
Irrigation and Nitrogen Application Promote Population Density through Altered Bud Bank Size and Components in Leymus chinensis

Abstract
:1. Introduction
2. Materials and Methods
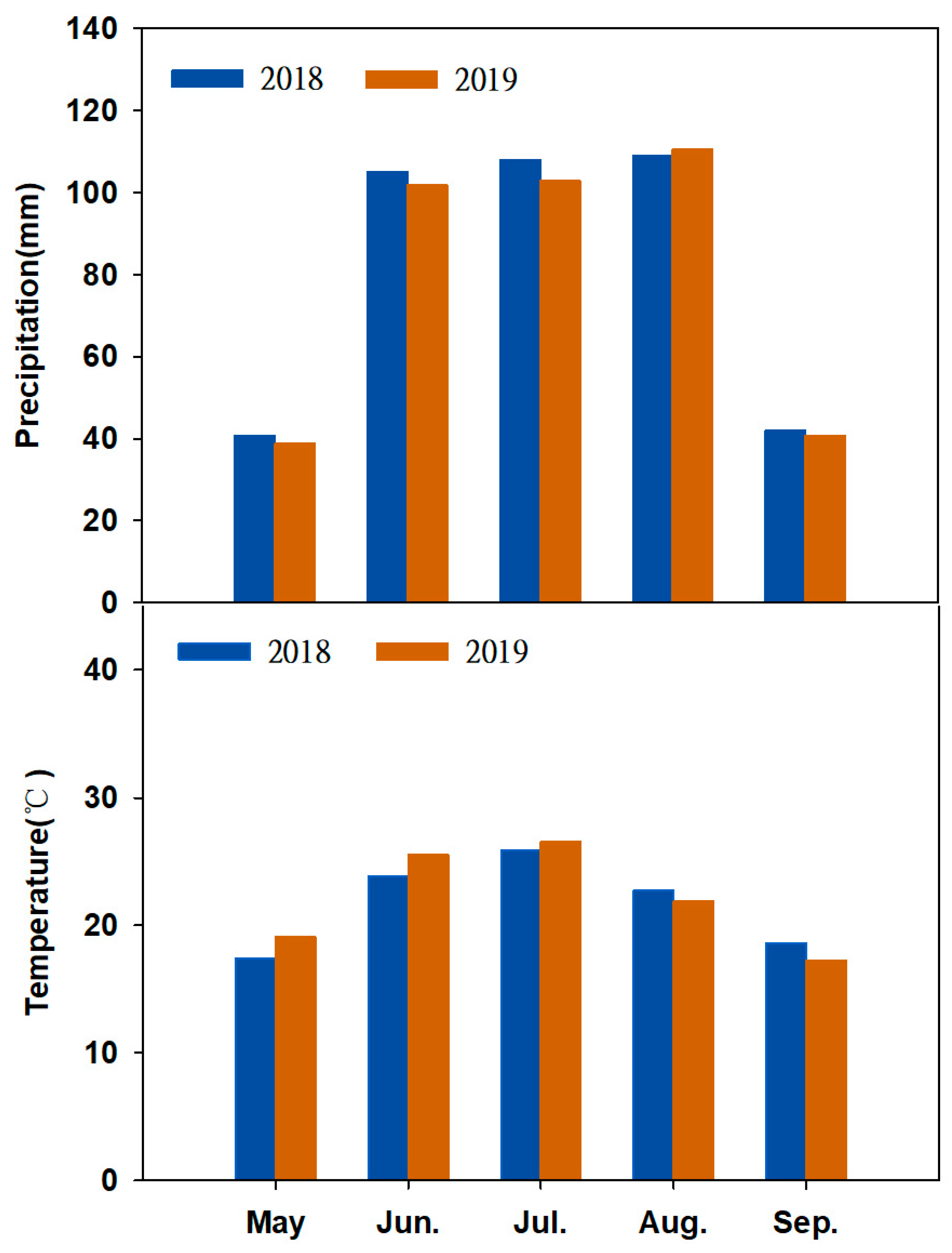
2.1. Study Sites
2.2. Experimental Design
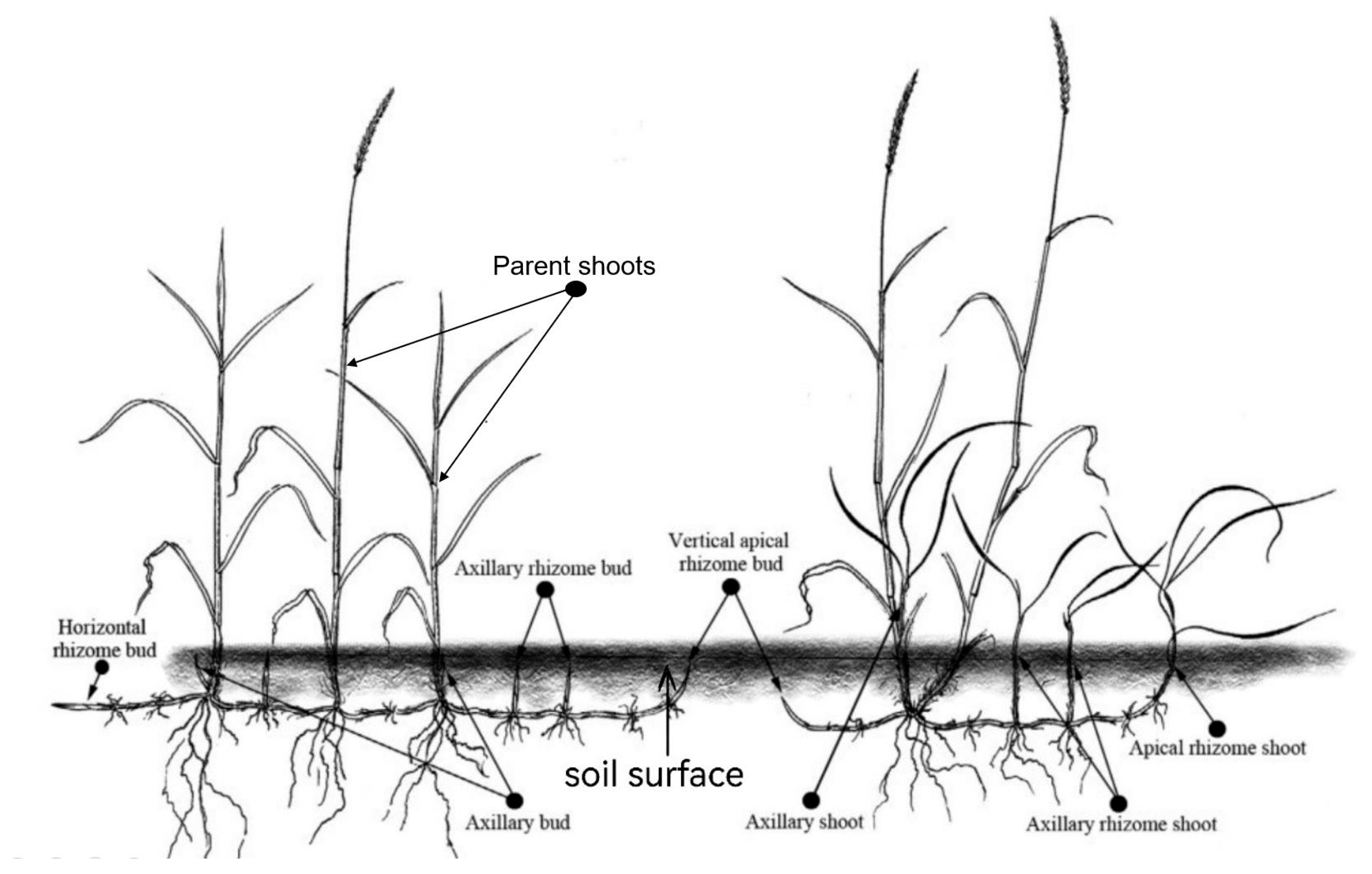
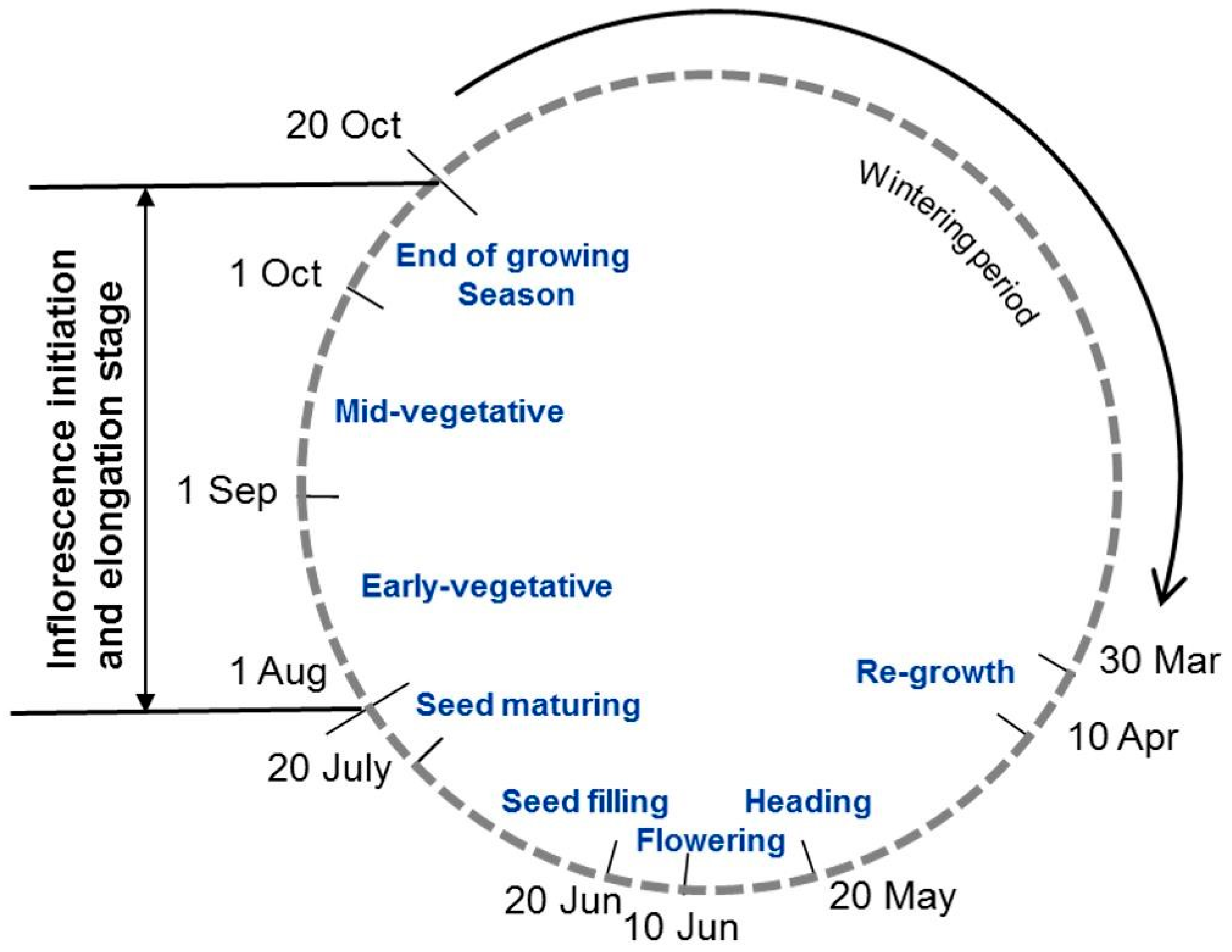
2.3. Data Collection
2.4. Data Analysis
3. Results
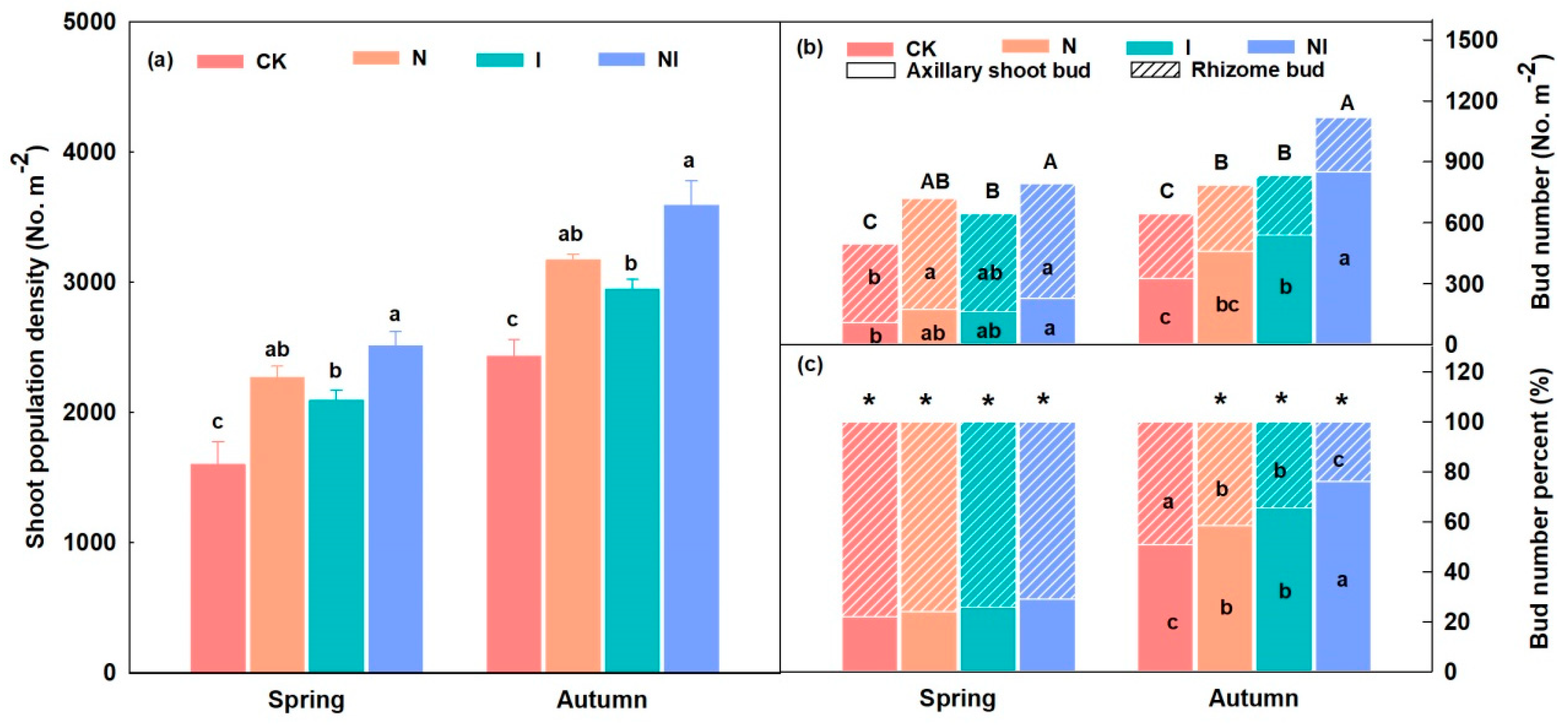
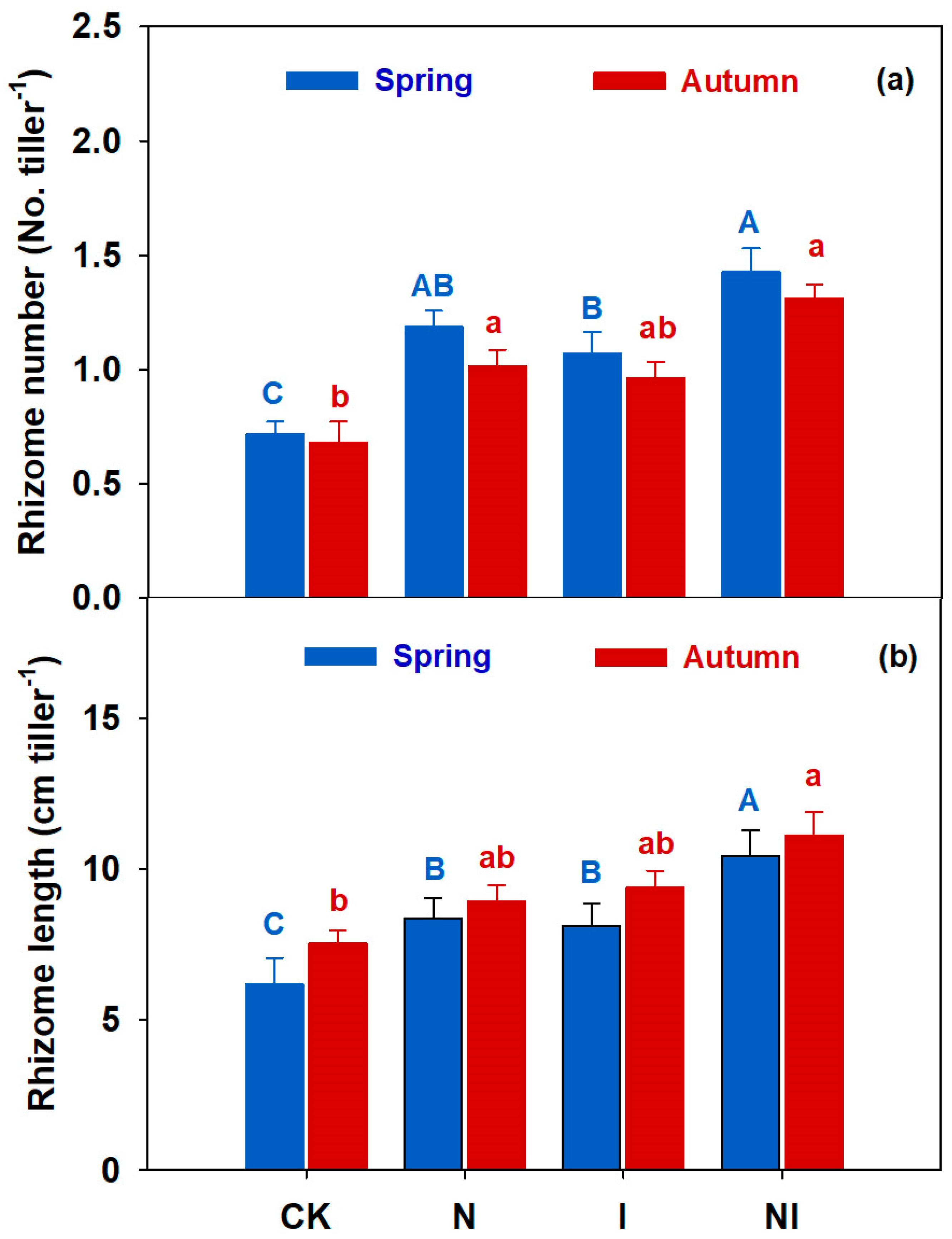
3.1. Shoot Population Density and Bud Bank and Rhizome Traits
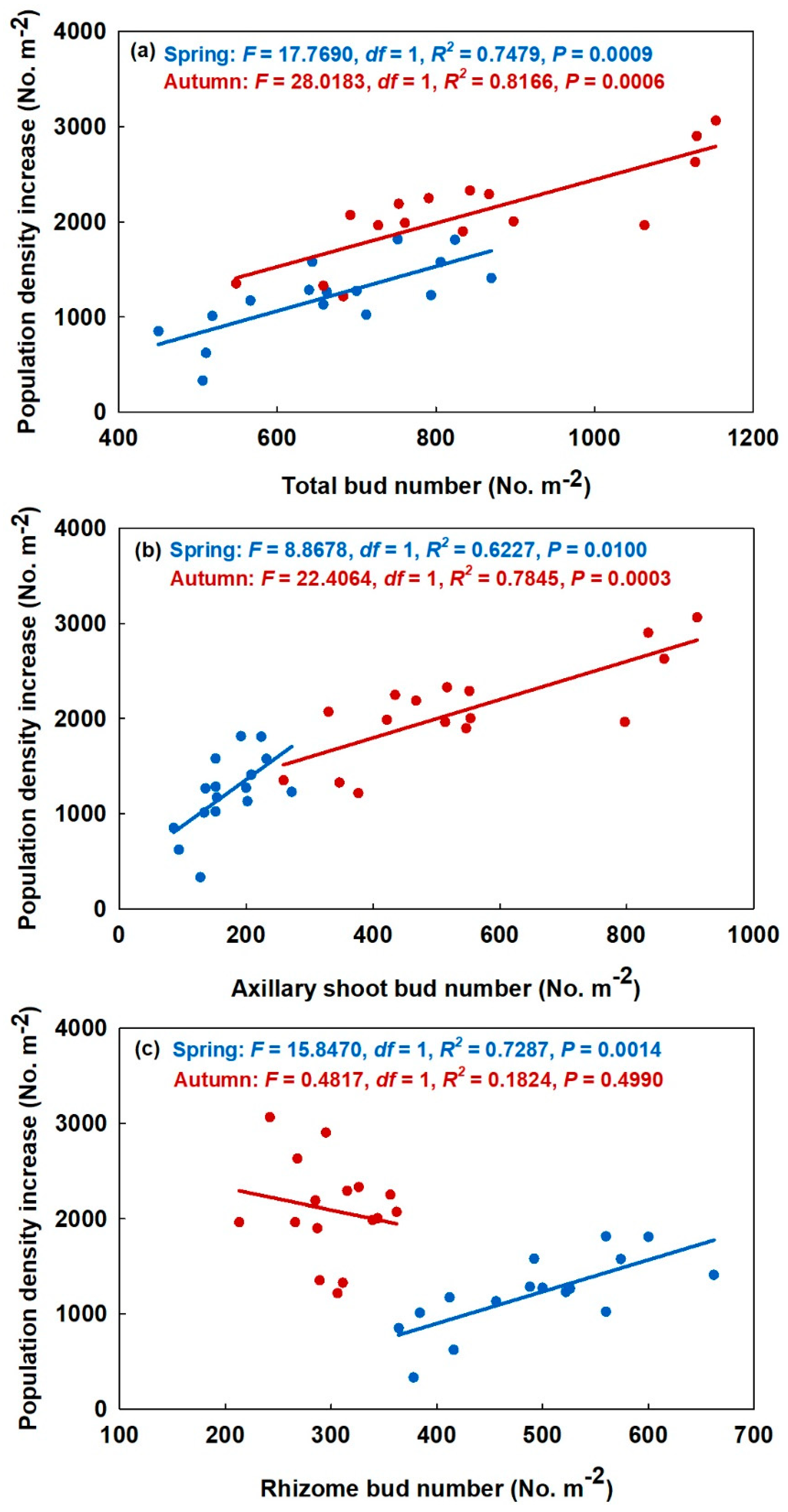
3.2. Relationship between Bud Bank and the Aboveground Population
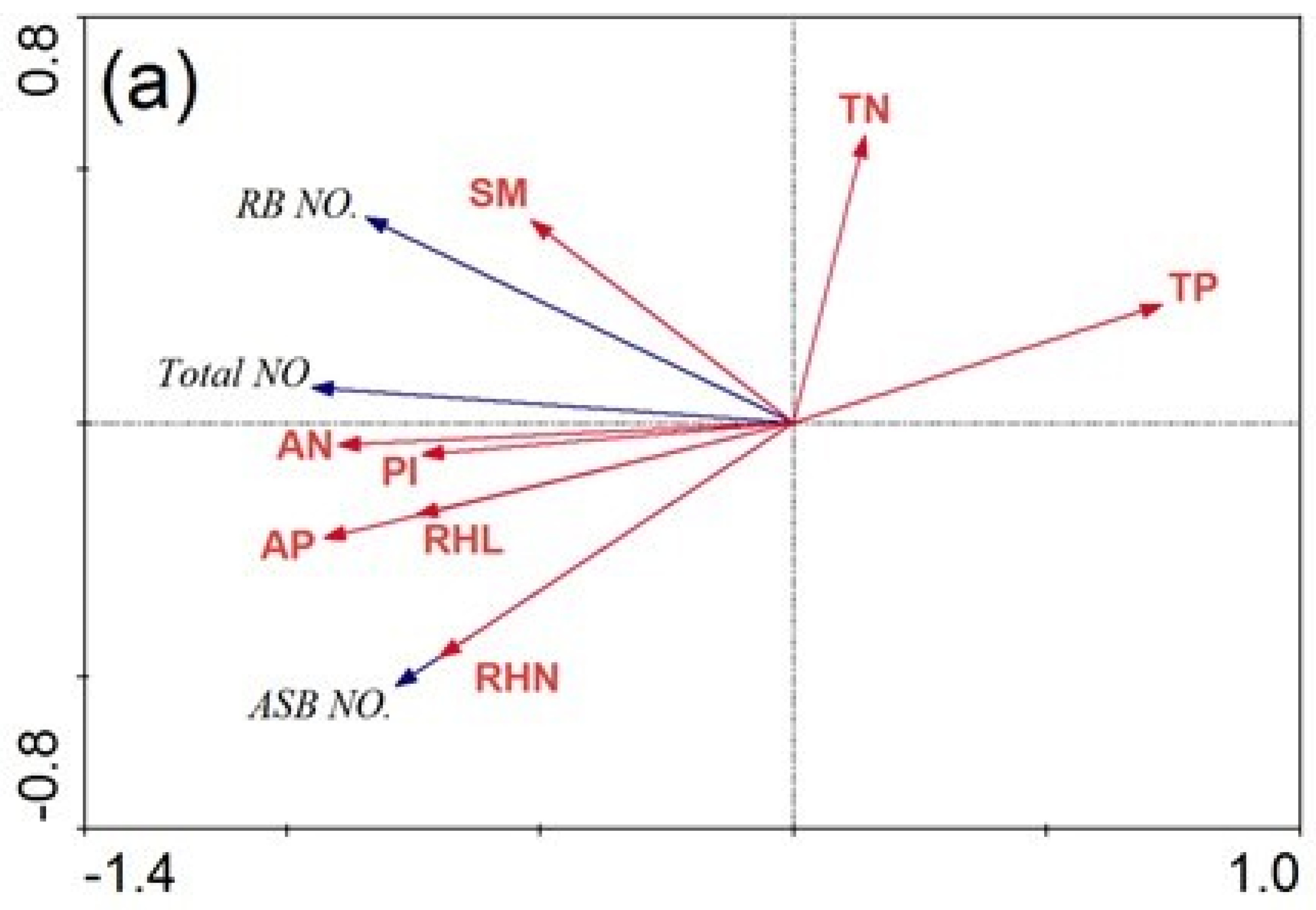
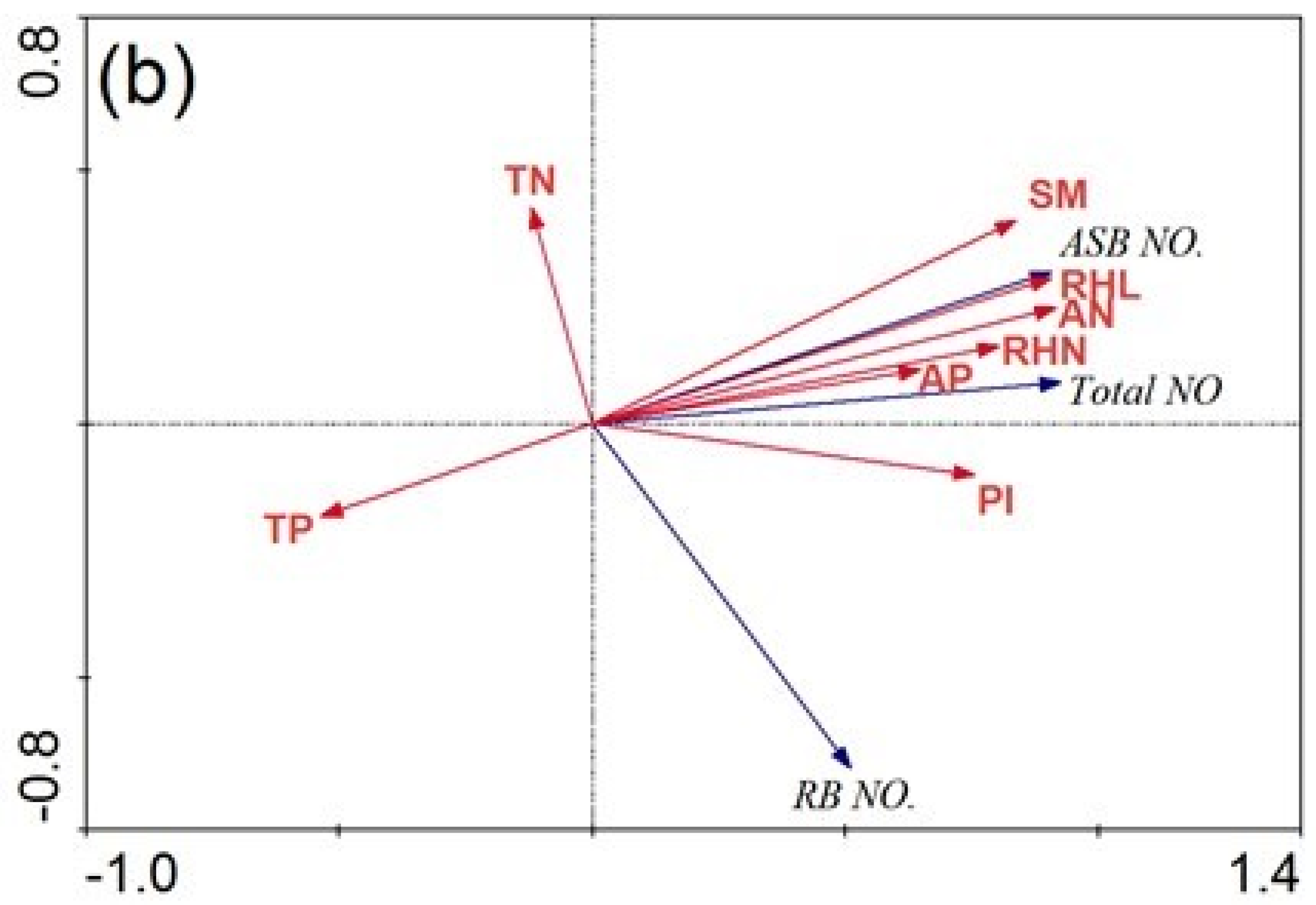
3.3. The Impact of Soil Nutrient Properties and Rhizome Traits on the Bud Bank
4. Discussion
4.1. Bud Bank Composition and Seasonal Variation
4.2. Relationship between the Bud Bank and Its Adaptive Strategy and Shoot Population Density Increase
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Controlling Factors | Parameters | Treatment | |||
|---|---|---|---|---|---|---|
| CK | N | I | N*I | |||
| Spring | SE | TN (mg·kg−1) | 235.76 ± 10.80 | 249.90 ± 61.19 | 277.49 ± 70.92 | 234.14 ± 22.97 |
| TP (mg·kg−1) | 300.70 ± 68.36 | 192.41 ± 39.60 | 255.77 ± 19.37 | 121.68 ± 10.33 | ||
| AN (mg·kg−1) | 2.95 ± 0.10 C | 5.33 ± 0.55 AB | 4.80 ± 0.25 B | 6.74 ± 0.65 A | ||
| AP (mg·kg−1) | 11.35 ± 2.13 C | 16.21 ± 0.44 AB | 15.12 ± 0.69 AB | 17.98 ± 0.64 A | ||
| SM (%) | 13.37 ± 0.25 B | 13.94 ± 0.19 AB | 14.75 ± 0.20 A | 14.15 ± 0.40 AB | ||
| PT | PI (No·m−2) | 704.75 ± 147.68 C | 1382.50 ± 73.66 AB | 1153.00 ± 53.70 B | 1608.25 ± 137.81 A | |
| Autumn | SE | TN (mg·kg−1) | 233.19 ± 10.09 | 311.91 ± 45.03 | 245.13 ± 51.68 | 259.35 ± 29.34 |
| TP (mg·kg−1) | 306.76 ± 17.46 A | 189.29 ± 11.95 B | 196.80 ± 41.86 B | 199.27 ± 9.79 B | ||
| AN (mg·kg−1) | 3.15 ± 0.16 C | 5.15 ± 0.44 B | 5.50 ± 0.24 B | 7.44 ± 0.34 A | ||
| AP (mg·kg−1) | 12.55 ± 1.60 C | 17.31 ± 0.32 A | 15.38 ± 0.68 AB | 19.11 ± 0.45 A | ||
| SM (%) | 13.38 ± 0.25 C | 14.01 ± 0.13 BC | 14.95 ± 0.29 AB | 15.08 ± 0.16 A | ||
| PT | PI (No·m−2) | 1492.00 ± 195.59 C | 2188.00 ± 72.57 AB | 2039.50 ± 86.20 B | 2639.50 ± 242.48 A | |

References
- Xia, J.; Wan, S. Global response patterns of terrestrial plant species to nitrogen addition. New Phytol. 2008, 179, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Rao, L.E.; Allen, E.B. Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts. Oecologia 2009, 162, 1035–1046. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-F.; Gao, S.; Lin, J.-X.; Mu, Y.-G.; Mu, C.-S. Summer warming effects on biomass production and clonal growth of Leymus chinensis. Crop Pasture Sci. 2010, 61, 670–676. [Google Scholar] [CrossRef]
- Ott, J.P.; Butler, J.L.; Rong, Y.P.; Xu, L. Greater bud outgrowth of Bromus inermis than Pascopyrum smithii under multiple environmental conditions. J. Plant Ecol. 2017, 10, 518–527. [Google Scholar]
- Wang, J.F.; Shi, Y.J.; Ao, Y.N.; Yu, D.F.; Wang, J.; Gao, S. Summer drought decreases Leymus chinensis productivity through constraining the bud, tiller and shoot production. J. Agron. Crop Sci. 2019, 205, 554–561. [Google Scholar] [CrossRef]
- Hartnett, D.C.; Setshogo, M.P.; Dalgleish, H.J. Bud banks of perennial savanna grasses in Botswana. Afr. J. Ecol. 2006, 44, 256–263. [Google Scholar] [CrossRef]
- Ott, J.P.; Klimešová, J.; Hartnett, D.C. The ecology and significance of below-ground bud banks in plants. Ann. Bot. 2019, 123, 1099–1118. [Google Scholar] [CrossRef]
- Chen, X.-S.; Deng, Z.-M.; Xie, Y.-H.; Li, F.; Hou, Z.-Y.; Li, X. Belowground bud banks of four dominant macrophytes along a small-scale elevational gradient in Dongting Lake wetlands, China. Aquat. Bot. 2015, 122, 9–14. [Google Scholar] [CrossRef]
- Ma, Q.; Qian, J.; Tian, L.; Liu, Z. Responses of belowground bud bank to disturbance and stress in the sand dune ecosystem. Ecol. Indic. 2019, 106, 105521. [Google Scholar] [CrossRef]
- Worley, A.C.; Harder, L.D. Consequences of preformation for dynamic resource allocation by a carnivorous herb, Pinguicula vulgaris (Lentibulariaceae). Am. J. Bot. 1999, 86, 1136–1145. [Google Scholar] [CrossRef] [Green Version]
- VanderWeide, B.L.; Hartnett, D.C. Belowground bud bank response to grazing under severe, short-term drought. Oecologia 2015, 178, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, L.G.; Sala, O. Differential sensitivities of grassland structural components to changes in precipitation mediate productivity response in a desert ecosystem. Funct. Ecol. 2014, 28, 1292–1298. [Google Scholar] [CrossRef] [Green Version]
- Ott, J.P.; Hartnett, D.C. Contrasting bud bank dynamics of two co-occurring grasses in tallgrass prairie: Implications for grassland dynamics. Plant Ecol. 2012, 213, 1437–1448. [Google Scholar] [CrossRef]
- Ott, J.P.; Hartnett, D.C. Higher-order bud production increases tillering capacity in the perennial caespitose grass Scribner’s Panicum (Dichanthelium oligosanthes). Botany 2012, 90, 884–890. [Google Scholar] [CrossRef]
- Shi, Y.; Gao, S.; Zhou, D.; Liu, M.; Wang, J.; Knops, J.M.; Mu, C. Fall nitrogen application increases seed yield, forage yield and nitrogen use efficiency more than spring nitrogen application in Leymus chinensis, a perennial grass. Field Crop. Res. 2017, 214, 66–72. [Google Scholar] [CrossRef]
- Ma, H.; Zheng, C.; Gao, Y.; Baskin, C.C.; Sun, H.; Yang, H. Moderate clipping stimulates over-compensatory growth of Leymus chinensis under saline-alkali stress through high allocation of biomass and nitrogen to shoots. Plant Growth Regul. 2020, 92, 95–106. [Google Scholar] [CrossRef]
- Zhang, J.-T.; Mu, C.-S.; Wang, D.-L.; Wang, J.-F.; Chen, G.-X. Shoot population recruitment from a bud bank over two seasons of undisturbed growth of Leymus chinensis. Botany 2009, 87, 1242–1249. [Google Scholar] [CrossRef]
- Bernard, J.M. Life history and vegetative reproduction in Carex. Can. J. Bot. 1990, 68, 13–21. [Google Scholar] [CrossRef]
- Chen, X.-S.; Xie, Y.-H.; Deng, Z.-M.; Li, F.; Hou, Z.-Y. A change from phalanx to guerrilla growth form is an effective strategy to acclimate to sedimentation in a wetland sedge species Carex brevicuspis (Cyperaceae). Flora 2011, 206, 347–350. [Google Scholar] [CrossRef]
- Deng, Z.; Chen, X.; Xie, Y.; Li, X.; Pan, Y.; Li, F. Effects of size and vertical distribution of buds on sprouting and plant growth of the clonal emergent macrophyte Miscanthus sacchariflorus (Poaceae). Aquat. Bot. 2013, 104, 121–126. [Google Scholar] [CrossRef]
- Wang, Z.W.; Xu, A.K.; Zhu, T.C. Plasticity in bud demography of a rhizomatous clonal plant Leymus chinensis L. in response to soil water status. J. Plant Biol. 2008, 51, 102–107. [Google Scholar] [CrossRef]
- Dalgleish, H.J.; Kula, A.R.; Hartnett, D.C.; Sandercock, B.K. Responses of two bunchgrasses to nitrogen addition in tallgrass prairie: The role of bud bank demography. Am. J. Bot. 2008, 95, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.; Sun, X.; Wang, Z.; Li, L. Nitrogen addition and rhizome severing modify clonal growth and reproductive modes of Leymus chinensis population. Plant Ecol. 2009, 205, 13–21. [Google Scholar] [CrossRef]
- Tomlinson, K.W.; O’Connor, T.G. Control of tiller recruitment in bunchgrasses: Uniting physiology and ecology. Funct. Ecol. 2004, 18, 489–496. [Google Scholar] [CrossRef]
- Zhang, L.-H.; Zhang, Y.-W.; Zhao, X.-N.; Huang, S.-J.; Zhao, J.-M.; Yang, Y.-F. Effects of different nutrient sources on plasticity of reproductive strategies in a monoecious species, Sagittaria graminea (Alismataceae). J. Syst. Evol. 2013, 52, 84–91. [Google Scholar] [CrossRef]
- Ye, X.-H.; Yu, F.-H.; Dong, M. A Trade-off Between Guerrilla and Phalanx Growth Forms in Leymus secalinus Under Different Nutrient Supplies. Ann. Bot. 2006, 98, 187–191. [Google Scholar] [CrossRef] [Green Version]
- Williamson, M.M.; Wilson, G.W.; Hartnett, D.C. Controls on bud activation and tiller initiation in C3 and C4 tallgrass prairie grasses: The role of light and nitrogen. Botany 2012, 90, 1221–1228. [Google Scholar] [CrossRef]
- Russell, M.L.; Vermeire, L.; Ganguli, A.C.; Hendrickson, J.R. Season of fire manipulates bud bank dynamics in northern mixed-grass prairie. Plant Ecol. 2015, 216, 835–846. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, G.I. Environmental control of bud and rhizome development in the seedling of Agropyron repens L. Beauv. Can. J. Bot. 1967, 67, 1315–1326. [Google Scholar] [CrossRef]
- Ott, J.P.; Hartnett, D.C. Bud bank dynamics and clonal growth strategy in the rhizomatous grass, Pascopyrum smithii. Plant Ecol. 2014, 216, 395–405. [Google Scholar] [CrossRef]
- Russell, M.L.; Vermeire, L.T.; Ganguli, A.C.; Hendrickson, J.R. Phenology of perennial, native grass, belowground axillary buds in the northern mixed-grass prairie. Am. J. Bot. 2017, 104, 915–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, B.-C.; Zhang, M.-X.; Alpert, P.; Lei, G.-C.; Yu, F.-H. Effects of Orientation on Survival and Growth of Small Fragments of the Invasive, Clonal Plant Alternanthera philoxeroides. PLoS ONE 2010, 5, e13631. [Google Scholar] [CrossRef] [PubMed]
- Hodoki, Y.; Ohbayashi, K.; Kunii, H. Analysis of population clonal diversity using microsatellite markers in the salt marsh sedge Carex scabrifolia in western Japan. Landsc. Ecol. Eng. 2013, 10, 9–15. [Google Scholar] [CrossRef]
- Ikegami, M.; Whigham, D.; Werger, M.J.A. Responses of rhizome length and ramet production to resource availability in the clonal sedge Scirpus olneyi A. Gray. Plant Ecol. 2006, 189, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Dalgleish, H.J.; Hartnett, D.C. Below-ground bud banks increase along a precipitation gradient of the North American Great Plains: A test of the meristem limitation hypothesis. New Phytol. 2006, 171, 81–89. [Google Scholar] [CrossRef]
- Li, H.Y.; Yan, Y.F. Bud banks of two perennial grasses: Composition, size, dynamics and contribution to population maintenance during the flooded restoration succession on the Songnen Meadow, China. Afr. J. Agric. Res. 2011, 6, 2198–2203. [Google Scholar]
- Lü, X.-T.; Dijkstra, F.A.; Kong, D.-L.; Wang, Z.-W.; Han, X.-G. Plant nitrogen uptake drives responses of productivity to nitrogen and water addition in a grassland. Sci. Rep. 2014, 4, 4817. [Google Scholar] [CrossRef] [Green Version]
- Benson, E.J.; Hartnett, D.C.; Mann, K.H. Belowground bud banks and meristem limitation in tallgrass prairie plant populations. Am. J. Bot. 2004, 91, 416–421. [Google Scholar] [CrossRef]
- Benson, E.J.; Hartnett, D.C. The Role of Seed and Vegetative Reproduction in Plant Recruitment and Demography in Tallgrass Prairie. Plant Ecol. 2006, 187, 163–178. [Google Scholar] [CrossRef]
- Ding, X.; Su, P.; Zhou, Z.; Shi, R. Belowground Bud Bank Distribution and Aboveground Community Characteristics along Different Moisture Gradients of Alpine Meadow in the Zoige Plateau, China. Sustainability 2019, 11, 2602. [Google Scholar] [CrossRef] [Green Version]
- Connolly, J.; Wayne, P. Assessing determinants of community biomass composition in two-species plant competition studies. Oecologia 2004, 142, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Nie, K.; Zou, X.; Ji, S.; Jiang, Z.; Li, G. Clonal growth response of Hippophae rhamnoides ssp. sinensis to irrigation intensity and its hormone regulation mechanism. Acta Ecol. Sin. 2018, 38, 4942–4952. [Google Scholar]
- Amiaud, B.; Touzard, B.; Bonis, A.; Bouzillé, J.-B. After grazing exclusion, is there any modification of strategy for two guerrilla species: Elymus repens (L.) Gould and Agrostis stolonifera (L.)? Plant Ecol. 2007, 197, 107–117. [Google Scholar] [CrossRef]







| Source | df | Total | ASB | RB | PD | ||||
|---|---|---|---|---|---|---|---|---|---|
| f | p | f | p | f | p | f | p | ||
| T | 1 | 76.32 | 0.000 *** | 855.47 | 0.000 *** | 167.86 | 0.000 *** | 241.77 | 0.000 *** |
| N | 1 | 92.26 | 0.000 *** | 123.00 | 0.000 *** | 15.14 | 0.000 *** | 109.85 | 0.000 *** |
| I | 1 | 78.74 | 0.000 *** | 193.37 | 0.000 *** | 0.20 | ns | 49.49 | 0.000 *** |
| T × N | 1 | 12.44 | 0.011 * | 37.01 | 0.000 *** | 18.58 | 0.000 *** | 1.90 | ns |
| T × I | 1 | 12.44 | 0.001 *** | 92.16 | 0.000 *** | 11.08 | 0.002 ** | 0.62 | ns |
| N × I | 1 | 0.74 | ns | 11.99 | 0.001 *** | 3.16 | ns | 1.84 | ns |
| T × N × I | 1 | 6.81 | 0.012 * | 11.59 | 0.001 *** | 0.52 | ns | 0.43 | ns |
| Total | 64 | ||||||||
| Bud Type | Controlling Factors | Parameters | Spring | Autumn | ||||
|---|---|---|---|---|---|---|---|---|
| Explanatory Power | R2 | p | Explanatory Power | R2 | p | |||
| Total | SE | TN | 0.019 | 0.13 | 0.685 | <0.001 | 0.00 | 0.998 |
| TP | 0.626 | 0.80 | 0.002 ** | 0.336 | 0.54 | 0.071 | ||
| AN | 0.730 | 0.85 | 0.000 *** | 0.838 | 0.91 | 0.000 *** | ||
| AP | 0.735 | 0.83 | 0.001 ** | 0.534 | 0.73 | 0.007 ** | ||
| SM | 0.162 | 0.37 | 0.241 | 0.543 | 0.72 | 0.008 ** | ||
| PT | RHN | 0.679 | 0.80 | 0.002 ** | 0.557 | 0.73 | 0.008 ** | |
| RHL | 0.567 | 0.74 | 0.006 ** | 0.754 | 0.86 | 0.000 *** | ||
| PI | 0.683 | 0.80 | 0.001 ** | 0.817 | 0.90 | 0.001 ** | ||
| ASB | SE | TN | 0.001 | 0.03 | 0.937 | 0.014 | 0.09 | 0.786 |
| TP | 0.615 | 0.78 | 0.003 ** | 0.305 | 0.48 | 0.116 | ||
| AN | 0.788 | 0.90 | 0.000 *** | 0.832 | 0.90 | 0.000 *** | ||
| AP | 0.810 | 0.85 | 0.001 ** | 0.499 | 0.71 | 0.009 ** | ||
| SM | 0.153 | 0.28 | 0.371 | 0.644 | 0.76 | 0.004 ** | ||
| PT | RHN | 0.646 | 0.77 | 0.003 ** | 0.542 | 0.71 | 0.010 * | |
| RHL | 0.585 | 0.78 | 0.003 ** | 0.788 | 0.87 | 0.000 *** | ||
| PI | 0.764 | 0.87 | 0.010 * | 0.808 | 0.90 | 0.000 *** | ||
| RB | SE | TN | 0.036 | 0.17 | 0.588 | 0.224 | 0.46 | 0.136 |
| TP | 0.525 | 0.72 | 0.008 ** | 0.014 | 0.16 | 0.620 | ||
| AN | 0.566 | 0.73 | 0.007 ** | 0.044 | 0.20 | 0.544 | ||
| AP | 0.577 | 0.72 | 0.008 ** | 0.011 | 0.09 | 0.777 | ||
| SM | 0.152 | 0.37 | 0.234 | 0.186 | 0.42 | 0.171 | ||
| PT | RHN | 0.570 | 0.72 | 0.008 ** | 0.005 | 0.07 | 0.827 | |
| RHL | 0.448 | 0.63 | 0.027 * | 0.086 | 0.29 | 0.355 | ||
| PI | 0.520 | 0.68 | 0.001 ** | 0.053 | 0.23 | 0.499 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, D.; Zheng, X.; Mu, C.; Wang, J. Irrigation and Nitrogen Application Promote Population Density through Altered Bud Bank Size and Components in Leymus chinensis. Agronomy 2022, 12, 1436. https://doi.org/10.3390/agronomy12061436
Yu D, Zheng X, Mu C, Wang J. Irrigation and Nitrogen Application Promote Population Density through Altered Bud Bank Size and Components in Leymus chinensis. Agronomy. 2022; 12(6):1436. https://doi.org/10.3390/agronomy12061436
Chicago/Turabian StyleYu, Dafu, Xingyu Zheng, Chunsheng Mu, and Junfeng Wang. 2022. "Irrigation and Nitrogen Application Promote Population Density through Altered Bud Bank Size and Components in Leymus chinensis" Agronomy 12, no. 6: 1436. https://doi.org/10.3390/agronomy12061436