
Morphological and Physiological Responses of Melia azedarach Seedlings of Different Provenances to Drought Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Determination of Indexes
2.2.1. Determination of Field Water Capacity
2.2.2. Measurement of Seedling Height and Ground Diameter
2.2.3. Determination of Biomass
2.2.4. Leaf Relative Water Content and Leaf Water Saturation Deficit
2.2.5. MDA Content
2.2.6. Superoxide Dismutase Activity
2.2.7. Chlorophyll Content
2.2.8. Photosynthetic Indexes
2.3. Data Statistics and Analysis
2.3.1. Principal Component Analysis
2.3.2. Calculation of Affiliation Function Values
2.3.3. Comprehensive Evaluation of Drought Resistance
3. Results
3.1. Seedling Height Increment and Ground Diameter Increase
3.2. The Biomass and Root–Crown Ratio
3.3. Leaf Water Status
3.4. Response of Photosynthesis and Photosynthetic Pigment
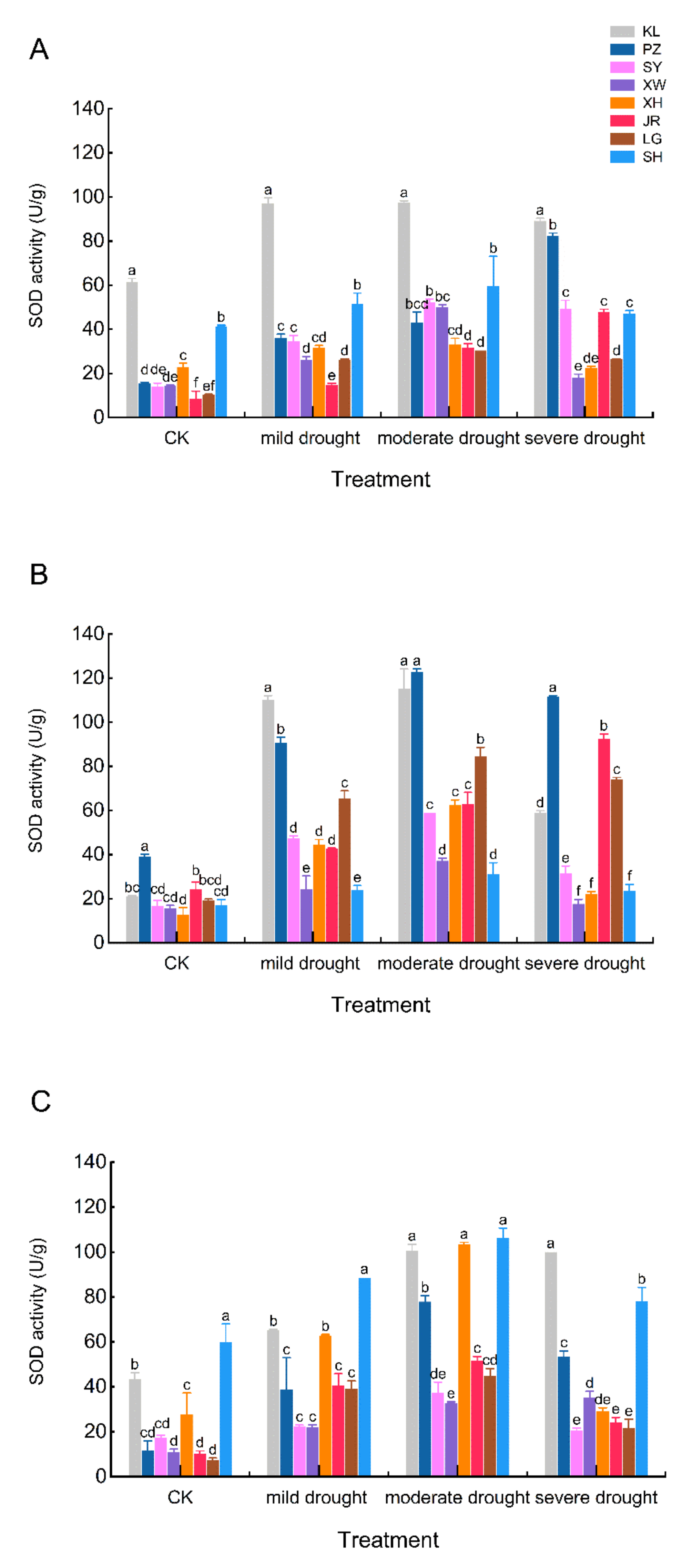
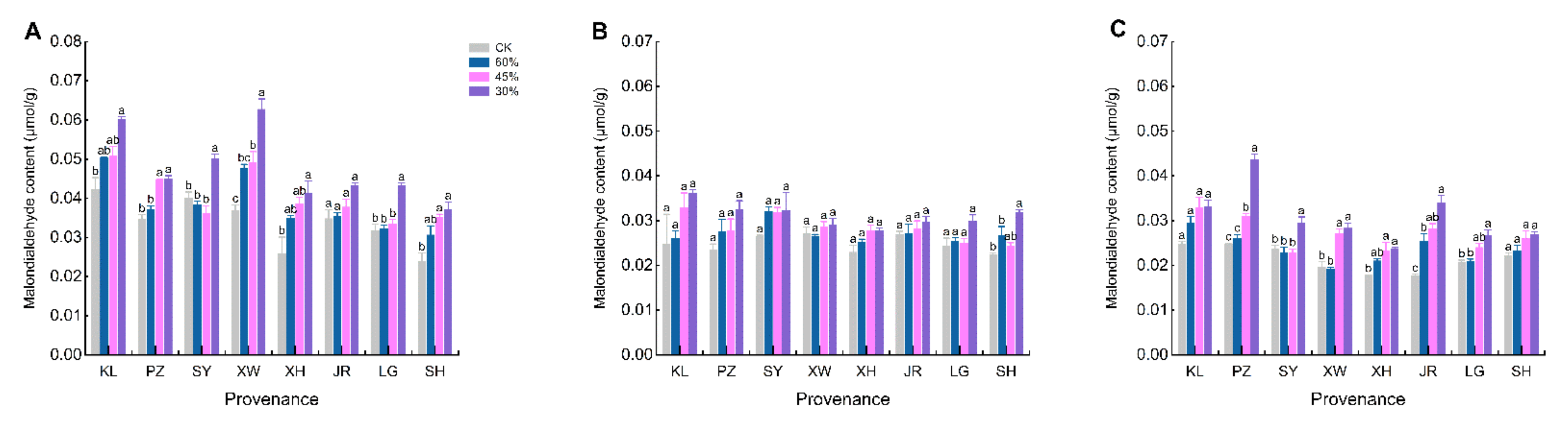
3.5. Response of MDA and SOD
3.6. Comprehensive Evaluation of the Drought Resistance of Melia azedarach in Different Provenances
4. Discussion
4.1. Morphological Growth
4.2. Water Status
4.3. Photosynthesis and Photosynthetic Pigment
4.4. Membrane Lipid Peroxidation and Protective Enzyme Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator | RWC | WSD | ChlA | Tr | Pn | Gs | Ci | MDA | SOD | Seedling Height | Ground Diameter | Root–Crown Ratio |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RWC | 1.000 | |||||||||||
| WSD | 0.142 | 1.000 | ||||||||||
| ChlA | −0.329 | 0.717 | 1.000 | |||||||||
| Tr | −0.349 | −0.354 | 0.075 | 1.000 | ||||||||
| Pn | −0.087 | −0.650 | −0.304 | 0.460 | 1.000 | |||||||
| Gs | −0.510 | −0.324 | 0.007 | 0.601 | 0.199 | 1.000 | ||||||
| Ci | −0.118 | 0.034 | 0.315 | 0.199 | −0.028 | 0.453 | 1.000 | |||||
| MDA | −0.443 | −0.546 | −0.046 | 0.333 | 0.469 | 0.447 | 0.706 | 1.000 | ||||
| SOD | 0.245 | −0.129 | −0.297 | 0.211 | 0.070 | −0.524 | −0.659 | −0.422 | 1.000 | |||
| Seedling height | −0.170 | −0.185 | 0.014 | 0.016 | 0.152 | 0.407 | −0.268 | −0.190 | −0.340 | 1.000 | ||
| Ground diameter | −0.326 | −0.103 | 0.324 | 0.257 | 0.190 | 0.484 | 0.948 | 0.857 | −0.673 | −0.204 | 1.000 | |
| Root–crown ratio | −0.230 | 0.024 | 0.027 | −0.413 | −0.074 | 0.377 | 0.531 | 0.439 | −0.910 | 0.162 | 0.567 | 1.000 |
| Traits | Components | |||
|---|---|---|---|---|
| P1 | P2 | P3 | P4 | |
| Ground diameter | 0.44 | −0.08 | 0.23 | −0.12 |
| MDA | 0.41 | 0.18 | 0.17 | −0.22 |
| Ci | 0.40 | −0.17 | 0.26 | −0.16 |
| SOD | −0.37 | 0.28 | 0.32 | 0.04 |
| Gs | 0.35 | 0.15 | −0.19 | 0.31 |
| Root–crown ratio | 0.32 | −0.25 | −0.36 | −0.25 |
| WSD | −0.12 | −0.52 | 0.16 | 0.23 |
| Pn | 0.13 | 0.45 | −0.02 | −0.08 |
| Tr | 0.15 | 0.38 | 0.32 | 0.40 |
| Chl A | 0.10 | −0.37 | 0.26 | 0.47 |
| Seedling height | 0.04 | 0.08 | −0.63 | 0.36 |
| RWC | −0.24 | −0.07 | 0.01 | −0.43 |
| Provenance | Yik | |||
|---|---|---|---|---|
| Yi1 | Yi2 | Yi3 | Yi4 | |
| KL | 0.84 | 0.37 | 0.93 | 0.57 |
| PZ | 0.90 | 0.17 | 0.92 | 0.36 |
| SY | 1.01 | −0.18 | 0.87 | 0.48 |
| XW | 1.35 | 0.09 | 0.37 | 0.76 |
| XH | 1.26 | 0.36 | 1.20 | 0.54 |
| JR | 1.14 | 0.24 | 1.02 | 0.58 |
| LG | 0.21 | 0.80 | 1.44 | 0.79 |
| SH | 1.68 | −0.17 | 0.31 | 0.34 |
| Indicator | RWC | WSD | ChlA | Tr | Pn | Gs | Ci | MDA | SOD | Seedling Height | Ground Diameter | Root–Crown Ratio |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RWC | 1.000 | |||||||||||
| WSD | 0.105 | 1.000 | ||||||||||
| ChlA | −0.479 | 0.366 | 1.000 | |||||||||
| Tr | −0.412 | −0.372 | 0.005 | 1.000 | ||||||||
| Pn | −0.105 | −0.024 | 0.162 | 0.550 | 1.000 | |||||||
| Gs | −0.413 | −0.283 | −0.048 | 0.929 | 0.600 | 1.000 | ||||||
| Ci | −0.021 | 0.663 | 0.312 | 0.163 | −0.118 | 0.156 | 1.000 | |||||
| MDA | 0.156 | −0.604 | −0.175 | 0.620 | 0.070 | 0.384 | 0.034 | 1.000 | ||||
| SOD | −0.130 | −0.366 | −0.225 | −0.485 | −0.647 | −0.480 | −0.613 | −0.230 | 1.000 | |||
| Seedling height | 0.005 | −0.230 | −0.002 | 0.792 | 0.857 | 0.708 | 0.019 | 0.546 | −0.711 | 1.000 | ||
| Ground diameter | 0.079 | 0.199 | 0.203 | 0.436 | −0.125 | 0.274 | 0.727 | 0.598 | −0.442 | 0.262 | 1.000 | |
| Root–crown ratio | 0.815 | 0.305 | −0.044 | −0.300 | −0.184 | −0.358 | 0.441 | 0.273 | −0.398 | 0.001 | 0.464 | 1.000 |
| Traits | Components | |||
|---|---|---|---|---|
| P1 | P2 | P3 | P4 | |
| Tr | 0.4527 | −0.1419 | −0.0007 | 0.1780 |
| Seedling height | 0.4391 | −0.0490 | 0.0931 | −0.2727 |
| Gs | 0.4193 | −0.1644 | −0.0761 | 0.0629 |
| SOD | −0.3560 | −0.2988 | 0.0972 | 0.2899 |
| Pn | 0.3367 | −0.1160 | −0.1373 | −0.5226 |
| Root–crown ratio | −0.0198 | 0.4834 | 0.3202 | −0.1168 |
| Ci | 0.1406 | 0.4644 | −0.2162 | 0.2286 |
| WSD | −0.0908 | 0.4102 | −0.3861 | −0.1886 |
| Ground diameter | 0.2420 | 0.3623 | 0.0823 | 0.4376 |
| RWC | −0.1024 | 0.2850 | 0.4826 | −0.3560 |
| Chl A | 0.0531 | 0.1321 | −0.4765 | 0.1208 |
| MDA | 0.2928 | 0.0104 | 0.4385 | 0.3135 |
| Provenance | Yik | |||
|---|---|---|---|---|
| Yi1 | Yi2 | Yi3 | Yi4 | |
| KL | 1.002 | 0.778 | 0.607 | 0.993 |
| PZ | 0.072 | 0.885 | 1.040 | 1.185 |
| SY | 0.451 | 1.306 | 0.504 | 0.900 |
| XW | 0.859 | 0.926 | 0.573 | 0.914 |
| XH | 0.621 | 1.308 | 0.873 | 1.134 |
| JR | 0.421 | 0.995 | 0.705 | 1.377 |
| LG | −0.009 | 0.378 | 0.606 | 1.200 |
| SH | 1.374 | 1.809 | 0.651 | 0.375 |
| Indicator | RWC | WSD | ChlA | Tr | Pn | Gs | Ci | MDA | SOD | Seedling Height | Ground Diameter | Root–Crown Ratio |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RWC | 1.000 | |||||||||||
| WSD | 0.051 | 1.000 | ||||||||||
| Chl A | −0.339 | −0.252 | 1.000 | |||||||||
| Tr | 0.332 | −0.575 | 0.145 | 1.000 | ||||||||
| Pn | 0.296 | −0.401 | 0.373 | 0.728 | 1.000 | |||||||
| Gs | 0.325 | −0.259 | −0.222 | 0.864 | 0.610 | 1.000 | ||||||
| Ci | −0.388 | 0.522 | 0.021 | −0.027 | −0.092 | 0.194 | 1.000 | |||||
| MDA | 0.134 | −0.219 | 0.068 | 0.065 | −0.036 | −0.098 | −0.660 | 1.000 | ||||
| SOD | −0.076 | 0.405 | −0.416 | −0.759 | −0.913 | −0.686 | −0.174 | 0.301 | 1.000 | |||
| Seedling height | 0.241 | −0.344 | −0.122 | 0.113 | 0.162 | 0.157 | −0.711 | 0.447 | −0.077 | 1.000 | ||
| Ground diameter | 0.135 | −0.446 | 0.564 | 0.670 | 0.741 | 0.503 | −0.008 | −0.226 | −0.870 | 0.286 | 1.000 | |
| Root–crown ratio | −0.004 | 0.193 | 0.370 | 0.167 | 0.549 | 0.145 | 0.066 | 0.377 | −0.338 | −0.095 | 0.085 | 1.000 |
| Traits | Components | |||
|---|---|---|---|---|
| P1 | P2 | P3 | P4 | |
| SOD | −0.438 | 0.193 | −0.024 | 0.039 |
| Pn | 0.434 | −0.039 | 0.074 | 0.185 |
| Tr | 0.419 | 0.010 | −0.179 | 0.022 |
| Ground diameter | 0.407 | −0.077 | 0.076 | −0.242 |
| Gs | 0.349 | −0.068 | −0.400 | 0.146 |
| WSD | −0.265 | −0.258 | −0.098 | 0.454 |
| Ci | −0.048 | −0.592 | −0.104 | 0.070 |
| Seedling height | 0.124 | 0.478 | −0.041 | −0.117 |
| MDA | −0.004 | 0.473 | 0.269 | 0.323 |
| Chl A | 0.175 | −0.129 | 0.626 | −0.139 |
| RWC | 0.125 | 0.244 | −0.411 | 0.321 |
| Root–crown ratio | 0.152 | −0.067 | 0.376 | 0.657 |
| Provenance | Yik | |||
|---|---|---|---|---|
| Yi1 | Yi2 | Yi3 | Yi4 | |
| KL | 0.376 | 0.574 | 0.583 | 2.649 |
| PZ | −0.669 | 1.192 | 0.497 | 2.891 |
| SY | 0.236 | 0.335 | 0.377 | 2.697 |
| XW | 0.623 | 0.844 | 0.469 | 2.509 |
| XH | 0.983 | 0.296 | 0.680 | 2.997 |
| JR | −0.618 | 0.742 | 0.789 | 2.983 |
| LG | −0.291 | 0.715 | 0.432 | 2.347 |
| SH | 0.979 | 0.336 | 1.074 | 3.474 |





References
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H.; Lü, S. Molecular Cloning and Functional Analysis of GmLACS2-3 Reveals Its Involvement in Cutin and Suberin Biosynthesis along with Abiotic Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef] [PubMed]
- Rane, J.; Singh, A.K.; Tiwari, M.; Prasad, P.V.V.; Jagadish, S.V.K. Effective Use of Water in Crop Plants in Dryland Agriculture: Implications of Reactive Oxygen Species and Antioxidative System. Front. Plant Sci. 2022, 12, 778270. [Google Scholar] [CrossRef] [PubMed]
- Prăvălie, R. Drylands extent and environmental issues. A global approach. Earth-Sci. Rev. 2016, 161, 259–278. [Google Scholar] [CrossRef]
- Mao, D.; Wang, Z.; Wu, B.; Zeng, Y.; Luo, L.; Zhang, B. Land degradation and restoration in the arid and semiarid zones of China: Quantified evidence and implications from satellites. Land Degrad. Dev. 2018, 29, 3841–3851. [Google Scholar] [CrossRef]
- Sakadevan, K.; Nguyen, M.-L. Extent, Impact, and Response to Soil and Water Salinity in Arid and Semiarid Regions. Adv. Agron. 2010, 109, 55–74. [Google Scholar] [CrossRef]
- Editorial Committee of the Flora of China, Chinese Academy of Sciences. Flora of China; Science Press: Beijing, China, 1997; Volume 143, ISBN 9787030012050. (In Chinese) [Google Scholar]
- Sen, A.; Batra, A. Melia Azedarach L.-A paradise tree. Indian J. 2011, 1, 59–69. [Google Scholar]
- Adan, Y.; Rubea, A.L. The Potential Uses of Melia Azedarach L. as pesticidal and Medicinal Plant, Review. Am.-Eurasian J. Sustain. Agric. 2009, 3, 185–194. [Google Scholar]
- Cheng, S.M.; Gu, W.C. The phenological Division of Distribution area in China for Melia Azedarach. Sci. Silvae Scinicae 2005, 41, 186–191. (In Chinese) [Google Scholar]
- Vishnukanta, A.R. Melia azedarach: A phytopharmacological review. Pharmacogn. Rev. 2008, 2, 173–179. [Google Scholar]
- Sultana, S.; Asif, H.M.; Akhtar, N.; Waqas, M.; Rehman, S.U. Comprehensive review on ethanobotanical use, phytochemistry and pharmacological properties of Melia Azederach Linn. Asian J. Pharm. Res. Health Care 2014, 16, 26–32. [Google Scholar]
- Kumar, R.; Singh, R.; Meera, P.S.; Kalidhar, S.B. Chemical components and insecticidal properties of Bakain (Melia azedarach L.)–A review. Agric. Rev. 2003, 24, 101–115. [Google Scholar]
- Silva, S.; Pinto, G.; Correia, B.; Pinto-Carnide, O.; Santos, C. Rye oxidative stress under long term Al exposure. J. Plant Physiol. 2013, 170, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.C.; Pinto, G.; Santos, C. Acclimatization of micropropagated plantlets induces an antioxidative burst: A case study with Ulmus minor Mill. Photosynthetica 2011, 49, 259–266. [Google Scholar] [CrossRef]
- Jhou, H.-C.; Wang, Y.-N.; Wu, C.-S.; Yu, J.-C.; Chen, C.-I. Photosynthetic gas exchange responses of Swietenia macrophylla King and Melia azedarach L. plantations under drought conditions. Bot. Stud. 2017, 58, 57. [Google Scholar] [CrossRef] [Green Version]
- Dias, M.C.; Azevedo, C.; Costa, M.; Pinto, G.; Santos, C. Melia azedarach plants show tolerance properties to water shortage treatment: An ecophysiological study. Plant Physiol. Biochem. 2014, 75, 123–127. [Google Scholar] [CrossRef]
- DB11/T 770-2010; Growth media for ornamental. Beijing Quality and Technology Supervision Bureau: Beijing, China, 2010.
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000; ISBN 9787040080766. (In Chinese) [Google Scholar]
- Wang, X.K. Principles and Techniques of Plant Physiological and Biochemical Experiments, 3rd ed.; Higher Education Press: Beijing, China, 2006; ISBN 9787040396461. (In Chinese) [Google Scholar]
- Kuromori, T.; Fujita, M.; Takahashi, F.; Shinozaki, K.Y.; Shinozaki, K. Inter-tissue and inter-organ signaling in drought stress response and phenotyping of drought tolerance. Plant J. 2021, 109, 342–358. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Jabbari, H.; Akbari, G.A.; Sima, N.A.K.K.; Rad, A.H.S.; Alahdadi, I.; Hamed, A.; Shariatpanahi, M.E. Relationships between seedling establishment and soil moisture content for winter and spring rapeseed genotypes. Ind. Crop. Prod. 2013, 49, 177–187. [Google Scholar] [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Bogati, K.; Walczak, M. The Impact of Drought Stress on Soil Microbial Community, Enzyme Activities and Plants. Agronomy 2022, 12, 189. [Google Scholar] [CrossRef]
- Zhang, X.; Zang, R.; Li, C. Population differences in physiological and morphological adaptations of Populus davidiana seedlings in response to progressive drought stress. Plant Sci. 2004, 166, 791–797. [Google Scholar] [CrossRef]
- Li, F.; Bao, W.; Wu, N.; You, C. Growth, biomass partitioning, and water-use efficiency of a leguminous shrub (Bauhinia faberi var. microphylla) in response to various water availabilities. New For. 2008, 36, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Lei, Y. Physiological responses of Populus przewalskii to oxidative burst caused by drought stress. Russ. J. Plant Physiol. 2008, 55, 857–864. [Google Scholar] [CrossRef]
- Brunner, I.; Herzog, C.; Dawes, M.A.; Arend, M.; Sperisen, C. How tree roots respond to drought. Front. Plant Sci. 2015, 6, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eying, Y.; Song, L.L.; Jacobs, D.; Emei, L.; Eliu, P.; Ejin, S.; Ewu, J. Physiological response to drought stress in Camptotheca acuminata seedlings from two provenances. Front. Plant Sci. 2015, 6, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought re-sistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef]
- Lei, Y.; Yin, C.; Li, C. Differences in some morphological, physiological, and biochemical responses to drought stress in two contrasting populations of Populus przewalskii. Physiol. Plant. 2006, 127, 182–191. [Google Scholar] [CrossRef]
- Goche, T.; Shargie, N.G.; Cummins, I.; Brown, A.P.; Chivasa, S.; Ngara, R. Comparative physiological and root proteome analyses of two sorghum varieties responding to water limitation. Sci. Rep. 2020, 10, 11835. [Google Scholar] [CrossRef]
- Corso, D.; Delzon, S.; Lamarque, L.J.; Cochard, H.; Torres-Ruiz, J.M.; King, A.; Brodribb, T. Neither xylem collapse, cavitation, or changing leaf conductance drive stomatal closure in wheat. Plant Cell Environ. 2020, 43, 854–865. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Siddique, M.R.B.; Hamid, A.; Islam, M.S. Drought stress effects on water relations of wheat. Bot. Bull. Acad. Sin. 2000, 41, 35–39. [Google Scholar]
- Nowicka, B.; Ciura, J.; Szymanska, R.; Kruk, J. Improving photosynthesis, plant productivity and abiotic stress tolerance current trends and future perspectiv-es. J. Plant Physiol. 2018, 231, 415–433. [Google Scholar] [CrossRef] [PubMed]
- Bohnert, H.J.; Jensen, R.G. Strategies for engineering water-stress tolerance in plants. Trends Biotechnol. 1996, 14, 89–97. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Rawat, J.M.; Rawat, B.; Tewari, A.; Joshi, S.C.; Nandi, S.K.; Palni, L.M.S.; Prakash, A. Alterations in growth, photosynthetic activity and tissue-water relations of tea clones in response to different soil moisture content. Trees 2017, 31, 941–952. [Google Scholar] [CrossRef]
- Silva, E.N.; Ribeiro, R.V.; Ferreira-Silva, S.L.; Vieira, S.A.; Ponte, L.F.; Silveira, J.A. Coordinate changes in photosynthesis, sugar accumulation and antioxidative enzymes improve the performance of Jatropha curcas plants under drought stress. Biomass-Bioenergy 2012, 45, 270–279. [Google Scholar] [CrossRef]
- Wu, F.Z.; Bao, W.K.; Li, F.L.; Wu, N. Effects of water stress and nitrogen supply on leaf gas exchange and fluorescence parameters of Sophora davidii seedlings. Photosynthetica 2008, 46, 40–48. [Google Scholar] [CrossRef]
- Gao, L.; Caldwell, C.D.; Jiang, Y. Photosynthesis and Growth of Camelina and Canola in Response to Water Deficit and Applied Nitrogen. Crop Sci. 2018, 58, 393–401. [Google Scholar] [CrossRef]
- Smirnoff, N. Antioxidant systems and plant response to the environment. In Environment and Plant Metab-Olism: Flexibility and Acclimation; Smirnoff, N., Ed.; BIOS Scientific Publishers: Oxford, UK, 1995; pp. 217–243. [Google Scholar]
- Oster, U.; Tanaka, R.; Tanaka, A.; Rüdiger, W. Cloning and functional expression of the gene encoding the key enzyme for chlorophyll b biosynthesis (CAO) from Arabidopsis thaliana. Plant J. 2000, 21, 305–310. [Google Scholar] [CrossRef]
- Ghobadi, M.; Taherabadi, S.; Ghobadi, M.-E.; Mohammadi, G.-R.; Jalali-Honarmand, S. Antioxidant capacity, photosynthetic characteristics and water relations of sunflower (Helianthus annuus L.) cultivars in response to drought stress. Ind. Crop. Prod. 2013, 50, 29–38. [Google Scholar] [CrossRef]
- Efeoğlu, B.; Ekmekçi, Y.; Çiçek, N. Physiological responses of three maize cultivars to drought stress and recovery. S. Afr. J. Bot. 2009, 75, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Zheng, F.; Zhang, D.; Li, C.; Li, M.; Ye, J.; Zhang, Y.; Wang, T.; Ouyang, B.; Hong, Z.; et al. Tomato methionine sulfoxide reductase B2 functions in drought tolerance by promoting ROS scavenging and chlorophyll accumulation through interaction with Catalase 2 and RBCS3B. Plant Sci. 2022, 318, 111206. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.R.; Guo, X.P.; Wang, W.M.; Zhang, L.J.; Wang, Q. Effect of water stress and rehydration on leaf chloroplast pigments during rice nodulation. J. Hohai Univ.-Y (Nat. Sci. Ed.) 2006, 4, 397–400. [Google Scholar]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, M.; Munné-Bosch, S. Malondialdehyde: Facts and Artifacts. Plant Physiol. 2019, 180, 1246–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 19250. [Google Scholar] [CrossRef]
- Carvalho, M.; Castro, I.; Moutinho-Pereira, J.; Correia, C.; Egea-Cortines, M.; Matos, M.; Rosa, E.; Carnide, V.; Lino-Neto, T. Evaluating stress responses in cowpea under drought stress. J. Plant Physiol. 2019, 241, 153001. [Google Scholar] [CrossRef]
- Wang, L.-F. Physiological and molecular responses to drought stress in rubber tree (Hevea brasiliensis Muell. Arg.). Plant Physiol. Biochem. 2014, 83, 243–249. [Google Scholar] [CrossRef]
- Zhang, M.; Jin, Z.-Q.; Zhao, J.; Zhang, G.; Wu, F. Physiological and biochemical responses to drought stress in cultivated and Tibetan wild barley. Plant Growth Regul. 2015, 75, 567–574. [Google Scholar] [CrossRef]















| Original Area | Longitude | Latitude | Altitude/m | Annual Average Temperature/°C | Annual Precipitation/mm | Frost-Free Season/d |
|---|---|---|---|---|---|---|
| KL | 118.58 | 37.42 | 3~5 | 12.8 | 555.9 | 206 |
| PZ | 117.99 | 34.35 | 18~32 | 14.2 | 868.6 | 210 |
| SY | 112.18 | 30.37 | 30~55 | 15.9 | 1250 | 255 |
| XW | 118.83 | 32.09 | 12~20 | 15.4 | 1106.5 | 237 |
| XH | 115.87 | 28.67 | 55~112 | 17.6 | 1458 | 277 |
| JR | 119.13 | 32.07 | 5~30 | 15.2 | 1055.6 | 233 |
| LG | 113.25 | 23.11 | 15~22 | 21 | 1720 | 190 |
| SH | 114.01 | 31.82 | 60~150 | 15.3 | 1100 | 225 |
| Provenance | Seedling Height Growth/cm | Relative Decline I/% | Relative Decline II/% | Relative Decline III/% | |||
|---|---|---|---|---|---|---|---|
| CK | I | II | III | ||||
| KL | 22.80 ± 5.25 a | 11.75 ± 4.25 ab | 8.98 ± 2.53 b | 3.65 ± 1.93 b | 48.47% | 60.64% | 83.99% |
| PZ | 19.00 ± 0.53 a | 14.88 ± 1.13 a | 2.93 ± 2.33 b | 3.48 ± 1.14 b | 21.71% | 84.61% | 81.71% |
| SY | 24.63 ± 5.44 a | 13.93 ± 3.94 ab | 10.40 ± 2.52 b | 4.18 ± 0.65 b | 43.45% | 57.77% | 83.05% |
| XW | 19.53 ± 0.95 a | 14.50 ± 2.10 ab | 13.25 ± 3.57 ab | 9.40 ± 2.39 b | 25.77% | 32.17% | 51.88% |
| XH | 24.55 ± 0.05 a | 17.35 ± 3.45 a | 14.70 ± 0.80 a | 9.00 ± 8.10 a | 29.33% | 40.12% | 63.34% |
| JR | 19.67 ± 6.63 a | 13.08 ± 4.51 ab | 3.30 ± 1.19 b | 1.63 ± 0.31 b | 33.52% | 83.22% | 91.74% |
| LG | 15.93 ± 1.26 a | 14.65 ± 4.01 a | 6.65 ± 2.02 b | 4.55 ± 2.20 b | 8.01% | 71.43% | 58.24% |
| SH | 13.43 ± 3.42 a | 11.68 ± 1.66 a | 7.23 ± 3.40 ab | 1.78 ± 0.63 b | 13.00% | 46.00% | 87.00% |
| Provenance | Diameter Growth/cm | Relative Decline I/% | Relative Decline II/% | Relative Decline III/% | |||
|---|---|---|---|---|---|---|---|
| CK | I | II | III | ||||
| KL | 1.02 ± 0.30 a | 0.80 ± 0.10 a | 0.97 ± 0.40 a | 0.31 ± 0.10 a | 22% | 5% | 70% |
| PZ | 1.10 ± 0.08 a | 0.40 ± 0.23 b | 0.34 ± 0.05 b | 0.19 ± 0.07 b | 64% | 69% | 83% |
| SY | 0.89 ± 0.13 a | 0.76 ± 0.25 a | 0.53 ± 0.30 a | 0.28 ± 0.06 a | 15% | 40% | 69% |
| XW | 1.39 ± 0.32 a | 0.89 ± 0.08 b | 0.44 ± 0.05 c | 0.30 ± 0.10 c | 36% | 69% | 79% |
| XH | 1.05 ± 0.07 a | 0.64 ± 0.21 a | 0.50 ± 0.16 a | 0.52 ± 0.10 a | 40% | 53% | 51% |
| JR | 1.10 ± 0.21 a | 0.52 ± 0.05 b | 0.47 ± 0.13 b | 0.23 ± 0.04 b | 53% | 57% | 79% |
| LG | 2.74 ± 1.18 a | 0.89 ± 0.26 ab | 0.59 ± 0.14 b | 0.34 ± 0.09 b | 68% | 78% | 88% |
| SH | 1.28 ± 0.11 a | 0.62 ± 0.13 b | 0.40 ± 0.08 bc | 0.20 ± 0.07 c | 52% | 69% | 85% |
| Provenance | Biomass/g·Plant-1 | |||||||
|---|---|---|---|---|---|---|---|---|
| The Aboveground Part/g·Plant-1 | The Underground Part/g·Plant-1 | |||||||
| CK | I | II | II | CK | I | II | III | |
| KL | 16.76 ± 3.04 a | 16.59 ± 0.99 a | 16.49 ± 3.14 a | 11.98 ± 1.34 a | 13.81 ± 3.79 a | 14.51 ± 2.30 a | 14.33 ± 3.02 a | 16.27 ± 1.79 a |
| PZ | 16.40 ± 1.47 a | 13.94 ± 2.78 a | 13.91 ± 2.73 a | 12.14 ± 3.10 a | 17.10 ± 3.03 a | 17.56 ± 4.44 a | 23.51 ± 2.68 a | 21.98 ± 1.97 a |
| SY | 17.03 ± 2.54 a | 16.62 ± 3.04 a | 15.32 ± 0.87 a | 14.71 ± 0.39 a | 10.47 ± 0.81 a | 12.35 ± 0.32 a | 13.67 ± 3.26 a | 15.09 ± 1.64 a |
| XW | 19.31 ± 5.24 a | 15.76 ± 0.74 a | 14.24 ± 2.43 a | 13.49 ± 1.21 a | 15.75 ± 0.61 a | 16.61 ± 2.94 a | 16.63 ± 2.39 a | 17.43 ± 1.70 a |
| XH | 27.49 ± 11.15 a | 20.02 ± 2.71 a | 16.37 ± 4.98 a | 14.25 ± 0.18 a | 12.74 ± 2.83 a | 13.44 ± 2.57 a | 16.11 ± 1.65 a | 17.79 ± 6.65 a |
| JR | 28.72 ± 0.98 a | 25.62 ± 1.23 a | 23.00 ± 2.47 ab | 18.85 ± 0.17 b | 17.33 ± 1.29 a | 19.70 ± 2.05 a | 20.04 ± 2.12 a | 23.16 ± 0.56 a |
| LG | 26.85 ± 6.07 a | 26.40 ± 5.61 a | 22.48 ± 0.05 a | 21.13 ± 1.65 a | 18.25 ± 3.56 a | 13.91 ± 2.37 a | 17.17 ± 3.78 a | 21.00 ± 2.01 a |
| SH | 21.26 ± 3.20 a | 16.04 ± 1.01 a | 15.58 ± 2.30 a | 15.53 ± 3.16 a | 11.41 ± 0.88 b | 14.46 ± 1.44 b | 15.18 ± 1.89 b | 23.55 ± 1.86 a |
| Provenance | Root-Crown Ratio | |||
|---|---|---|---|---|
| CK | I | II | III | |
| KL | 0.81 ± 0.08 a | 0.89 ± 0.19 a | 0.87 ± 0.02 a | 1.39 ± 0.31 a |
| PZ | 1.03 ± 0.09 a | 1.38 ± 0.59 a | 1.80 ± 0.55 a | 1.89 ± 0.32 a |
| SY | 0.62 ± 0.05 a | 0.77 ± 0.12 a | 0.92 ± 0.20 a | 0.98 ± 0.05 a |
| XW | 0.89 ± 0.27 a | 1.16 ± 0.15 a | 1.06 ± 0.01 a | 1.31 ± 0.24 a |
| XH | 0.60 ± 0.35 a | 0.67 ± 0.04 a | 1.05 ± 0.22 a | 1.25 ± 0.48 a |
| JR | 0.61 ± 0.07 b | 0.77 ± 0.12 b | 0.87 ± 0.00 b | 1.23 ± 0.02 a |
| LG | 0.76 ± 0.31 a | 0.52 ± 0.02 a | 0.81 ± 0.18 a | 0.93 ± 0.02 a |
| SH | 0.54 ± 0.18 b | 0.90 ± 0.28 b | 0.97 ± 0.19 ab | 1.52 ± 0.25 a |
| Principal Component | Eigenvalues | Contribution Ratio/% | Cumulative Contribution Rate/% |
|---|---|---|---|
| 1 | 4.404 | 36.699 | 36.699 |
| 2 | 2.729 | 22.744 | 59.442 |
| 3 | 1.665 | 13.877 | 73.320 |
| 4 | 1.586 | 13.219 | 86.539 |
| Principal Component | Eigenvalues | Contribution Ratio/% | Cumulative Contribution Rate/% |
|---|---|---|---|
| 1 | 4.285 | 35.707 | 35.707 |
| 2 | 3.005 | 25.040 | 60.748 |
| 3 | 2.164 | 18.034 | 78.782 |
| 4 | 1.500 | 12.503 | 91.285 |
| Principal Component | Eigenvalues | Contribution Ratio/% | Cumulative Contribution Rate/% |
|---|---|---|---|
| 1 | 4.285 | 35.707 | 35.707 |
| 2 | 3.005 | 25.040 | 60.748 |
| 3 | 2.164 | 18.034 | 78.782 |
| 4 | 1.500 | 12.503 | 91.285 |
| Provenance | U(Yik) | D Value | Rank | |||
|---|---|---|---|---|---|---|
| U(Yi1) | U(Yi2) | U(Yi3) | U(Yi4) | |||
| KL | 0.429 | 0.561 | 0.549 | 0.511 | 0.495 | 5 |
| PZ | 0.469 | 0.357 | 0.540 | 0.044 | 0.386 | 7 |
| SY | 0.544 | 0 | 0.496 | 0.311 | 0.358 | 8 |
| XW | 0.776 | 0.276 | 0.053 | 0.933 | 0.553 | 4 |
| XH | 0.714 | 0.551 | 0.788 | 0.4448 | 0.642 | 1 |
| JR | 0.633 | 0.429 | 0.6288 | 0.5338 | 0.563 | 3 |
| LG | 0 | 1 | 1 | 1 | 0.576 | 2 |
| SH | 1 | 0.010 | 0 | 0 | 0.427 | 6 |
| Weights | 0.424 | 0.263 | 0.160 | 0.153 | ||
| Provenance | U(Yik) | D Value | Rank | |||
|---|---|---|---|---|---|---|
| U(Yi1) | U(Yi2) | U(Yi3) | U(Yi4) | |||
| KL | 0.731 | 0.280 | 0.192 | 0.617 | 0.485 | 3 |
| PZ | 0.059 | 0.354 | 1 | 0.808 | 0.429 | 6 |
| SY | 0.333 | 0.648 | 0 | 0.524 | 0.380 | 7 |
| XW | 0.628 | 0.383 | 0.129 | 0.538 | 0.450 | 5 |
| XH | 0.456 | 0.650 | 0.688 | 0.757 | 0.596 | 2 |
| JR | 0.311 | 0.431 | 0.375 | 1 | 0.451 | 4 |
| LG | 0 | 0 | 0.190 | 0.823 | 0.150 | 8 |
| SH | 1 | 1 | 0.274 | 0 | 0.719 | 1 |
| Weights | 0.391 | 0.274 | 0.198 | 0.137 | ||
| Provenance | U(Yik) | D Value | Rank | |||
|---|---|---|---|---|---|---|
| U(Yi1) | U(Yi2) | U(Yi3) | U(Yi4) | |||
| KL | 0.633 | 0.310 | 0.296 | 0.268 | 0.444 | 4 |
| PZ | 0 | 1 | 0.172 | 0.483 | 0.346 | 5 |
| SY | 0.548 | 0.044 | 0 | 0.311 | 0.296 | 7 |
| XW | 0.782 | 0.612 | 0.132 | 0.144 | 0.541 | 3 |
| XH | 1 | 0 | 0.435 | 0.577 | 0.595 | 2 |
| JR | 0.031 | 0.498 | 0.591 | 0.564 | 0.316 | 6 |
| LG | 0.229 | 0.468 | 0.079 | 0 | 0.232 | 8 |
| SH | 0.998 | 0.045 | 1 | 1 | 0.759 | 1 |
| Weights | 0.442 | 0.251 | 0.170 | 0.137 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, C.; Chen, J.; Liu, Z.; Chen, H.; Yu, F.; Yu, W. Morphological and Physiological Responses of Melia azedarach Seedlings of Different Provenances to Drought Stress. Agronomy 2022, 12, 1461. https://doi.org/10.3390/agronomy12061461
Han C, Chen J, Liu Z, Chen H, Yu F, Yu W. Morphological and Physiological Responses of Melia azedarach Seedlings of Different Provenances to Drought Stress. Agronomy. 2022; 12(6):1461. https://doi.org/10.3390/agronomy12061461
Chicago/Turabian StyleHan, Chao, Junna Chen, Zemao Liu, Hong Chen, Fangyuan Yu, and Wanwen Yu. 2022. "Morphological and Physiological Responses of Melia azedarach Seedlings of Different Provenances to Drought Stress" Agronomy 12, no. 6: 1461. https://doi.org/10.3390/agronomy12061461