Effect of Foliar Treatment with Aqueous Dispersions of Silver Nanoparticles on Legume-Rhizobium Symbiosis and Yield of Soybean (Glycine max L. Merr.)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Preparation of Silver NPs Dispersions
2.3. UV-Visible Spectrophotometry
2.4. Dynamic Light Scattering (DLS)
2.5. Transmission Electron Microscopy (TEM)
2.6. Powder X-ray Diffraction
2.7. Field Experiment on the Biological Effectiveness of Silver NPs Dispersions
- Control—N0P0 (without additional application of fertilizers—mineral nitrogen and phosphorus), no treatment with Ag-PHMB;
- Background—N30P40 (additional application of fertilizers—30 kg/ha of mineral nitrogen and 40 kg/ha of mineral P2O5; the sources of mineral nitrogen and P2O5 were NH4NO3 and NH4H2PO4), no treatment with Ag-PHMB;
- Background + Ag-PHMB—seed treatment (25 mL/ton, working solution consumption 10 L/ton) + plant spraying: 1st—in the phase of 3rd trifoliate (V3 phase), 2nd—in the phase of beginning bloom (R1 phase) (40 mL/ha, working solution consumption 300 L/ha);
- Background + Ag-PHMB—seed treatment (50 mL/ton, working solution consumption 10 L/ton) + plant spraying: 1st—in the phase of 3rd trifoliate (V3 phase), 2nd—in the phase of beginning bloom (R1 phase) (80 mL/ha, working solution consumption 300 L/ha);
- Background + Ag-PHMB—seed treatment (75 mL/ton, working solution consumption 10 L/ton) + plant spraying: 1st—in the phase of 3rd trifoliate (V3 phase), 2nd—in the phase of beginning bloom (R1 phase) (120 mL/ha, working solution consumption 300 L/ha);
- Background + PHMB (pure stabilizer without silver NPs)—seed treatment (75 mL/ton, working solution consumption 10 L/ton) + plant spraying: 1st—in the phase of 3rd trifoliate (V3 phase), 2nd—in the phase of beginning bloom (R1 phase) (120 mL/ha, working solution consumption 300 L/ha).
2.8. Determination of the Specific Activity of Peroxidase in Soybean Biomass
2.9. Determination of the Specific Activity of Polyphenol Oxidase in Soybean Biomass
2.10. Determination of Residual Amounts of Silver in Biomass
2.11. Statistical Processing of Results
3. Results and Discussion
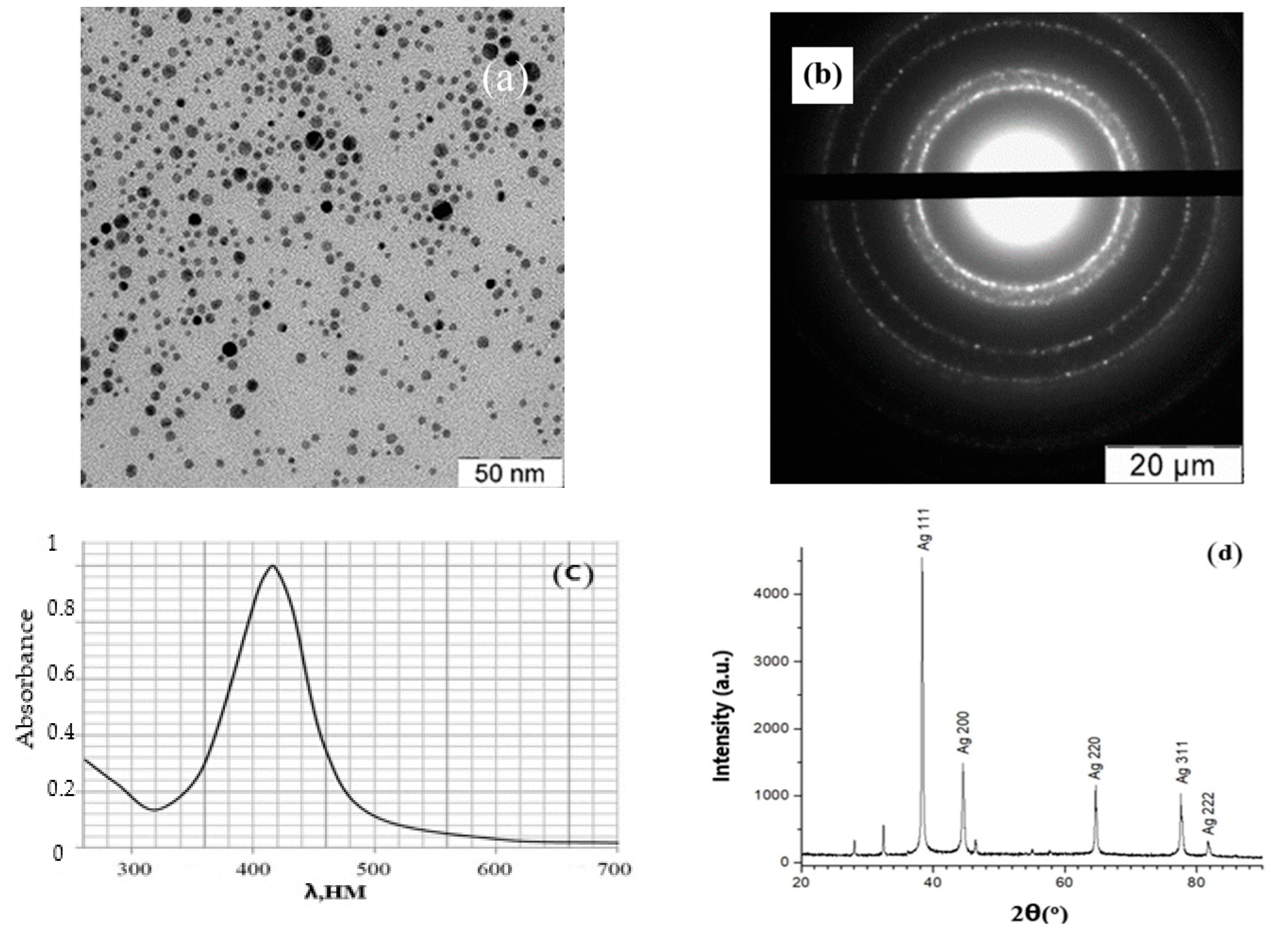
3.1. Synthesis of Aqueous Dispersions of Silver NPs Stabilized with PHMB
3.2. Field Experiments
3.2.1. Weather Conditions
3.2.2. Effect of Different Seed Treatments
3.3. Determination of Residual Amounts of Silver in Plant Biomass
4. Conclusions
- (a)
- Effects associated with blocking the ethylene-dependent signaling system, and consisting mainly in stimulating vegetation, root and shoot growth, biomass accumulation and slowing down maturation and wilting processes;
- (b)
- Effects associated with oxidative stress, consisting in disruption of normal cell activity, on the one hand, and stimulation of resistance to adverse external influences, including infection with phytopathogens, on the other hand.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kalaitzandonakes, N.G.; Kaufman, J.D.; Zahringer, K.A. The Economics of Soybean Disease Control; CABI: Wallingford, UK, 2019; 220p. [Google Scholar]
- George, B. Oilseeds World Trade Markets and Trade; USDA: Washington, DC, USA, 2022; p. 15. Available online: https://usda.library.cornell.edu/concern/publications/tx31qh68h?locale=en (accessed on 23 March 2022).
- Singh, G. The Soybean: Botany, Production and Uses; CABI: Wallingford, UK, 2010; 512p. [Google Scholar]
- DiCenzo, G.C.; Zamani, M.; Checcucci, A.; Fondi, M.; Griffitts, J.S.; Finan, T.M.; Mengoni, A. Multidisciplinary approaches for studying rhizobium-legume symbioses. Can. J. Microbiol. 2019, 65, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, L.; Lesueur, D. Challenges of formulation and quality of biofertilizers for successful inoculation. Appl. Microbiol. Biotechnol. 2013, 97, 8859–8873. [Google Scholar] [CrossRef] [PubMed]
- Glyan’ko, A. Signaling Systems of Rhizobia (Rhizobiaceae) and Leguminous Plants (Fabaceae) upon the Formation of a Legume-Rhizobium Symbiosis (Review). Appl. Biochem. Microbiol. 2015, 51, 494–504. [Google Scholar] [CrossRef]
- Mangialardi, J.P.; Orlowski, J.M.; Lawrence, B.H.; Bond, J.A.; Golden, B.R.; Catchot, A.; Peeples, J.D.; Eubank, T.W. Growth regulation with lactofen does not affect seed yield of irrigated soybean. Agron. J. 2016, 108, 1112–1115. [Google Scholar] [CrossRef] [Green Version]
- Ariyoshi, Y.; Itoyama, H.; Nakagawa, A.; Nobuyuki, A.; Yukari, K.; Yuki, T.; Seiya, T.; Megumi, N.; Kanji, T.; Mari, I.I.; et al. Regulation of brassinosteroid on pod growth through cell hypertrophy in soybean (Glycine max (L.) Merr.). Plant Growth Regul. 2016, 80, 391–395. [Google Scholar] [CrossRef]
- Bayat, M.; Zargar, M.; Astarkhanova, T.; Pakina, E.; Ladan, S.; Lyashko, M.; Shkurkin, S.I. Facile Biogenic Synthesis and Characterization of Seven Metal-Based Nanoparticles Conjugated with Phytochemical Bioactives Using Fragaria ananassa Leaf Extract. Molecules 2021, 26, 3025. [Google Scholar] [CrossRef]
- Nowack, B.; Krug, H.F.; Height, M. 120 Years of Nanosilver History: Implications for Policy Makers. Environ. Sci. Technol. 2011, 45, 1177–1183. [Google Scholar] [CrossRef]
- Calderón-Jiménez, B.; Johnson, M.E.; Montoro, B.A.R.; Murphy, K.E.; Winchester, M.R.; Baudrit, V.J.R. Silver Nanoparticles: Technological Advances, Societal Impacts, and Metrological Challenges. Front. Chem. 2017, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Rajwade, J.M.; Paknikar, K.M.; Kumbhar, J.V. Applications of bacterial cellulose and its composites in biomedicine. Appl. Microbiol. Biotechnol. 2015, 99, 2491–2511. [Google Scholar] [CrossRef]
- Bayat, M.; Zargar, M.; Chudinova, E.; Astarkhanova, T.; Pakina, E. In Vitro Evaluation of Antibacterial and Antifungal Activity of Biogenic Silver and Copper Nanoparticles: The First Report of Applying Biogenic Nanoparticles against Pilidium concavum and Pestalotia sp. Fungi. Molecules 2021, 26, 5402. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. Impacts of Silver Nanoparticles on Plants: A Focus on the Phytotoxicity and Underlying Mechanism. Int. J. Mol. Sci. 2019, 20, 1003. [Google Scholar] [CrossRef] [PubMed]
- Bayat, M.; Zargar, M.; Murtazova, K.M.-S.; Nakhaev, M.R.; Shkurkin, S.I. Ameliorating Seed Germination and Seedling Growth of Nano-Primed Wheat and Flax Seeds Using Seven Biogenic Metal-Based Nanoparticles. Agronomy 2022, 12, 811. [Google Scholar] [CrossRef]
- Cunningham, S.; Brennan-Fournet, M.E.; Ledwith, D.; Byrnes, L.; Joshi, L. Effect of Nanoparticle Stabilization and Physicochemical Properties on Exposure Outcome: Acute Toxicity of Silver Nanoparticle Preparations in Zebrafish (Danio rerio). Environ. Sci. Technol. 2013, 47, 3883–3892. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Bandyopadhyay, K.; Ghosh, M.M. Effect of stabilizer concentration on the size of silver nanoparticles synthesized through chemical route. Inorg. Chem. Commun. 2021, 123, 108319. [Google Scholar] [CrossRef]
- Akter, M.; Sikder, M.T.; Rahman, M.M.; Ullah, A.K.M.A.; Hossain, K.F.B.; Banik, S.; Hosokawa, T.; Saito, T.; Kurasaki, M. A systematic review on silver nanoparticles-induced cytotoxicity: Physicochemical properties and perspectives. J. Adv. Res. 2017, 2, 1–16. [Google Scholar]
- Salachna, P.; Byczynska, A.; Zawadzinska, A.; Piechocki, R.; Mizielinska, M. Stimulatory Effect of Silver Nanoparticles on the Growth and Flowering of Potted Oriental Lilies. Agronomy 2019, 9, 610. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Baez, G.A.; Trejo-Tellez, L.I.; Ramirez-Olvera, S.M.; Salinas-Ruiz, J.; Bello-Bello, J.J.; Alcantar-Gonzalez, G.; Hidalgo-Contreras, J.V.; Gómez-Merino, F.C. Silver nanoparticles increase nitrogen, phosphorus, and potassium concentrations in leaves and stimulate root length and number of roots in tomato seedlings in a hormetic manner. Dose-Response 2021, 19, 15593258211044576. [Google Scholar] [CrossRef]
- Gupta, S.D.; Agarwal, A.; Pradhan, S. Phytostimulatory effect of silver nanoparticles (AgNPs) on rice seedling growth: An insight from antioxidative enzyme activities and gene expression patterns. Ecotoxicol. Environ. Saf. 2018, 161, 624–633. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, S.; Srivastava, P.K.; Singh, V.P.; Singh, S.; Prasad, S.M.; Singh, P.K.; Dubey, N.K.; Pandey, A.C.; et al. Nitric oxide alleviates silver nanoparticles (AgNps)-induced phytotoxicity in Pisum sativum seedlings. Plant Physiol. Biochem. 2017, 110, 167–177. [Google Scholar] [CrossRef]
- Quah, B.; Musante, C.; White, J.C.; Ma, X. Phytotoxicity, uptake, and accumulation of silver with different particle sizes and chemical forms. J. Nanopart. Res. 2015, 17, 277. [Google Scholar] [CrossRef]
- Faisal, M.; Saquib, Q.; Alatar, A.A.; Al-Khedhairy, A.A. Phytotoxicity of Nanoparticles, 1st ed.; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Krutyakov, Y.A.; Kudrinsky, A.A.; Gusev, A.A.; Zakharova, O.V.; Klimov, A.I.; Yapryntsev, A.D.; Zherebin, P.M.; Shapoval, O.A.; Lisichkin, G.V. Synthesis of positively charged hybrid PHMB-stabilized silver nanoparticles: The search for a new type of active substances used in plant protection products. Mater. Res. Express 2017, 4, 075018. [Google Scholar] [CrossRef]
- Nichiporovich, A.A. Photosynthetic Activity of Plants in Crops (Methods and Tasks of Accounting in Connection with Yield Potential); USSR Academy of Sciences: Saint Petersburg, Moscow, 1961; pp. 109–119. [Google Scholar]
- Ermakov, A.I.; Arasimovich, V.V.; Smirnova-Ikonnikova, M.I.; Yarosh, N.P.; Lukovnikova, G.A. Methods of biochemical research in plants. Agropromizdat 1987, 2, 430–435. [Google Scholar]
- Dospekhov, B.A. Field Experimentation: Statistical Procedures; Mir Publishers: Moscow, Russia, 1984. [Google Scholar]
- Huynh, K.A.; Chen, K.L. Aggregation Kinetics of Citrate and Polyvinylpyrrolidone Coated Silver Nanoparticles in Monovalent and Divalent Electrolyte Solutions. Environ. Sci. Technol. 2011, 45, 5564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Yang, S.; Tang, F.; Zhu, H. Symbiosis specificity in the legume: Rhizobial mutualism. Cell Microbiol. 2012, 14, 334–342. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, C.; Yang, J.; Yu, N.; Wang, E. Hormone modulation of legume-rhizobial symbiosis. J. Integr. Plant Biol. 2018, 60, 632. [Google Scholar] [CrossRef]
- Murray, J.D. Invasion by Invitation: Rhizobial Infection in Legumes. Mol. Plant-Microbe Interact. 2011, 24, 631. [Google Scholar] [CrossRef] [Green Version]
- Ramongolalaina, C. Dynamics of symbiotic relationship of soybean with Bradyrhizobium diazoefficiens and involvements of root-secreted daidzein behind the continuous cropping. Eur. J. Soil Biol. 2019, 93, 103098. [Google Scholar] [CrossRef]
- Glyan’ko, A.K.; Vasil’eva, G.G. Reactive oxygen and nitrogen species in legume-rhizobial symbiosis: A review. Appl. Biochem. Microbiol. 2010, 46, 15–22. [Google Scholar] [CrossRef]
- Gupta, S.D. Reactive Oxygen Species and Antioxidants in Higher Plants; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Saiz, A.; Peñuelas, M.; Bustos-Sanmamed, P.; Mulet, J.M.; Barja, M.V.; Rouhier, N.; Moore, M.; James, E.K.; Dietz, K.J.; et al. Function of glutathione peroxidases in legume root nodules. J. Exp. Bot. 2015, 66, 2979–2990. [Google Scholar] [CrossRef] [Green Version]
- Arthikala, M.K.; Montiel, J.; Sanchez-Lopez, R.; Noreide, N.; Luis, C.; Carmen. Q. Respiratory Burst Oxidase Homolog Gene A Is Crucial for Rhizobium Infection and Nodule Maturation and Function in Common Bean. Front. Plant Sci. 2017, 8, 2003. [Google Scholar] [CrossRef] [Green Version]
- Glyan’ko, A.K.; Vasil’eva, G.G.; Ischenco, A.A.; Mironova, N.V. The NADPH oxidase activity of pea seedling roots in rhizobial infection depending on abiotic and biotic factors. Appl. Biochem. Microbiol. 2010, 46, 438. [Google Scholar] [CrossRef]
- Glyan’ko, A.K.; Akimova, G.P.; Sokolova, M.G.; Makarova, L.E.; Vasil’eva, G.G. The defense and regulatory mechanisms during development of legume—Rhizobium symbiosis. Appl. Biochem. Microbiol. 2007, 43, 260–267. [Google Scholar] [CrossRef]
- Zakharova, O.V.; Gusev, A.A.; Zherebin, P.M.; Skripnikova, E.V.; Skripnikova, M.K.; Ryzhikh, V.E.; Lisichkin, G.V.; Shapoval, O.A.; Bukovskii, M.E.; Krutyakov, Y.A. Amphopolycarboxyglycinate-Stabilized Silver Nanoparticles Suppress Early and Late Blight of Solanum lycopersicum and Stimulate the Growth of Tomato Plants. Bionanoscience 2017, 7, 692. [Google Scholar] [CrossRef]
- Glyan’ko, A.K.; Ischenko, A.A. Immunity of a leguminous plant infected by nodular bacteria Rhizobium spp. F.: Review. Appl. Biochem. Microbiol. 2017, 53, 140. [Google Scholar] [CrossRef]
- Bhatla, S.C.; Lal, M.A. Plant Physiology, Development and Metabolism; Springer: Singapore, 2018. [Google Scholar] [CrossRef]
- Schaller, G.E.; Binder, B.M. Inhibitors of Ethylene Biosynthesis and Signaling; Humana Press: New York, NY, USA, 2017; p. 223. [Google Scholar] [CrossRef]
- Sisler, E.C.; Serek, M. Inhibitors of ethylene responses in plants at the receptor level: Recent developments. Physiol. Plant 1997, 100, 577. [Google Scholar] [CrossRef]
- Zuverza-Mena, N.; Armendariz, R.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Effects of Silver Nanoparticles on Radish Sprouts: Root Growth Reduction and Modifications in the Nutritional Value. Front. Plant Sci. 2016, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Abd-Alla, M.H.; Nafady, N.A.; Khalaf, D.M. Assessment of silver nanoparticles contamination on faba bean-Rhizobium leguminosarum bv. viciae-Glomus aggregatum symbiosis: Implications for induction of autophagy process in root nodule. Agric. Ecosyst. Environ. 2016, 218, 163. [Google Scholar] [CrossRef]
- Strader, L.C.; Beisner, E.R.; Bartel, B. Silver ions increase auxin efflux independently of effects on ethylene response. Plant Cell 2009, 21, 3585. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Bhatt, D.; Zaidi, M.G.H.; Saradhi, P.P.; Khanna, P.K.; Arora, S. Silver nanoparticle-mediated enhancement in growth and antioxidant status of Brassica juncea. Appl. Biochem. Biotechnol. 2012, 167, 2225. [Google Scholar] [CrossRef]
- Mustafa, G.; Sakata, K.; Hossain, Z.; Komatsu, S. Proteomic study on the effects of silver nanoparticles on soybean under flooding stress. J. Proteom. 2015, 122, 100. [Google Scholar] [CrossRef]
- Mustafa, G.; Sakata, K.; Komatsu, S. Proteomic analysis of soybean root exposed to varying sizes of silver nanoparticles under flooding stress. J. Proteom. 2016, 148, 133. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Mustafa, G.; Nishiuchi, T.; Komatsu, S. Comparative Analysis of the Effect of Inorganic and Organic Chemicals with Silver Nanoparticles on Soybean under Flooding Stress. Int. J. Mol. Sci. 2020, 21, 1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezeshki, S.R.; DeLaune, R.D. Soil Oxidation-Reduction in Wetlands and Its Impact on Plant Functioning. Biology 2012, 1, 196–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burlutskiy, V.A.; Peliy, A.F.; Borodina, E.S.; Diop, A.; Batygin, A.S.; Zargar, M.; Plushchikov, V.G. Efficiency of advanced sprayers for nutrient and pesticide application under precision cultivation of spring rapeseed (Brassica napus). Res. Crops 2020, 21, 466–472. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatments | Average Number of Root Nodules per Plant | Average Weight of Root Nodules, mg per Plant | ||
|---|---|---|---|---|
| Phase R1 | Phase R6 | Phase R1 | Phase R6 | |
| Control-N0P0, no treatment with Ag-PHMB dispersion | 4.8 ± 0.1 | 15 ± 1 | 24 ± 2 | 218 ± 5 |
| Background (N30P40), no treatment with Ag-PHMB dispersion | 4.3 ± 0.2 | 22 ± 1 | 23 ± 2 | 259 ± 6 |
| Background, Ag-PHMB dispersion—seed treatment 25 mL/t + double plant spraying −40 mL/ha | 5.2 ± 0.1 | 30 ± 2 | 34 ± 3 | 568 ± 4 |
| Background, Ag-PHMB dispersion—seed treatment −50 mL/t + double plant spraying −80 mL/ha | 5.2 ± 0.2 | 31 ± 1 | 37 ± 2 | 621 ± 6 |
| Background, Ag-PHMB dispersion—seed treatment −75 mL/t + double plant spraying −120 mL/ha | 5.7 ± 0.2 | 33 ± 2 | 40 ± 3 | 732 ± 6 |
| LSD05 | 0.31 | 0.16 | 9.02 | 6.95 |
| Treatments | Polyphenol Oxidase Activity, [Arbitrary Units (AU) per 1 g of Raw Plant Tissue per Minute] | Peroxidase Activity, [Arbitrary Units (AU) per 1 g of Raw Plant Tissue per Second] | ||
|---|---|---|---|---|
| Roots | Leaves | Roots | Leaves | |
| Control—N0P0, no treatment with Ag-PHMB dispersion | 8.7 ± 0.3 | 5.8 ± 0.1 | 291 ± 9 | 169 ± 3 |
| Background (N30P40), no treatment with Ag-PHMB dispersion | 9.1 ± 0.2 | 5.4 ± 0.2 | 276 ± 7 | 155 ± 2 |
| Background, Ag-PHMB dispersion—seed treatment −25 mL/t + double plant spraying −40 mL/ha | 8.1 ± 0.3 | 6.1 ± 0.1 | 249 ± 8 | 180 ± 5 |
| Background, Ag-PHMB dispersion—seed treatment −50 mL/t + double plant spraying −80 mL/ha | 7.8 ± 0.1 | 6.3 ± 0.3 | 245 ± 8 | 191 ± 7 |
| Background, Ag-PHMB dispersion—seed treatment −75 mL/t + double plant spraying −120 mL/ha | 7.5 ± 0.2 | 6.3 ± 0.1 | 249 ± 5 | 187 ± 7 |
| Treatments | Plant Height, cm | Leaf Area, cm2 | ||
|---|---|---|---|---|
| Phase R1 | Phase R6 | Phase R1 | Phase R6 | |
| Control—N0P0, no treatment with Ag-PHMB dispersion | 68 ± 2 | 83 ± 3 | 535 ± 15 | 585 ± 31 |
| Background (N30P40), no treatment with Ag-PHMB dispersion | 72 ± 3 | 88 ± 2 | 559 ± 18 | 645 ± 37 |
| Background, Ag-PHMB dispersion—seed treatment −25 mL/t + double plant spraying −40 mL/ha | 78 ± 3 | 92 ± 3 | 600 ± 16 | 712 ± 39 |
| Background, Ag-PHMB dispersion—seed treatment −50 mL/t + double plant spraying −80 mL/ha | 81 ± 3 | 101 ± 2 | 638 ± 22 | 781 ± 41 |
| Background, Ag-PHMB dispersion—seed treatment −75 mL/t + double plant spraying −120 mL/ha | 85 ± 3 | 105 ± 3 | 672 ± 24 | 843 ± 42 |
| LSD0.5 | 3.4 | 4.2 | 28.9 | 46.5 |
| Treatment | Yield, kg/ha | Increase to Control, kg/ha | Increase in the Background, kg/ha | |||
|---|---|---|---|---|---|---|
| 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | |
| Control—N0P0, no treatment with Ag-PHMB dispersion | 2100 ± 100 | 2000 ± 100 | - | - | - | - |
| Background (N30P40), no treatment with Ag-PHMB dispersion | 2300 ± 100 | 2200 ± 100 | 200 ± 200 | 200 ± 200 | - | - |
| Background, Ag-PHMB dispersion—seed treatment −25 mL/t + double plant spraying −40 mL/ha | 2600 ± 100 | 2500 ± 100 | 600 ± 200 | 500 ± 200 | 300 ± 200 | 300 ± 200 |
| Background, Ag-PHMB dispersion—seed treatment −50 mL/t + double plant spraying −80 mL/ha | 2700 ± 100 | 2500 ± 100 | 600 ± 200 | 600 ± 20 | 400 ± 200 | 400 ± 200 |
| Background, Ag-PHMB dispersion—seed treatment −75 mL/t + double plant spraying −120 mL/ha | 2700 ± 100 | 2600 ± 100 | 700 ± 200 | 600 ± 200 | 400 ± 200 | 400 ± 200 |
| LSD0.5 | 1.46 | 1.18 | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krutyakov, Y.A.; Mukhina, M.T.; Shapoval, O.A.; Zargar, M. Effect of Foliar Treatment with Aqueous Dispersions of Silver Nanoparticles on Legume-Rhizobium Symbiosis and Yield of Soybean (Glycine max L. Merr.). Agronomy 2022, 12, 1473. https://doi.org/10.3390/agronomy12061473
Krutyakov YA, Mukhina MT, Shapoval OA, Zargar M. Effect of Foliar Treatment with Aqueous Dispersions of Silver Nanoparticles on Legume-Rhizobium Symbiosis and Yield of Soybean (Glycine max L. Merr.). Agronomy. 2022; 12(6):1473. https://doi.org/10.3390/agronomy12061473
Chicago/Turabian StyleKrutyakov, Yurii A., Maria T. Mukhina, Olga A. Shapoval, and Meisam Zargar. 2022. "Effect of Foliar Treatment with Aqueous Dispersions of Silver Nanoparticles on Legume-Rhizobium Symbiosis and Yield of Soybean (Glycine max L. Merr.)" Agronomy 12, no. 6: 1473. https://doi.org/10.3390/agronomy12061473