp. 419–428. [Google Scholar]
- Shull, G.H. The Composition of a Field of Maize; American Breeders Association: Philadelphia, PA, USA, 1908; pp. 296–301. [Google Scholar]
- Xiao, J.; Li, J.; Yuan, L.; Tanksley, S.D. Dominance is the major genetic basis of heterosis in rice as revealed by QTL analysis using molecular markers. Genetics 1995, 140, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Cockerham, C.C.; Zeng, Z.B. Design III with marker loci. Genetics 1996, 143, 1437–1456. [Google Scholar] [CrossRef]
- Stuber, C.W.; Lincoln, S.E.; Wolff, D.W.; Helentjaris, T.; Lander, E.S. Identification of genetic factors contributing to heterosis in a hybrid from two elite maize inbred lines using molecular markers. Genetics 1992, 132, 823–839. [Google Scholar] [CrossRef]
- Li, Z.K.; Luo, L.J.; Mei, H.W.; Wang, D.L.; Shu, Q.Y. Overdominant epistatic loci are the primary genetic basis of inbreeding depression and heterosis in rice. I. Biomass and grain yield. Genetics 2001, 153, 1737–1753. [Google Scholar] [CrossRef]
- Luo, L.J.; Li, Z.K.; Mei, H.W.; Shu, Q.Y.; Tabien, R. Overdominant epistatic loci are the primary genetic basis of inbreeding depression and heterosis in rice. II. Grain yield components. Genetic 2001, 158, 1755–1771. [Google Scholar] [CrossRef]
- Crow, J.F. The rise and fall of overdominance. Plant Breed 2000, 17, 225–257. [Google Scholar]
- Lippman, Z.B.; Zamir, D. Heterosis: Revisiting the magic. Trends Genet. 2007, 23, 60–66. [Google Scholar] [CrossRef]
- Schnell, F.W.; Cockerham, C.C. Multiplicative vs. arbitrary gene action in heterosis. Genetics 1992, 131, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Greaves, I.K.; Groszmann, M.; Wu, L.M.; Dennis, E.S.; Peacock, W.J. Hybrid mimics and hybrid vigour in Arabidopsis. Proc. Natl. Acad. Sci. USA 2015, 112, E4959–E4967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johns, M.A.; Mao, L. Differentiation of the two rice subspecies indica and japonica: A Gene Ontology perspective. Funct. Integr. Genom. 2007, 7, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Yuan, Q.; Pei, X.; Wang, F.; Hu, N.; Yao, K.; Wang, Z. Rice transgene flow: Its patterns, model and risk management. Plant Biotechnol. J. 2014, 12, 1259–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yuan, L. Hybrid rice: Genetics, Breeding, and Seed Production. Plant Breed. Rev. 2000, 17, 15–142. [Google Scholar]
- Perez-Prat, E.; van Lookeren Campagne, M.M. Hybrid seed production and the challenge of propagating male-sterile plants. Trends Plant Sci. 2002, 5, 199–203. [Google Scholar] [CrossRef]
- Guzman, C.D.; Oard, J. Genetics and Breeding System for Cytoplasmic and Genetic Male Sterility in Rice. In Protecting Rice Grains in the Post-Genomic Era; Jia, Y., Ed.; IntechOpen: London, UK, 2009. [Google Scholar]
- Yuan, L. Hybrid Rice Breeding for Super High Yield. In China and IRRI: Improving China’s Rice Productivity in the 21st Century; Denning, G.L., Mew, T.W., Eds.; International Rice Research Institute: Manila, Philippines, 1998; pp. 10–12. [Google Scholar]
- Pu, H.; Zhou, Z.; Xu, D. Development history and problems of three-line hybrid japonica rice. J. Jiangsu Agric. Sci. 2015, 43, 74–77. [Google Scholar]
- Deng, H. Japonica Hybrid Rice in China, 1st ed.; China Agriculture Press: Beijing, China, 2008. [Google Scholar]
- Deng, J.; Wang, A.; Tu, X. Planting performance and high-yield cultivation techniques of two-line hybrid japonica rice variety Feng-Liang-You No.1. Anhui Agric. Sci. 2006, 34, 6161–6163. [Google Scholar]
- Pan, Y. Characteristics and high-yield cultivation techniques of super rice D-You-527. China Seed Ind. 2006, 2, 23. [Google Scholar]
- He, Y.; Zhang, Y.; Liao, Y.; Dennis, E.S.; Peacock, W.J.; Wu, X. Rice hybrid mimics have stable yields equivalent to those of the F1 hybrid and suggest a basis for hybrid vigour. Planta 2021, 254, 51. [Google Scholar] [CrossRef]
- Zhu, A.; Wang, A.; Zhang, Y.; Dennis, E.S.; Peacock, W.J.; Greaves, I.K. Early Establishment of Photosynthesis and Auxin Biosynthesis Plays a Key Role in Early Biomass Heterosis in Brassica napus (Canola) Hybrids. Plant Cell Physiol. 2020, 61, 1134–1143. [Google Scholar] [CrossRef]
- Scafaro, A.P.; Negrini, A.A.; O’Leary, B.; Rashid, F.A.; Hayes, L.; Fan, Y.; Zhang, Y.; Chochois, V.; Badger, M.R.; Millar, A.H.; et al. The combination of gas-phase fluorophore technology and automation to enable high-throughput analysis of plant respiration. Plant Methods 2017, 13, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtois, B.; Audebert, A.; Dardou, A.; Roques, S.; Ghneim- Herrera, T.; Droc, G.; Frouin, J.; Rouan, L.; Goze, E.; Kilian, A.; et al. Genome-Wide Association Mapping of Root Traits in a Japonica Rice Panel. PLoS ONE 2003, 8, e78037. [Google Scholar] [CrossRef] [Green Version]
- Murtagh, F.; Legendre, P. Ward’s hierarchical agglomerative clustering method: Which algorithms implement Ward’s criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Schliep, K. An environmrnt for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- McDonald, D.J. Temperate rice technology for the 21st century: An Australian example. Aust. J. Exp. Agric. 1994, 34, 877–888. [Google Scholar] [CrossRef]
- Lewin, L.; Reinke, R.; Snell, P. Registration and description of rice variety Reiziq. Plant Var. J. 2006, 19, 138–142. [Google Scholar]
- Hornbuckle, J.; Christen, E. Physical Properties of Soils in the Murrumbidgee and Coleambally Irrigation Areas; CSIRO Land and Water: Griffith, NSW, Australia, 1999. [Google Scholar]
- Coombes, N.E. The Reactive Tabu Search for Efficient Correlated Experimental Designs. Ph.D. Thesis, Liverpool John Moores University, Liverpool, UK, 2002. [Google Scholar]
- Reinke, R.F. Genetic Improvement of Seedling Vigour in Temperate Rice. Ph.D. Thesis, Australian National University, Canberra, ACT, Australia, 2000. [Google Scholar]
- Butler, D.; Cullis, B.R.; Gilmour, A.R.; Gogel, B.J. ASReml-R Reference Manual; Department of Primary Industries and Fisheries: Brisbane, QLD, Australia, 2009.
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.L.; Moorman, A.F.M. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.B.; Moorman, A.F.M. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [Green Version]
- Meyer, R.C.; Törjék, O.; Becher, M.; Altmann, T. Heterosis of Biomass Production in Arabidopsis Establishment during Early Development. Plant Physiol. 2004, 134, 1813–1823. [Google Scholar] [CrossRef] [Green Version]
- Paschold, A.; Marcon, C.; Hoecker, N.; Hochholdinger, F. Molecular dissection of heterosis manifestation during early maize root development. Theor. Appl. Genet. 2010, 120, 383–388. [Google Scholar] [CrossRef]
- Liu, P.; Peacock, W.J.; Wang, L.; Furbank, R.; Larkum, A.; Dennis, E.S. Leaf growth in early development is key to biomass heterosis in Arabidopsis. J. Exp. Bot. 2020, 71, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Campbell, N.; Reece, J. Biology, 7th ed.; Pearson: London, UK, 2004. [Google Scholar]
- Saeki, N.; Kawanabe, T.; Ying, H.; Shimizu, M.; Kojima, M.; Abe, H.; Okazaki, K.; Kaji, M.; Taylor, J.M.; Sakakibara, H.; et al. Molecular and cellular characteristics of hybrid vigour in a commercial hybrid of Chinese cabbage. BMC Plant Biol. 2016, 16, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckardt, N.A. From Darkness into Light: Factors Controlling Photomorphogenesis. Plant Cell 2001, 13, 219–221. [Google Scholar] [CrossRef]
- Kawano, K.; Tanaka, A. Vegetative vigour in relation to yield and nitrogen response in the rice plant. Jpn. J. Breed. 1969, 19, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Kawano, K.; Kurosawa, K.; Takahashi, M. Heterosis in vegetative growth of the rice plant. Jpn. J. Breed. 1969, 19, 335–342. [Google Scholar] [CrossRef]
- Blanco, L.; Akita, S.; Virmani, S. Growth and yield of F1 rice hybrids in different levels of nitrogen. Jpn. J. Crop Sci. 1986, 19, 12–13. [Google Scholar]
- Akita, S. Physiological Bases of Heterosis in Rice. In Hybrid Rice; International Rice Research Institute: Manila, Philippines, 1986. [Google Scholar]
- Li, C.C.; Rutger, J.N. Inheritance of cool-temperature seedling vigor in rice and its relationship with other agronomic characters. Crop Sci. 1980, 20, 295–298. [Google Scholar] [CrossRef]
- Khush, G.S. Rice breeding: Past, present and future. J. Genet. 1987, 66, 195–216. [Google Scholar] [CrossRef]
- Yuan, L. Development of hybrid rice to ensure food security. Rice Sci. 2014, 21, 1–2. [Google Scholar] [CrossRef]
- Zheng, W.; Ma, Z.; Zhao, M.; Xiao, M.; Zhao, J.; Wang, C.; Gao, H.; Bai, Y.; Wang, H.; Sui, G. Research and Development Strategies for Hybrid japonica Rice. Rice 2020, 13, 36. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Photosynthesis Starting Time (HAS) |
|---|---|
| Doongara | 47.49 ± 1.39 |
| Reiziq | 49.51 ± 5.51 |
| MPV | 48.5 ± 2.66 |
| F1 | 35.25 ± 3.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Ovenden, B.; He, Y.; Ye, W.; Wu, X.; Peacock, W.J.; Dennis, E.S. Hybrid Vigour and Hybrid Mimics in Japonica Rice. Agronomy 2022, 12, 1559. https://doi.org/10.3390/agronomy12071559
Zhang Y, Ovenden B, He Y, Ye W, Wu X, Peacock WJ, Dennis ES. Hybrid Vigour and Hybrid Mimics in Japonica Rice. Agronomy. 2022; 12(7):1559. https://doi.org/10.3390/agronomy12071559
Chicago/Turabian StyleZhang, You, Ben Ovenden, Yao He, Wenwei Ye, Xianjun Wu, William James Peacock, and Elizabeth S. Dennis. 2022. "Hybrid Vigour and Hybrid Mimics in Japonica Rice" Agronomy 12, no. 7: 1559. https://doi.org/10.3390/agronomy12071559