
Identification and Genetic Mapping of Potential QTLs Conferring Heat Tolerance in Cotton (Gossypium hirsutum L.) by Using Micro Satellite Marker’s Approach

Abstract
:1. Introduction
2. Materials and Methods
2.1. Heat Stress Estimation
2.2. Parental Lines Screening
2.3. Mapping Population
2.4. Phenotypic Data Collection Statistical Analysis
2.5. Microsatellite Analysis
2.6. QTL Mapping
3. Results
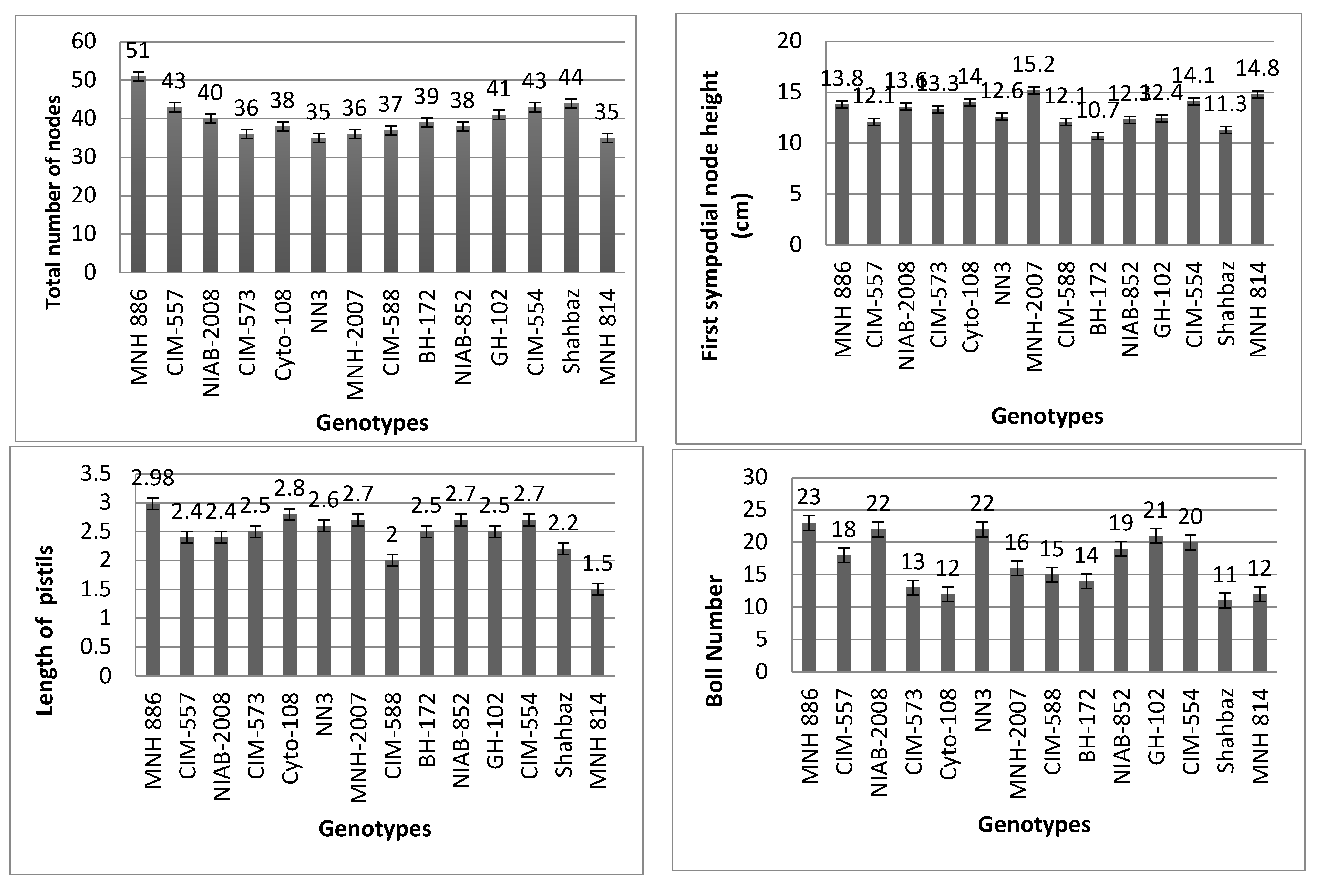
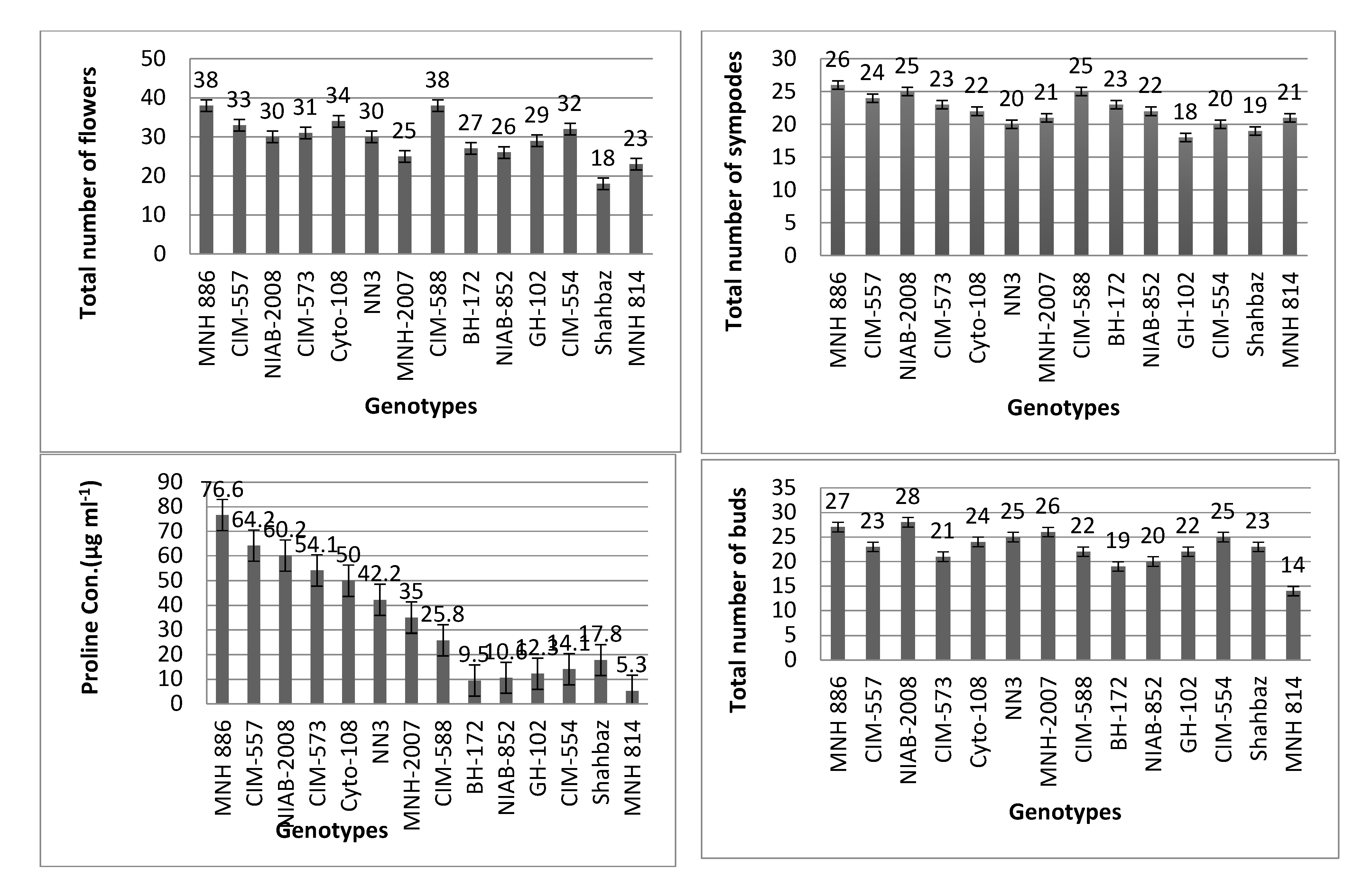
3.1. Average Performance of Cotton Varieties Based on Morpho-Physiological Traits
3.2. Stress Determining Physiological Traits
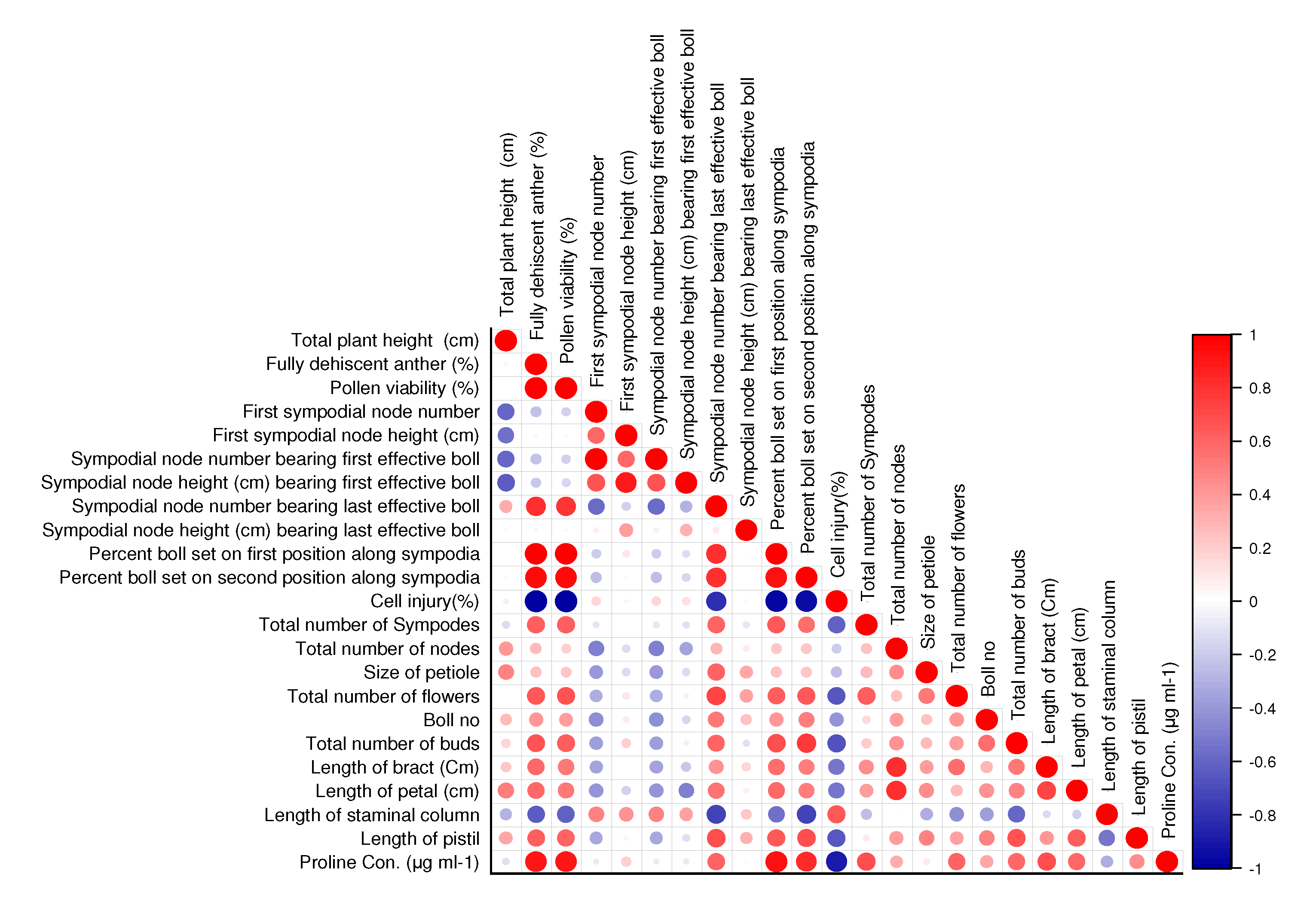
3.3. Correlation
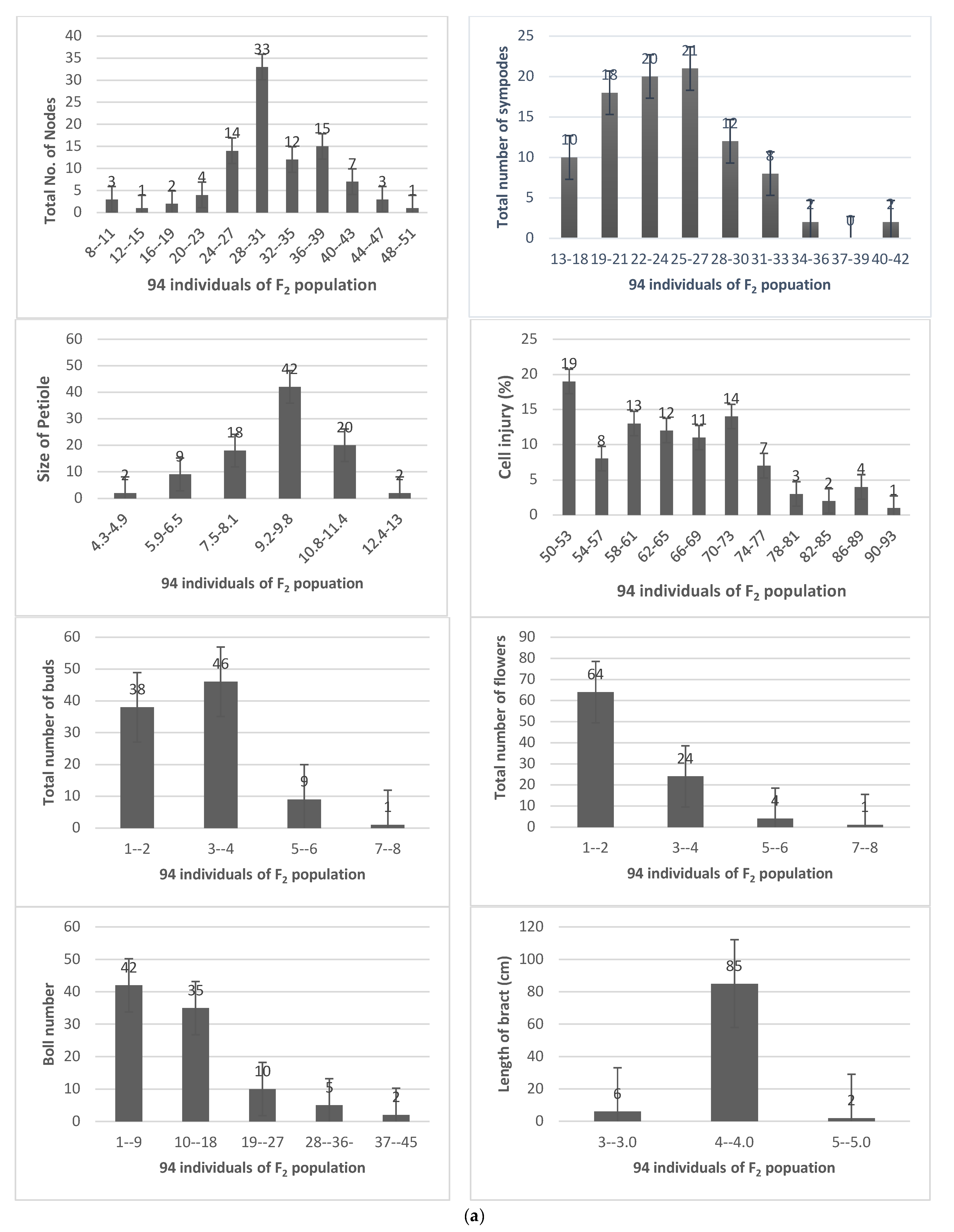
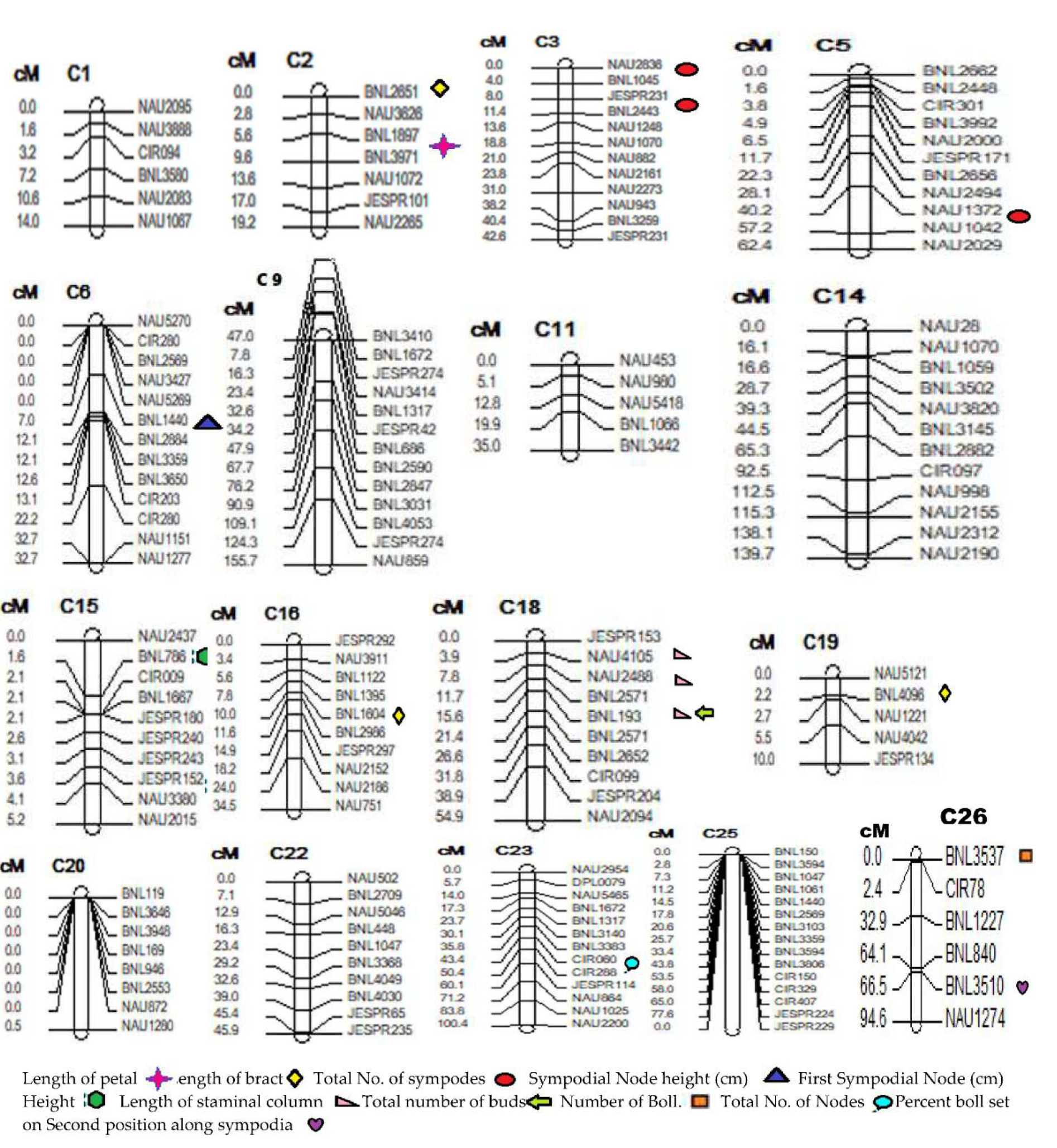
3.4. Construction and Characterization of Intra Specific Linkage Map
3.5. QTLs Mapping for Traits Associated with Heat Tolerance in Cotton
3.5.1. QTLs for First Sympodial Node Height (FSH)
3.5.2. QTLs for Sympodial Node Height Bearing First Effective Boll Set (SNH)
3.5.3. QTLs for Percent Boll Set on Second Position along Sympodia (PBS)
3.5.4. QTLs for Total No of Sympodes (TNS)
3.5.5. QTL for Total No of Nodes (TNN)
3.5.6. QTLs for Number of Bolls (NOB)
3.5.7. QTLs for Total No of Buds (TNB)
3.5.8. QTLs for Length of Bract (LOB)
3.5.9. QTL for Length of Staminal Column (LOS)
3.5.10. QTLs for Length of Petal (LOP)
4. Discussion
5. Conclusions
6. Future Recommendation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IVAN Study Investigators; Chakravarthy, U.; Harding, S.P.; Rogers, C.A.; Downes, S.M.; Lotery, A.J.; Wordsworth, S.; Reeves, B.C. Ranibizumab versus bevacizumab to treat neovascular age-related macular degeneration: One-year findings from the IVAN randomized trial. Ophthalmology 2012, 119, 1399–1411. [Google Scholar] [CrossRef] [PubMed]
- Dabbert, T.A.; Gore, M.A. Challenges and perspectives on improving heat and drought stress resilience in cotton. J. Cotton Sci. 2014, 18, 393–409. [Google Scholar]
- Chapagain, A.K.; Hoekstra, A.Y.; Savenije, H.H.; Gautam, R. The water footprint of cotton consumption: An assessment of the impact of worldwide consumption of cotton products on the water resources in the cotton producing countries. Ecol. Econ. 2006, 60, 186–203. [Google Scholar] [CrossRef]
- Ashraf, M. Inducing drought tolerance in plants: Recent advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Wendel, J.F.; Brubaker, C.; Alvarez, I.; Cronn, R.; Stewart, J.M. Evolution and natural history of the cotton genus. In Genetics and Genomics of Cotton; Springer: New York, NY, USA, 2009; pp. 3–22. [Google Scholar]
- Grover, C.E.; Zhu, X.; Grupp, K.K.; Jareczek, J.J.; Gallagher, J.P.; Szadkowski, E.; Seijo, J.G.; Wendel, J.F. Molecular confirmation of species status for the allopolyploid cotton species, Gossypium ekmanianum Wittmack. Genet. Resour. Crop Evolut. 2015, 62, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Senchina, D.S.; Alvarez, I.; Cronn, R.C.; Liu, B.; Rong, J.; Noyes, R.D.; Wendel, J.F. Rate variation among nuclear genes and the age of polyploidy in Gossypium. Mol. Biol. Evol. 2003, 20, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Cronn, R.; Wendel, J.F. Cryptic trysts, genomic mergers, and plant speciation. New Phytol. 2004, 161, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Banuri, T. Pakistan: Environmental impact of cotton production and trade. Int. Inst. Sustain. Dev. 1998, 161. [Google Scholar]
- Janjua, P.Z.; Samad, G.; Khan, N.U.; Nasir, M. Impact of climate change on wheat production: A case study of Pakistan [with comments]. Pak. Dev. Rev. 2010, 49, 799–822. [Google Scholar] [CrossRef] [Green Version]
- Riaz, M.; Farooq, J.; Sakhawat, G.; Mahmood, A.; Sadiq, M.A.; Yaseen, M. Genotypic variability for root/shoot parameters under water stress in some advanced lines of cotton (Gossypium hirsutum L.). Genet. Mol. Res. 2013, 12, 552–561. [Google Scholar] [CrossRef]
- Farooq, J.; Anwar, M.; Rizwan, M.; Riaz, M.; Mahmood, K.; Mahpara, S. Estimation of correlation and path analysis of various yield and related parameters in cotton (Gossypium hirsutum L.). Cotton Genom. Genet. 2015, 6, 1–6. [Google Scholar]
- Khan, N.; Faqir, M.A.; Khan, A.A.; Rashid, A. Measurement of canopy temperature for heat tolerance in upland cotton: Variability and its genetic basis. Pak. J. Agri. Sci. 2014, 51, 359–365. [Google Scholar]
- Reddy, A.R.; Reddy, K.R.; Padjung, R.; Hodges, H.F. Nitrogen nutrition and photosynthesis in leaves of Pima cotton. J. Plant Nutr. 1996, 19, 755–770. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Sarto, M.V.M.; Rocha, K.F.; Martins, J.D.L.; Alves, M.S. Does the introgression of BT gene affect physiological cotton response to water deficit? Planta Daninha 2019, 37, 1–7. [Google Scholar] [CrossRef]
- Jarwar, A.H.; Wang, X.; Iqbal, M.S.; Sarfraz, Z.; Wang, L.; Ma, Q.; Shuli, F. Genetic divergence on the basis of principal component, correlation and cluster analysis of yield and quality traits in cotton cultivars. Pak. J. Bot 2019, 51, 1143–1148. [Google Scholar] [CrossRef]
- Wang, M.; Tu, L.; Yuan, D.; Zhu, D.; Shen, C.; Li, J.; Liu, F.; Pei, L.; Wang, P.; Zhao, G.; et al. Reference genome sequences of two cultivated allotetraploid cottons, Gossypium hirsutum and Gossypium barbadense. Nat. Genet. 2019, 51, 224–229. [Google Scholar] [CrossRef] [Green Version]
- Tester, M.; Langridge, P. Breeding technologies to increase crop production in a changing world. Science 2010, 327, 818–822. [Google Scholar] [CrossRef]
- Swinnen, J.; Vandeplas, A. Rich consumers and poor producers: Quality and rent distribution in global value chains. J. Glob. Dev. 2012, 2, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Song, X.; Han, Z.; Guo, W.; John, Z.Y.; Sun, J.; Pan, J.; Kohel, R.J.; Zhang, T. Complete assignment of the chromosomes of Gossypium hirsutum L. by translocation and fluorescence in situ hybridization mapping. Theor. Appl. Genet. 2006, 113, 73–80. [Google Scholar] [CrossRef]
- Park, Y.-H.; Alabady, M.S.; Ulloa, M.; Sickler, B.; Wilkins, T.A.; Yu, J.; Stelly, D.; Kohel, R.J.; El-Shihy, O.M.; Cantrell, R.G. Genetic mapping of new cotton fiber loci using EST-derived microsatellites in an interspecific recombinant inbred line cotton population. Mol. Genet. Genom. 2005, 274, 428–441. [Google Scholar] [CrossRef]
- Li, H.; Pan, Z.; He, S.; Jia, Y.; Geng, X.; Chen, B.; Wang, L.; Pang, B.; Du, X. QTL mapping of agronomic and economic traits for four F2 populations of upland cotton. J. Cotton Res 2021, 4, 1–12. [Google Scholar] [CrossRef]
- Tyagi, P.; Gore, M.A.; Bowman, D.T.; Campbell, B.T.; Udall, J.A.; Kuraparthy, V. Genetic diversity and population structure in the US Upland cotton (Gossypium hirsutum L.). Theor. Appl. Genet. 2014, 127, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Stewart, J.M. Economical and rapid method for extracting cotton genomic DNA. J. Cotton Sci. 2000, 4, 193–201. [Google Scholar]
- Iqbal, M.; Ul-Allah, S.; Naeem, M.; Ijaz, M.; Sattar, A.; Sher, A. Response of cotton genotypes to water and heat stress: From field to genes. Euphytica 2017, 213, 1–11. [Google Scholar] [CrossRef]
- Akhtar, K.P.; Ullah, R.; Khan, I.A.; Saeed, M.; Sarwar, N.; Mansoor, S. First symptomatic evidence of infection of Gossypium arboreum with Cotton leaf curl Burewala virus through grafting. Int. J. Agric. Biol. 2013, 15, 157–160. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Li, F.; Fan, G.; Lu, C.; Xiao, G.; Zou, C.; Kohel, R.J. Genome sequence of cultivated Upland cotton (Gossypium hirsutum TM-1) provides insights into genome evolution. Nat. Biotechnol. 2015, 33, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Hu, Y.; Jiang, W.; Fang, L.; Guan, X.; Chen, J.; Zhang, J.; Saski, C.A.; Scheffler, B.E.; Stelly, D.M.; et al. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat. Biotechnol. 2015, 33, 531–537. [Google Scholar] [CrossRef] [Green Version]
- Said, J.I.; Lin, Z.; Zhang, X.; Song, M.; Zhang, J. A comprehensive meta QTL analysis for fiber quality, yield, yield related and morphological traits, drought tolerance, and disease resistance in tetraploid cotton. BMC Genom. 2013, 14, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Said, J.I.; Knapka, J.A.; Song, M.; Zhang, J. Cotton QTLdb: A cotton QTL database for QTL analysis, visualization, and comparison between Gossypium hirsutum and G. hirsutum × G. barbadense populations. Mol. Genet. Genom. 2015, 290, 1615–1625. [Google Scholar] [CrossRef]
- Burke, H.R.; Clark, W.E.; Cate, J.R.; Fryxell, P.A. Origin and dispersal of the boll weevil. Bull. ESA 1986, 32, 228–238. [Google Scholar] [CrossRef]
- Stewart, J.M.; Hsu, C.L. In-ovulo embryo culture and seedling development of cotton (Gossypium hirsutum L.). Planta 1977, 137, 113–117. [Google Scholar] [CrossRef]
- Barnett, N.M.; Naylor, A.W. Amino acid and protein metabolism in Bermuda grass during water stress. Plant Physiol. 1966, 41, 1222–1230. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldem, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Paterson, A.H.; Brubaker, C.L.; Wendel, J.F. A rapid method for extraction of cotton (Gossypium spp.) genomic DNA suitable for RFLP or PCR analysis. Plant Mol. Biol. Report. 1993, 11, 122–127. [Google Scholar] [CrossRef]
- Reddy, O.U.K.; Pepper, A.E.; Abdurakhmonov, I.; Saha, S.; Jenkins, J.N.; Brooks, T.; El-Zik, K.M. New Dinucleotide and Trinucleotide Microsatellite Marker Resources for Cotton Genome Research. J. Cotton Sci. 2001, 5, 103–113. [Google Scholar]
- Nguyen, T.-B.; Giband, M.; Brottier, P.; Risterucci, A.-M.; Lacape, J.-M. Wide coverage of the tetraploid cotton genome using newly developed microsatellite markers. Theor. Appl. Genet. 2004, 109, 167–175. [Google Scholar] [CrossRef]
- Han, Z.G.; Guo, W.Z.; Song, X.L.; Zhang, T.Z. Genetic mapping of EST-derived microsatellites from the diploid Gossypium arboreum in allotetraploid cotton. Mol. Genet. Genom. 2004, 272, 308–327. [Google Scholar] [CrossRef]
- Han, Z.; Wang, C.; Song, X.; Guo, W.; Gou, J.; Li, C. Characteristics, development and mapping of Gossypium hirsutum derived EST-SSRs in allotetraploid cotton. Theor. Appl. Genet. 2006, 112, 430–439. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, W.; Zhang, T. Molecular linkage map of allotetraploid cotton (Gossypium hirsutum L. × Gossypium barbadense L.) with a haploid population. Theor. Appl. Genet. 2002, 105, 1166–1174. [Google Scholar]
- Tanksley, S.D.; Young, N.D.; Paterson, A.H.; Bonierbale, M.W. RFLP mapping in plant breeding: New tool for an old science. Biotechnology 1989, 7, 257–264. [Google Scholar] [CrossRef]
- Soller, M.; Brody, T.; Genizi, A. On the power of experimental designs for the detection of linkage between marker loci and quantitative loci in crosses between inbred lines. Theor. Appl. Genet. 1976, 47, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S.; Botstein, D. Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics 1989, 121, 185–199. [Google Scholar] [CrossRef]
- Knapp, T.R. Treating ordinal scales as interval scales: An attempt to resolve the controversy. Nurs. Res. 1990, 39, 121–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haley, C.S.; Knott, S.A. A simple regression method for mapping quantitative trait loci in line crosses using flanking markers. Heredity 1992, 69, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Slocum, M.; Osborn, T. Molecular marker analysis of genes controlling morphological variation in Brassica rapa (syn. campestris). Theor. Appl. Genet. 1995, 90, 1–10. [Google Scholar] [CrossRef]
- Kosambi, D.D. The estimation of a map distance from recombination values. Ann. Eugen. 1994, 12, 172–175. [Google Scholar] [CrossRef]
- Larntz, K. Small-sample comparisons of exact levels for chi-squared goodness-of-fit statistics. J. Am. Stat. Assoc. 1978, 73, 253–263. [Google Scholar] [CrossRef]
- Mutschler, M.A.; Doerge, R.W.; Liu, S.C.; Kuai, J.P.; Liedl, B.E.; Shapiro, J.A. QTL analysis of pest resistance in the wild tomato Lycopersicon pennellii: QTLs controlling acylsugar level and composition. Theor. Appl. Genet. 1996, 92, 709–718. [Google Scholar] [CrossRef]
- Baloch, M.J.; Veesar, N.F. Identification of plant traits for characterization of early maturing upland cotton varieties. Biol. Sci.-PJSIR 2007, 50, 128–132. [Google Scholar]
- Reddy, K.R.; Hodges, H.F.; Reddy, V.R. Temperature effects on cotton fruit retention. Agron. J. 1992, 84, 26–30. [Google Scholar] [CrossRef]
- Azhar, F.M.; Ali, Z.; Akhtar, M.M.; Khan, A.A.; Trethowan, R. Genetic variability of heat tolerance, and its effect on yield and fibre quality traits in upland cotton (Gossypium hirsutum L.). Plant Breed. 2009, 128, 356–362. [Google Scholar] [CrossRef]
- Hussain, M.; Azhar, F.M.; Khan, A.A. Genetics of inheritance and correlations of some morphological and yield contributing traits in upland cotton. Pak. J. Bot. 2009, 41, 2975–2986. [Google Scholar]
- Wang, D. RFLP Mapping, QTL Identification, and Cytogenetic Analysis in Sour Cherry; Michigan State University: East Lansing, MI, USA, 1998. [Google Scholar]
- Siddique, M.R.B.; Hamid, A.I.M.S.; Islam, M.S. Drought stress effects on water relations of wheat. Bot. Bull. Acad. Sin. 2000, 41, 35–39. [Google Scholar]
- Baloch, A.; Rind, A.; Jamali, K. Genetic maps and marker assisted selection for major gene traits in rice. Pak. J. Biotechnol. 2004, 1, 33–46. [Google Scholar]
- Parida, A.K.; Dagaonkar, V.S.; Phalak, M.S.; Umalkar, G.V.; Aurangabadkar, L.P. Alterations in photosynthetic pigments, protein and osmotic components in cotton genotypes subjected to short-term drought stress followed by recovery. Plant Biotechnol. Rep. 2007, 1, 37–48. [Google Scholar] [CrossRef]
- Guo, W.; Cai, C.; Wang, C.; Han, Z.; Song, X.; Wang, K.; Niu, X.; Wang, C.; Lu, K.; Shi, B.; et al. A microsatellite-based, gene-rich linkage map reveals genome structure, function and evolution in Gossypium. Genetics 2007, 176, 527–541. [Google Scholar] [CrossRef] [Green Version]
- Malik, R.S.; Dhankar, J.S.; Turner, N.C. Influence of soil water deficits on root growth of cotton seedlings. Plant Soil 1979, 53, 109–115. [Google Scholar] [CrossRef]
- Baloch, A.W.; Solangi, A.M.; Baloch, M.; Baloch, G.M.; Abro, S. Estimation of heterosis and heterobeltiosis for yield and fiber traits in F1 hybrids of upland cotton (Gossypium hirsutum L.) genotypes. Pak. J. Agri. Agril. Engg. Vet. Sci. 2015, 31, 221–228. [Google Scholar]
- Peng, M.W. Global Strategy. Cengage Learning; Cengage 200 Pier 4 Boulevard: Boston, MA, USA, 2021. [Google Scholar]
- Raison, J.K.; Berry, J.A.; Armond, P.A.; Pike, C.S. Membrane properties in relation to the adaptation of plants to temperature stress. In Adaptation of Plants to Water and High Temperature Stress; Turner, N.C., Kramer, P.J., Eds.; John Wiley and Sons: New York, NY, USA, 1980; pp. 261–273. [Google Scholar]
- Kushanov, F.N.; Turaev, O.S.; Ernazarova, D.K.; Gapparov, B.M.; Oripova, B.B.; Kudratova, M.K.; Abdurakhmonov, I.Y. Genetic diversity, QTL mapping and MAS technology in cotton (Gossypium spp.). Front. Plant Sci. 2021, 12, 29–71. [Google Scholar] [CrossRef]
- Iqbal, M.; Naeem, M.; Rizwan, M.; Nazeer, W.; Shahid, M.Q.; Aziz, U.; Aslam, T.; Ijaz, M. Studies of genetic variation for yield related traits in upland cotton. Am. Eurasian J. Agric. Environ. Sci. 2013, 13, 611–618. [Google Scholar]
- Gipson, J.R.; Joham, H.E. Influence of night temperature on growth and development of cotton (Gossypium hirsutum L.). III. Fiber elongation. Crop Sci. 1969, 9, 127–129. [Google Scholar] [CrossRef]
- Odongo, I.; Ssemambo, R.; Kungu, J.M. Prevalence of Escherichia Coli and its antimicrobial susceptibility profiles among patients with UTI at Mulago Hospital, Kampala, Uganda. Interdiscip. Perspect. Infect. Dis. 2020, 2020, 8042540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.; Yasmin, T.; Tabassum, N.; Ullah, I.; Asif, M.; Zafar, Y. Studying the extent of genetic diversity among Gossypium arboreum L. genotypes/cultivars using DNA fingerprinting. Genet. Resour. Crop Evol. 2008, 55, 331–339. [Google Scholar] [CrossRef]
- Shaheen, T.; Zafar, Y.; Rahman, M.-U. QTL mapping of some productivity and fibre traits in Gossypium arboreum. Turk. J. Bot. 2013, 37, 802–810. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Air Temperature (°C) | Relative Humidity | Rainfall (mm) | Evapotranspiration (cm Day) | Soil Temperature (°C) | |||
|---|---|---|---|---|---|---|---|---|
| Max | Min | Max | Min | 5 cm | 10 cm | |||
| January | 5.3 | 19.1 | 63 | 92 | 1.5 | 0.24 | 9.4 | 10.5 |
| February | 6.9 | 20.5 | 52 | 76 | 0.0 | 0.39 | 12.3 | 12.7 |
| March | 13.9 | 27.4 | 45 | 65 | 0.0 | 0.67 | 19.2 | 19.7 |
| April | 20.6 | 32.8 | 55 | 72 | 24.7 | 0.86 | 26.5 | 27.0 |
| May | 25.7 | 39.4 | 54 | 57 | 1.10 | 1.22 | 31.7 | 32.0 |
| June | 28.6 | 39.4 | 58 | 64 | 0.0 | 1.26 | 35.4 | 35.4 |
| July | 28.8 | 38.1 | 61 | 73 | 16.9 | 1.11 | 35.8 | 36.0 |
| August | 28.0 | 35.6 | 72 | 76 | 16.1 | 0.84 | 34.9 | 35.1 |
| September | 25.7 | 33.1 | 80 | 87 | 167.0 | 0.59 | 29.8 | 30.2 |
| October | 18.9 | 31.7 | 62 | 83 | 3.2 | 0.48 | 24.3 | 25.1 |
| November | 13.1 | 26.8 | 81 | 87 | 0.0 | 0.28 | 17.7 | 18.6 |
| December | 7.8 | 21.9 | 80 | 87 | 4.0 | 0.19 | 12.8 | 13.8 |
| Cultivars | Total Plant Height (cm) | Fully Dehiscent Anther (%) | Pollen Viability (%) | First Sympodial Node Number | First Sympodial Node Height (cm) | Sympodial Node Number Bearing First Effective Boll | Sympodial Node Height (cm) Bearing First Effective Boll | Sympodial Node Number Bearing Last Effective Boll | Sympodial Node Height (cm) Bearing Last Effective Boll | Percent Boll Set on First Position along Sympodia | Percent Boll Set on Second Position along Sympodia | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MNH 886 | 90 | 92 | 88.3 | 7 | 13.8 | 8 | 15 | 34 | 114.2 | 51 | 32 | |
| CIM-557 | 67 | 91 | 87.4 | 7 | 12.1 | 8 | 14.8 | 32 | 93.4 | 49 | 31 | |
| NIAB-2008 | 55 | 89 | 86.1 | 7 | 13.6 | 8 | 15.2 | 33 | 82.5 | 50 | 32 | |
| CIM-573 | 54 | 88 | 85.8 | 8 | 13.3 | 9 | 15.9 | 31 | 112.3 | 48 | 31 | |
| Cyto-108 | 67 | 87 | 85.1 | 7 | 14 | 8 | 16.3 | 33 | 108.9 | 49 | 30 | |
| NN3 | 88 | 85 | 83.5 | 7 | 12.6 | 8 | 14.5 | 32 | 105.6 | 47 | 31 | |
| MNH-2007 | 67 | 83 | 82.2 | 8 | 15.2 | 9 | 17.9 | 31 | 87.3 | 47 | 30 | |
| CIM-588 | 77 | 82 | 80.5 | 7 | 12.1 | 8 | 15.6 | 33 | 92.7 | 45 | 29 | |
| BH-172 | 80 | 80 | 79.3 | 7 | 10.7 | 8 | 13.8 | 32 | 84.5 | 44 | 29 | |
| NIAB-852 | 85 | 79 | 77.5 | 7 | 12.3 | 8 | 14.9 | 33 | 113.8 | 45 | 28 | |
| GH-102 | 77 | 77 | 76.3 | 7 | 12.4 | 8 | 15.3 | 31 | 103.2 | 43 | 27 | |
| CIM-554 | 66 | 75 | 74.1 | 7 | 14.1 | 8 | 16.8 | 31 | 121.2 | 42 | 29 | |
| Shahbaz | 88 | 73 | 71.2 | 7 | 11.3 | 8 | 13.7 | 29 | 81.7 | 41 | 26 | |
| MNH 814 | 54 | 64 | 66.4 | 8 | 14.8 | 9 | 17.3 | 27 | 106.5 | 38 | 23 | |
| Max | 90 | 92 | 88 | 8 | 15 | 9 | 17 | 34 | 121 | 51 | 32 | |
| Min | 54 | 64 | 66 | 7 | 10 | 8 | 13 | 27 | 81 | 38 | 23 | |
| Variance | 219 | 60 | 42 | 181 | 1.70 | 181 | 1.52 | 3.34 | 176 | 14 | 6 | |
| Std. Dev. | ±14.79 | ±7.80 | ±6.53 | ±42 | ±1.30 | ±42 | ±1.23 | ±1.82 | ±13.2 | ±3.75 | ±2.50 | |
| Cultivars | Cell Injury (%) | Total Number of Sympodes | Total Number of Nodes | Size of Petiole | Total Number of Flowers | Number of Bolls | Total Number of Buds | Length of Bract (cm) | Length of Petal (cm) | Length of Staminal Column | Length of Pistil | Proline Con. (μg mL−1) |
| MNH-886 | 65 | 26 | 51 | 9.3 | 38 | 23 | 27 | 5 | 4 | 2.90 | 2.98 | 76.7 |
| CIM-557 | 66 | 24 | 43 | 7.3 | 33 | 18 | 23 | 4.5 | 2.9 | 2.4 | 2.4 | 64.2 |
| NIAB-2008 | 67 | 25 | 40 | 7.6 | 30 | 22 | 28 | 3.6 | 2.8 | 2.3 | 2.4 | 60.2 |
| CIM-573 | 67 | 23 | 36 | 8.1 | 31 | 13 | 21 | 3.7 | 2.6 | 2.6 | 2.5 | 54.1 |
| Cyto-108 | 68 | 22 | 38 | 8.3 | 34 | 12 | 24 | 3.9 | 2.7 | 2.7 | 2.8 | 50 |
| NN3 | 68 | 20 | 35 | 7.5 | 30 | 22 | 25 | 3.4 | 2.6 | 2.1 | 2.6 | 42.2 |
| MNH-2007 | 68 | 21 | 36 | 7.8 | 25 | 16 | 26 | 3.3 | 2.5 | 2.5 | 2.7 | 35 |
| CIM-588 | 70 | 25 | 37 | 9.0 | 38 | 15 | 22 | 3.7 | 2.2 | 2 | 2 | 25.8 |
| BH-172 | 71 | 23 | 39 | 8.0 | 27 | 14 | 19 | 3.1 | 2.8 | 2.2 | 2.5 | 17.8 |
| NIAB-852 | 72 | 22 | 38 | 8.7 | 26 | 19 | 20 | 3.2 | 2.8 | 2.6 | 2.7 | 14.1 |
| GH-102 | 73 | 18 | 41 | 8.5 | 29 | 21 | 22 | 3.5 | 2.7 | 2.7 | 2.5 | 12.3 |
| CIM-554 | 74 | 20 | 43 | 8.3 | 32 | 20 | 25 | 3.8 | 2.4 | 2.5 | 2.7 | 10.6 |
| Shahbaz | 76 | 19 | 44 | 7.9 | 18 | 11 | 23 | 3.9 | 2.9 | 3 | 2.2 | 9.5 |
| MNH-814 | 80 | 21 | 35 | 7 | 23 | 12 | 14 | 3 | 2 | 4.5 | 1.5 | 5.3 |
| Max | 80 | 26 | 51 | 9 | 38 | 23 | 28 | 5 | 4 | 4 | 2 | 76 |
| Min | 65 | 18 | 35 | 7 | 18 | 11 | 14 | 3 | 2 | 2 | 1.5 | 5 |
| Variance | 18 | 5 | 19 | 0.41 | 27 | 19 | 13 | 0.29 | 0.20 | 0.36 | 0.13 | 56 |
| Std. Dev. | ±4.25 | ±2.40 | ±4.41 | ±0.64 | ±5.2 | ±4.4 | ±3.6 | ±0.54 | ±0.45 | ±0.60 | ±0.37 | ±23 |
| Population Size | Traits | Parents | F2 Population Statistical Data | |||||
|---|---|---|---|---|---|---|---|---|
| 94 | MNH-886 | MNH-814 | Max | Min | Mean | SD | Skew | |
| TPH | 90 | 54 | 48 | 105 | 74.37 | 12.07 | 0.390 | |
| FDA | 92 | 64 | 60 | 92. | 75.03 | 9.98 | 0.365 | |
| POV | 88.3 | 66.4 | 58 | 666 | 78.48 | 61.89 | 0.391 | |
| FSN | 7 | 8 | 7 | 9 | 7.82 | 0.824 | 0.328 | |
| FSH | 13.8 | 14.8 | 10 | 16.10 | 13.64 | 1.134 | −0.224 | |
| SNF | 8 | 9 | 6 | 11 | 8.64 | 0.912 | 0.934 | |
| SNH | 15 | 17.3 | 11.10 | 17.30 | 14.30 | 1.568 | 0.211 | |
| SNL | 34 | 27 | 22 | 34 | 28.92 | 3.26 | 0.002 | |
| SNB | 114.2 | 106.5 | 80.5 | 115 | 100.19 | 9.63 | −0.315 | |
| PBF | 51 | 38 | 31 | 51 | 43.44 | 5.30 | −0.210 | |
| PBS | 32 | 23 | 23 | 3191 | 62.47 | 326.15 | 0.691 | |
| CIY | 65 | 80 | 50 | 90 | 64.60 | 10.36 | 0.477 | |
| TNS | 26 | 21 | 13 | 39 | 21.77 | 5.33 | 0.732 | |
| TNN | 45 | 51 | 6 | 48 | 30.93 | 7.12 | −0.577 | |
| SOP | 9.3 | 7 | 4.30 | 12.30 | 8.69 | 1.51 | −0.428 | |
| TNF | 35 | 23 | 1 | 8 | 2.13 | 1.25 | 1.62 | |
| NOB | 23 | 12 | 1 | 45 | 12.10 | 8.80 | 1.26 | |
| TNB | 27 | 14 | 1 | 7 | 2.92 | 1.32 | 0.796 | |
| LOB | 5 | 3 | 3 | 5 | 3.94 | 0.33 | −0.770 | |
| LOP | 4 | 2 | 2.70 | 4 | 3.65 | 0.317 | −0.760 | |
| LOS | 2.90 | 4.5 | 2.0 | 3.10 | 2.64 | 0.261 | −0.434 | |
| LPI | 2.98 | 1.5 | 2.30 | 4.00 | 2.99 | 0.308 | 0.240 | |
| PCO | 5.3 | 76.6 | 5.20 | 76.7 | 33.43 | 27.05 | 0.274 | |
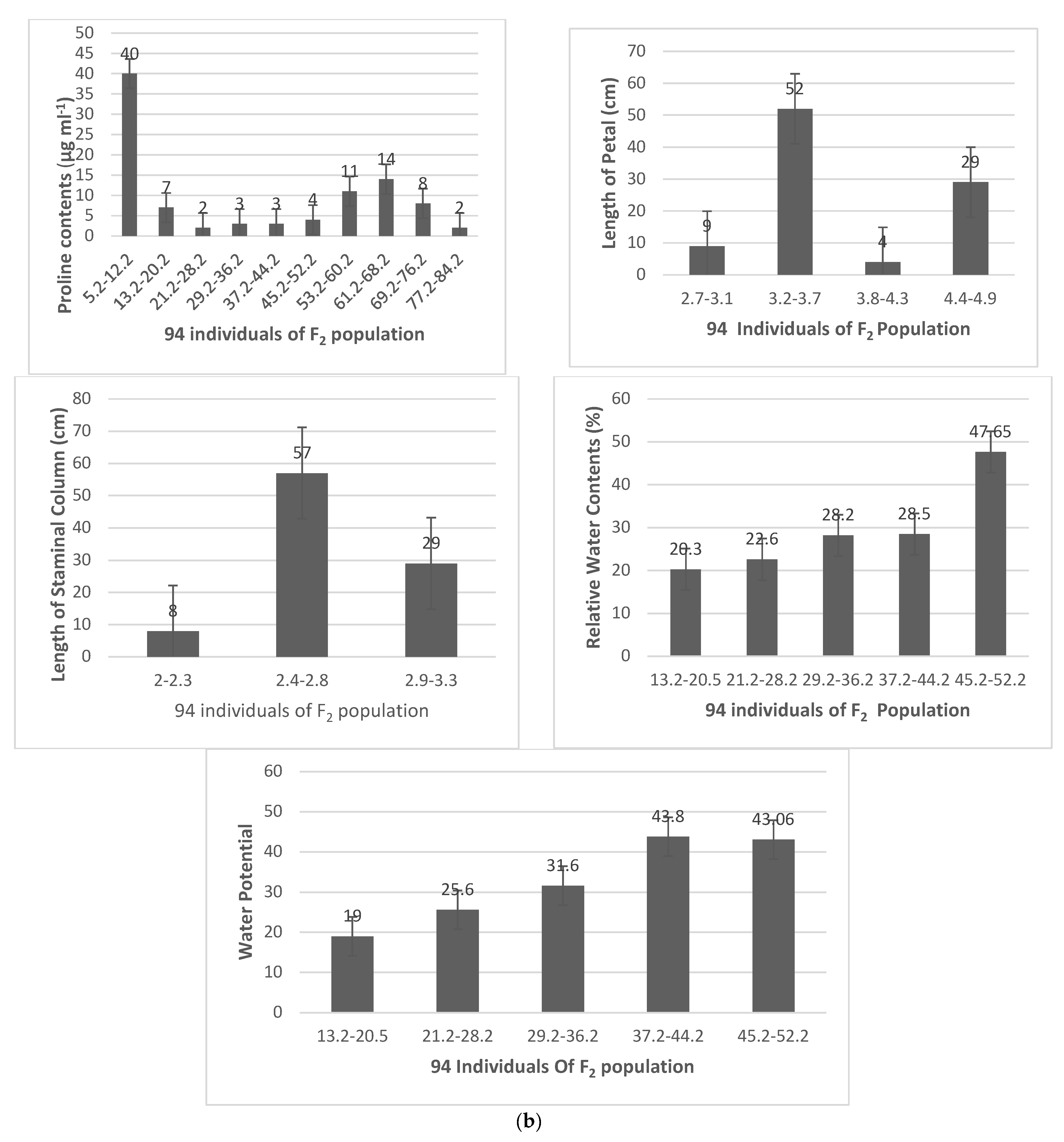
| Population Size | Traits | Parents (Means) | F2 Population Statistical Data | |||||
|---|---|---|---|---|---|---|---|---|
| 94 | MNH-886 | MNH-814 | Max | Min | Mean | SD | Skew | |
| RWC | 47.65 | 43.06 | 54.91 | 40.28 | 47.65308 | 3.815905 | 0.321 | |
| WP | 20.30 | 19.00 | 27 | 15 | 19.65 | 2.511 | 0.283 | |
| OP | 860.76 | 805.92 | 975 | 727 | 833.34 | 65.07 | 0.382 | |
| CIY | 65 | 80 | 50 | 90 | 64.60 | 10.36 | 0.477 | |
| PCO | 5.3 | 7.66 | 7.66 | 5.3 | 6.48 | 2.05 | 0.274 | |
| Item | Field Exp. Pop |
|---|---|
| Total no. of SSR loci | 175 |
| No. of mapped loci | 171 |
| No. of individuals | 94 |
| No. of linkage groups | 17 |
| No. of unlinked loci | 4 |
| Length of map (cm) | 4402.7 |
| Total no. of skewed loci | 24 |
| QTLs | Chr. No. | SSR Markers | LOD Value | Additive | Dominance | Dominance/Additive | PV% Age |
|---|---|---|---|---|---|---|---|
| First Sympodial Node Height (cm) | |||||||
| qFSHa1 | 15 | BNL786-CIR009 | 6.10 | 0.59 | −0.80 | −1.36 | 36.62 |
| qFSHa2 | 15 | JESPR152-NAU3380 | 6.09 | 0.58 | −0.81 | −1.39 | 35.98 |
| Sympodial Node Height (cm) | |||||||
| qSNH1 | 6 | BNL1440-BNL2884 | 3.42 | 0.77 | −0.31 | −0.40 | 17.59 |
| Percent Boll Set on Second Position Along Sympodia | |||||||
| qPBS1 | 26 | BNL3510-NAU1274 | 18.19 | 0.69 | 0.35 | 0.50 | 14.56 |
| Total No. of Sympodes | |||||||
| qTNSa1 | 03 | NAU2836-BNL1045 | 3.59 | 6.00 | 0.41 | 0.07 | 10.05 |
| qTNSa2 | 03 | JESPR231-BNL2443 | 3.71 | 6.27 | 0.38 | 0.06 | 10.12 |
| qTNSa3 | 05 | NAU1372-NAU1042 | 3.98 | 2.89 | −0.50 | −0.17 | 16.93 |
| Total No. of Nodes | |||||||
| qTNN1 | 23 | CIR080-CIR288 | 4.05 | 0.18 | 0.03 | 0.17 | 12.91 |
| Number of Bolls | |||||||
| qNOB1 | 26 | BNL3537-CIR078 | 3.80 | 4.25 | −3.15 | −0.74 | 21.52 |
| Total Number of Buds | |||||||
| qTNB1 | 18 | BNL193-BNL2571 | 3.79 | 1.05 | −0.74 | −0.70 | 17.67 |
| Length of Bract | |||||||
| qLOBa1 | 02 | BNL2651-NAU3626 | 3.24 | 0.18 | 0.04 | 0.20 | 8.59 |
| qLOBa2 | 16 | BNL1604-BNL2986 | 3.01 | −0.13 | −0.03 | 0.23 | 7.76 |
| qLOBa3 | 19 | NAU5121-BNL4096 | 4.05 | 0.18 | 0.03 | 0.17 | 12.91 |
| Length of Staminal Column | |||||||
| qLOSa1 | 18 | JESPR153-NAU4105 | 3.78 | 0.52 | 0.11 | 0.20 | 16.30 |
| qLOSa2 | 18 | NAU2488-BNL2571 | 3.76 | 0.52 | 0.11 | 0.20 | 15.84 |
| qLOSa3 | 18 | BNL193-BNL2571 | 3.07 | 0.30 | 0.11 | 0.36 | 14.57 |
| Length of Petal | |||||||
| qLOP1 | 02 | BNL1897-BNL3971 | 3.56 | 0.45 | -0.02 | -0.05 | 19.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rani, S.; Baber, M.; Naqqash, T.; Malik, S.A. Identification and Genetic Mapping of Potential QTLs Conferring Heat Tolerance in Cotton (Gossypium hirsutum L.) by Using Micro Satellite Marker’s Approach. Agronomy 2022, 12, 1381. https://doi.org/10.3390/agronomy12061381
Rani S, Baber M, Naqqash T, Malik SA. Identification and Genetic Mapping of Potential QTLs Conferring Heat Tolerance in Cotton (Gossypium hirsutum L.) by Using Micro Satellite Marker’s Approach. Agronomy. 2022; 12(6):1381. https://doi.org/10.3390/agronomy12061381
Chicago/Turabian StyleRani, Shazia, Muhammad Baber, Tahir Naqqash, and Saeed Ahmad Malik. 2022. "Identification and Genetic Mapping of Potential QTLs Conferring Heat Tolerance in Cotton (Gossypium hirsutum L.) by Using Micro Satellite Marker’s Approach" Agronomy 12, no. 6: 1381. https://doi.org/10.3390/agronomy12061381