
A Time to Sow, a Time to Reap: Modifications to Biological and Economic Rhythms in Southwest Asian Plant and Animal Domestication

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
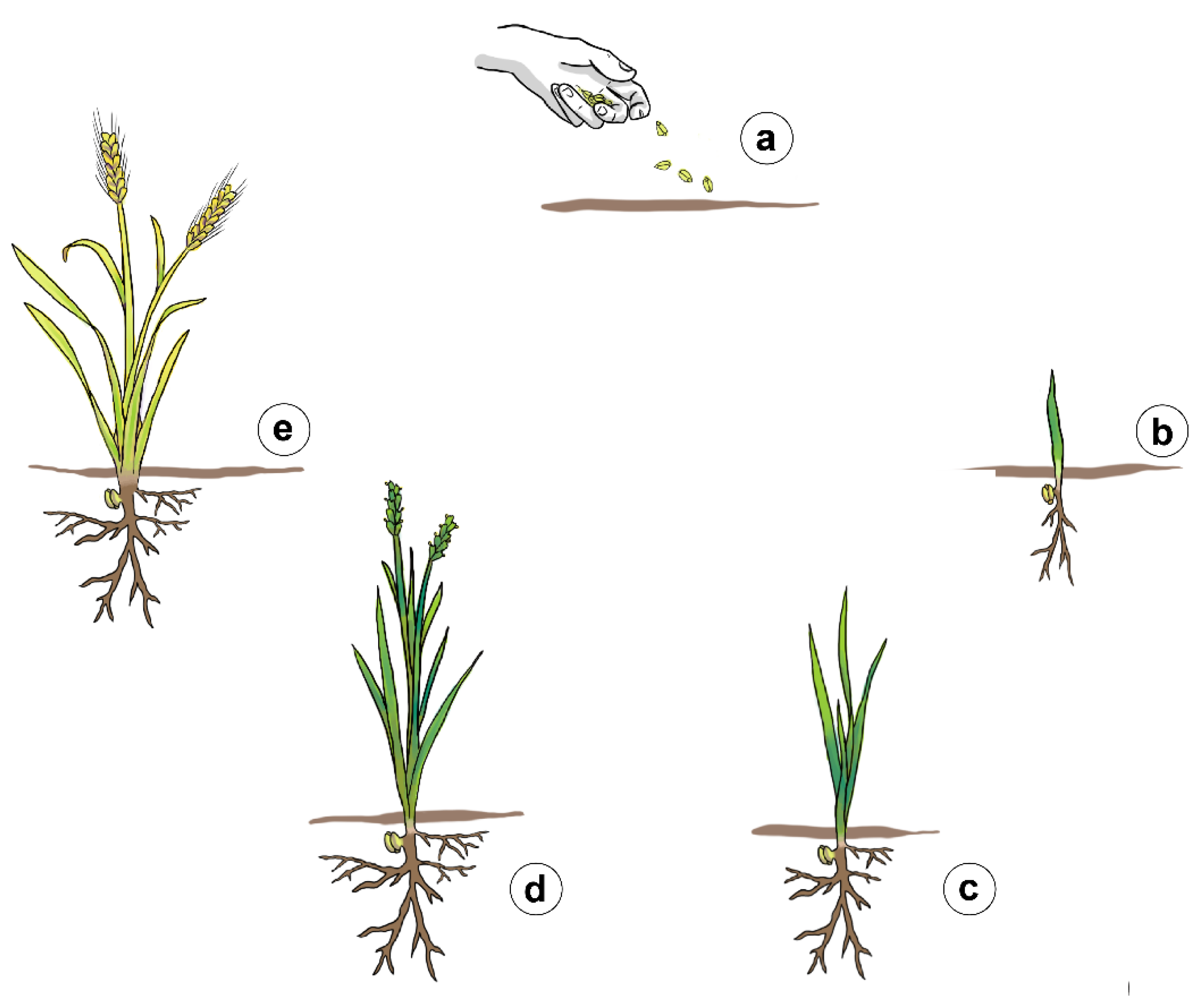
2. Cereals
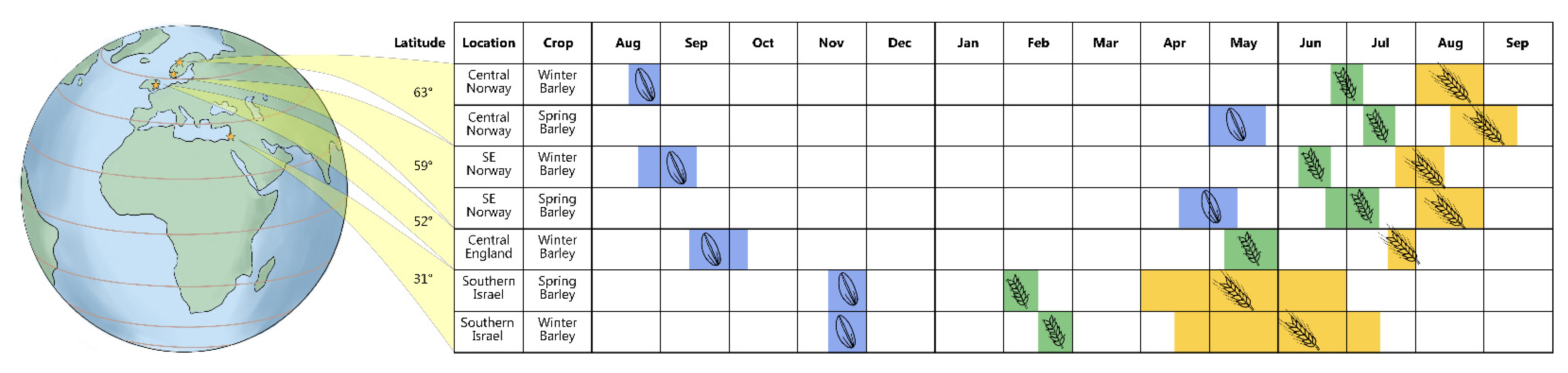
2.1. Dispersal
2.2. Germination
2.3. Vegetative Growth
2.4. Flowering
2.5. Maturation
3. Livestock
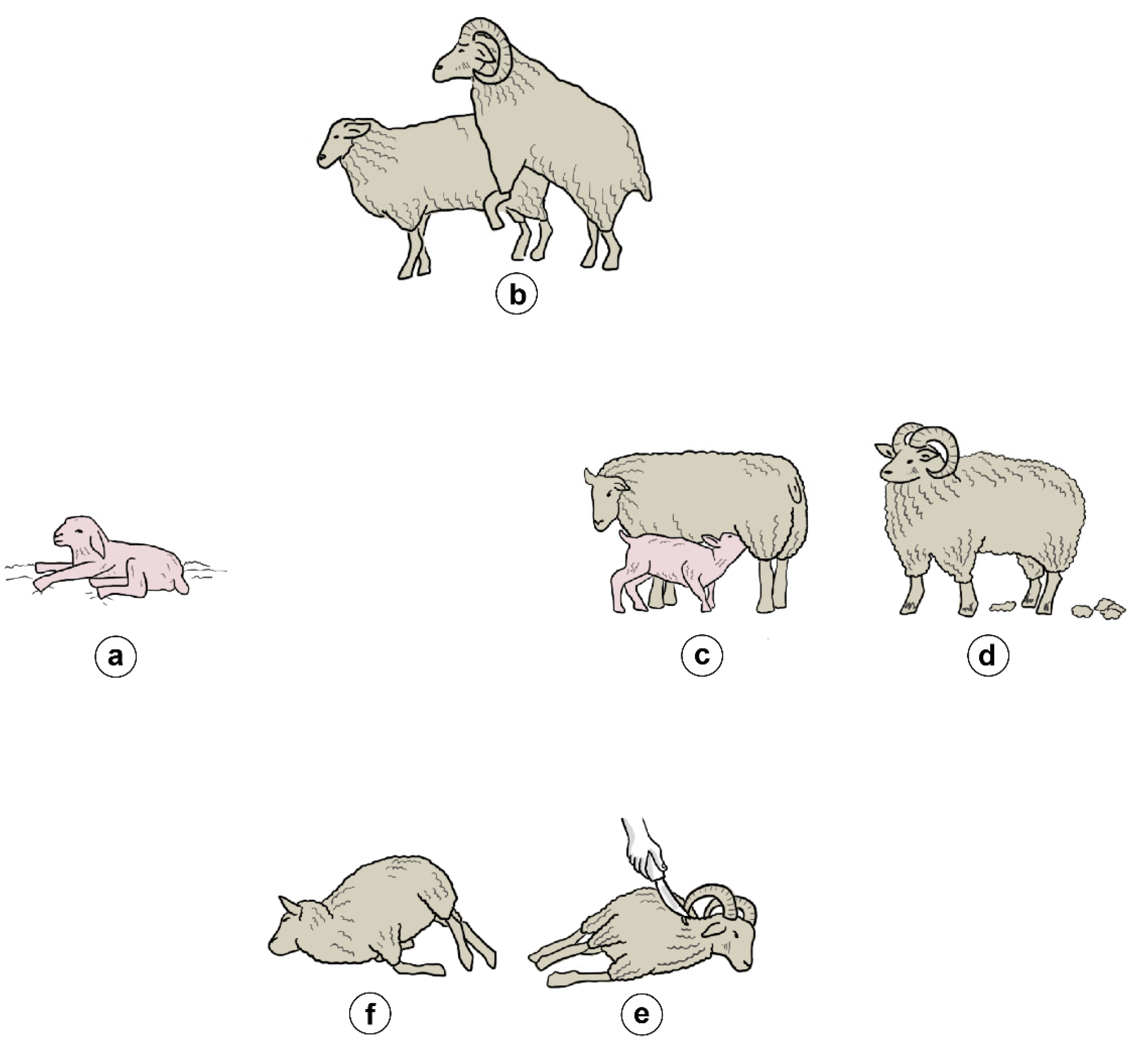
3.1. Reproduction and Birth
3.2. Lactation
3.3. Wool Growth
3.4. Death
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smits, S.A.; Leach, J.; Sonnenburg, E.D.; Gonzalez, C.G.; Lichtman, J.S.; Reid, G.; Knight, R.; Manjurano, A.; Changalucha, J.; Elias, J.E.; et al. Seasonal cycling in the gut microbiome of the Hadza hunter-gatherers of Tanzania. Science 2017, 357, 802–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuks, D.; Marom, N. Sheep and wheat domestication in southwest Asia: A meta-trajectory of intensification and loss. Anim. Front. 2021, 11, 20–29. [Google Scholar] [CrossRef]
- Hillman, G.C.; Davies, M.S. Measured domestication rates in wild wheats and barley under primitive cultivation, and their archaeological implications. J. World Prehistory 1990, 4, 157–222. [Google Scholar] [CrossRef]
- Harlan, J.R. Origin and processes of domestication. In Grass Evolution and Domestication; Chapman, G.P., Ed.; Cambridge University Press: Cambridge, UK, 1992; pp. 150–175. [Google Scholar]
- Hammer, K. Das Domestikationssyndrom. Die Kult. 1984, 32, 11–34. [Google Scholar] [CrossRef]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and processes in crop domestication: An historical review and quantitative analysis of 203 global food crops. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef]
- Brown, T.A. Is the domestication bottleneck a myth? Nat. Plants 2019, 5, 337–338. [Google Scholar] [CrossRef]
- Camacho Villa, T.C.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Resour. 2005, 3, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Held, D.; McGrew, A.; Goldblatt, D.; Perraton, J. Global Transformations: Politics, Economics and Culture; Stanford University Press: Stanford, CA, USA, 1999. [Google Scholar]
- Kislev, M.E. Early agriculture and palaeoecology of Netiv Hagdud. In An Early Neolithic Village in the Jordan Valley; Bar-Yosef, O., Gopher, A., Eds.; Peabody Museum of Archaeology and Ethnology: Cambridge, MA, USA, 1997; pp. 209–236. [Google Scholar]
- Tanno, K.-I.; Willcox, G. How fast was wild wheat domesticated? Science 2006, 311, 1886. [Google Scholar] [CrossRef] [Green Version]
- Weiss, E.; Kislev, M.E.; Hartmann, A. Autonomous cultivation before domestication. Science 2006, 312, 1608–1610. [Google Scholar] [CrossRef] [Green Version]
- Fuller, D.Q. Contrasting patterns in crop domestication and domestication rates: Recent archaeobotanical insights from the Old World. Ann. Bot. 2007, 100, 903–924. [Google Scholar] [CrossRef] [PubMed]
- Purugganan, M.D.; Fuller, D.Q. Archaeological data reveal slow rates of evolution during plant domestication. Evolution 2011, 54, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Riehl, S.; Zeidi, M.; Conard, N.J. Emergence of agriculture in the foothills of the Zagros mountains of Iran. Science 2013, 341, 65–67. [Google Scholar] [CrossRef]
- Fuller, D.Q.; Lucas, L.; Carretero, L.G.; Stevens, C. From intermediate economies to agriculture: Trends in wild food use, domestication and cultivation among early villages in Southwest Asia. Paléorient 2018, 44, 59–74. [Google Scholar]
- Bogaard, A.; Allaby, R.; Arbuckle, B.S.; Bendrey, R.; Crowley, S.; Cucchi, T.; Denham, T.; Frantz, L.; Fuller, D.; Gilbert, T.; et al. Reconsidering domestication from a process archaeology perspective. World Archaeol. 2021, 53, 56–77. [Google Scholar] [CrossRef]
- Abbo, S.; Peleg, Z.; Lev-Yadun, S.; Gopher, A. Does the proportion of shattering vs. non-shattering cereal remains in archeobotanical assemblages reflect Near Eastern Neolithic arable fields? Rev. Palaeobot. Palynol. 2021, 284, 104339. [Google Scholar] [CrossRef]
- Abbo, S.; Lev-Yadun, S.; Gopher, A. Harvest techniques: Hand-pulling and its potential impact on the archaeobotanical record vis a vis Near Eastern plant domestication. Agronomy 2021, 11, 1215. [Google Scholar] [CrossRef]
- Kabukcu, C.; Asouti, E.; Pöllath, N.; Peters, J.; Karul, N. Pathways to plant domestication in Southeast Anatolia based on new data from aceramic Neolithic Gusir Höyük. Sci. Rep. 2021, 11, 2112. [Google Scholar] [CrossRef]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Pourkheirandish, M.; Hensel, G.; Kilian, B.; Senthil, N.; Chen, G.; Sameri, M.; Azhaguvel, P.; Sakuma, S.; Dhanagond, S.; Sharma, R.; et al. Evolution of the grain dispersal system in barley. Cell 2015, 162, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Pourkheirandish, M.; Dai, F.; Sakuma, S.; Kanamori, H.; Distelfeld, A.; Willcox, G.; Kawahara, T.; Matsumoto, T.; Kilian, B.; Komatsuda, T. On the origin of the non-brittle rachis trait of domesticated einkorn wheat. Front. Plant Sci. 2018, 8, 2031. [Google Scholar] [CrossRef] [Green Version]
- Avni, R.; Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Hale, I.; Mascher, M.; Spannagl, M.; Wiebe, K.; et al. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 2017, 357, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Civáň, P.; Brown, T.A. A novel mutation conferring the nonbrittle phenotype of cultivated barley. New Phytol. 2017, 214, 468–472. [Google Scholar] [CrossRef] [Green Version]
- Kislev, M.E.; Weiss, E.; Hartmann, A. Impetus for sowing and the beginning of agriculture: Ground collecting of wild cereals. Proc. Natl. Acad. Sci. USA 2004, 101, 2692–2695. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, A.; Kislev, M.E.; Weiss, E. How and when was wild wheat domesticated? Science 2006, 313, 296–297. [Google Scholar] [CrossRef]
- Childe, V.G. Man Makes Himself; Watts & Company: London, UK, 1936. [Google Scholar]
- Fuller, D.Q.; Allaby, R. Seed dispersal and crop domestication: Shattering, germination and seasonality in evolution under cultivation. In Annual Plant Reviews Volume 38: Fruit Development and Seed Dispersal; Østergaard, L., Ed.; Wiley-Blackwell: Oxford, UK, 2009. [Google Scholar] [CrossRef]
- Abbo, S.; Pinhasi van-Oss, R.; Gopher, A.; Saranga, Y.; Ofner, I.; Peleg, Z. Plant domestication versus crop evolution: A conceptual framework for cereals and grain legumes. Trends Plant Sci. 2014, 19, 351–360. [Google Scholar] [CrossRef]
- Horovitz, A.; Ezrati, S.; Anikster, Y. Are soil seed banks relevant for agriculture in our day? Crop Wild Relat. 2013, 9, 27–30. Available online: https://pgrsecure.bham.ac.uk/sites/default/files/documents/newsletters/CWR_Issue_9.pdf (accessed on 20 March 2022).
- Gao, F.; Ayele, B.T. Functional genomics of seed dormancy in wheat: Advances and prospects. Front. Plant Sci. 2014, 5, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nave, M.; Avni, R.; Ben-Zvi, B.; Hale, I.; Distelfeld, A. QTLs for uniform grain dimensions and germination selected during wheat domestication are co-located on chromosome 4B. Theor. Appl. Genet. 2016, 129, 1303–1315. [Google Scholar] [CrossRef]
- Sato, K.; Yamane, M.; Yamaji, N.; Kanamori, H.; Tagiri, A.; Schwerdt, J.G.; Fincher, G.B.; Matsumoto, T.; Takeda, K.; Komatsuda, T. Alanine aminotransferase controls seed dormancy in barley. Nat. Commun. 2016, 7, 11625. [Google Scholar] [CrossRef]
- Rodríguez, M.V.; Barrero, J.M.; Corbineau, F.; Gubler, F.; Benech-Arnold, R.L. Dormancy in cereals (not too much, not so little): About the mechanisms behind this trait. Seed Sci. Res. 2015, 25, 99–119. [Google Scholar] [CrossRef] [Green Version]
- Kishchenko, O.; Zhou, Y.; Jatayev, S.; Shavrukov, Y.; Borisjuk, N. Gene editing applications to modulate crop flowering time and seed dormancy. aBIOTECH 2020, 1, 233–245. [Google Scholar] [CrossRef]
- Richards, R.A.; Rebetzke, G.J.; Condon, A.G.; Van Herwaarden, A.F. Breeding opportunities for increasing the efficiency of water use and crop yield in temperate cereals. Crop Sci. 2002, 42, 111–121. [Google Scholar] [CrossRef]
- Giles, B.E. The effects of variation in seed size on growth and reproduction in the wild barley Hordeum vulgare ssp. spontaneum. Heredity 1990, 64, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Volis, S.; Mendlinger, S.; Ward, D. Adaptive traits of wild barley plants from Mediterranean and desert origin. Oecologia 2002, 133, 131–138. [Google Scholar] [CrossRef]
- Verhoeven, K.; Biere, A.; Nevo, E.; Van Damme, J.M.M. Can a genetic correlation with seed mass constrain adaptive evolution of seedling desiccation tolerance in wild barley? Int. J. Plant Sci. 2004, 165, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Baillot, N.; Girousse, C.; Allard, V.; Piquet-Pissaloux, A.; Le Gouis, J. Different grainfilling rates explain grain-weight differences along the wheat ear. PLoS ONE 2018, 13, e0209597. [Google Scholar] [CrossRef]
- Fuller, D.Q.; Denham, T.; Arroyo-Kalin, M.; Lucas, L.; Stevens, C.J.; Qin, L.; Allaby, R.G.; Purugganan, M.D. Convergent evolution and parallelism in plant domestication revealed by an expanding archaeological record. Proc. Natl. Acad. Sci. USA 2014, 111, 6147–6152. [Google Scholar] [CrossRef] [Green Version]
- Motuzaite-Matuzeviciute, G.; Mir-Makhamad, B.; Spengler, R.N. Interpreting diachronic size variation in prehistoric Central Asian cereal grains. Front. Ecol. Evol. 2021, 9, 133. [Google Scholar] [CrossRef]
- Watt, M.; Schneebeli, K.; Dong, P.; Wilson, I.W. The shoot and root growth of Brachypodium and its potential as a model for wheat and other cereal crops. Funct. Plant Biol. 2009, 36, 960–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slafer, G.A.; Kantolic, A.G.; Appendino, M.L.; Tranquilli, G.; Miralles, D.J.; Savin, R. Chapter 12 - Genetic and environmental effects on crop development determining adaptation and yield. In Crop Physiology: Application for Genetic Improvement and Agronomy, 2nd ed.; Sadras, V., Calderini, D., Eds.; Academic Press; Elsevier: Amsterdam, The Netherlands, 2015; pp. 285–319. [Google Scholar] [CrossRef]
- Harlan, J.R.; De Wet, J.M.J.; Price, E.G. Comparative evolution of cereals. Evolution 1973, 27, 311–325. [Google Scholar] [CrossRef]
- Evans, L.T. Physiological adaptation to performance as crop plants. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1976, 275, 71–83. [Google Scholar] [CrossRef]
- Zhong, X.; Mei, X.; Li, Y.; Yoshida, H.; Zhao, P.; Wang, X.; Han, L.; Hu, X.; Huang, S.; Huang, J.; et al. Changes in frost resistance of wheat young ears with development during jointing stage. J. Agron. Crop Sci. 2008, 194, 343–349. [Google Scholar] [CrossRef]
- Frederiks, T.M.; Christopher, J.T.; Sutherland, M.W.; Borrell, A.K. Post-head-emergence frost in wheat and barley: Defining the problem, assessing the damage, and identifying resistance. J. Exp. Bot. 2015, 66, 3487–3498. [Google Scholar] [CrossRef] [Green Version]
- Fuller, M.P.; Fuller, A.M.; Kaniouras, S.; Christophers, J.; Fredericks, T. The freezing characteristics of wheat at ear emergence. Eur. J. Agron. 2007, 26, 435–441. [Google Scholar] [CrossRef]
- Mason, H.; Navabi, A.; Frick, B.; O’Donovan, J.; Spaner, D. Cultivar and seeding rate effects on the competitive ability of spring cereals grown under organic production in northern Canada. Agron. J. 2007, 99, 1199–1207. [Google Scholar] [CrossRef]
- Mason, H.E.; Spaner, D. Competitive ability of wheat in conventional and organic management systems: A review of the literature. Can. J. Plant Sci. 2006, 86, 333–343. [Google Scholar] [CrossRef]
- Hedden, P. The genes of the Green Revolution. Trends Genet. 2003, 19, 5–9. [Google Scholar] [CrossRef]
- Borojevic, K.; Borojevic, K. The transfer and history of “reduced height genes” (Rht) in wheat from Japan to Europe. J. Hered. 2005, 96, 455–459. [Google Scholar] [CrossRef] [Green Version]
- Kim, M. Wheat in ancient Korea: A size comparison of carbonized kernels. J. Archaeol. Sci. 2013, 40, 517–525. [Google Scholar] [CrossRef]
- Evans, L.T. Feeding the Ten Billion: Plants and Population Growth; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Jones, G.; Bogaard, A.; Halstead, P.; Charles, M.; Smith, H. Identifying the intensity of crop husbandry practices on the basis of weed floras. Annu. Br. Sch. Athens 1999, 94, 167–189. [Google Scholar] [CrossRef]
- Jones, G.; Bogaard, A.; Charles, M.; Hodgson, J. Distinguishing the effects of agricultural practices relating to fertility and disturbance: A functional ecological approach in archaeobotany. J. Archaeol. Sci. 2000, 27, 1073–1084. [Google Scholar] [CrossRef]
- Fischer, R.A. Growth and water limitation to dryland wheat yield in Australia: A physiological framework. J. Aust. Inst. Agric. 1979, 45, 83–94. [Google Scholar]
- Richards, R.A.; Hunt, J.R.; Kirkegaard, J.; Passioura, J. Yield improvement and adaptation of wheat to water-limited environments in Australia—A case study. Crop Pasture Sci. 2014, 65, 676–689. [Google Scholar] [CrossRef]
- Distelfeld, A.; Li, C.; Dubcovsky, J. Regulation of flowering in temperate cereals. Curr. Opin. Plant Biol. 2009, 12, 178–184. [Google Scholar] [CrossRef] [Green Version]
- Ochagavía, H.; Prieto, P.; Zikhali, M.; Griffiths, S.; Slafer, G.A. Earliness per se by temperature interaction on wheat development. Sci. Rep. 2019, 9, 2584. [Google Scholar] [CrossRef] [Green Version]
- Lister, D.L.; Jones, H.; Oliveira, H.R.; Petrie, C.A.; Liu, X.; Cockram, J.; Kneale, C.J.; Kovaleva, O.; Jones, M.K. Barley heads east: Genetic analyses reveal routes of spread through diverse Eurasian landscapes. PLoS ONE 2018, 13, e0196652. [Google Scholar] [CrossRef]
- Liu, X.; Jones, P.J.; Motuzaite-Matuzeviciute, G.M.; Hunt, H.V.; Lister, D.L.; An, T.; Przelomska, N.; Kneale, C.J.; Zhao, Z.; Jones, M.K. From ecological opportunism to multi-cropping: Mapping food globalisation in prehistory. Quat. Sci. Rev. 2019, 206, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Saisho, D.; Ishii, M.; Hori, K.; Sato, K. Natural variation of barley vernalization requirements: Implication of quantitative variation of winter growth habit as an adaptive trait in East Asia. Plant Cell Physiol. 2011, 52, 775–784. [Google Scholar] [CrossRef]
- Jones, G.; Jones, H.; Charles, M.P.; Jones, M.K.; Colledge, S.; Leigh, F.J.; Lister, D.A.; Smith, L.M.; Powell, W.; Brown, T.A. Phylogeographic analysis of barley DNA as evidence for the spread of Neolithic agriculture through Europe. J. Archaeol. Sci. 2012, 39, 3230–3238. [Google Scholar] [CrossRef]
- Turner, A.; Beales, J.; Faure, S.; Dunford, R.P.; Laurie, D.A. The pseudo-response regulator Ppd-H1 provides adaptation to photoperiod in barley. Science 2005, 310, 1031–1034. [Google Scholar] [CrossRef]
- Jones, H.; Leigh, F.J.; Mackay, I.; Bower, M.A.; Smith, L.M.; Charles, M.P.; Jones, G.; Jones, M.K.; Brown, T.A.; Powell, W. Population-based resequencing reveals that the flowering time adaptation of cultivated barley originated east of the Fertile Crescent. Mol. Biol. Evol. 2008, 25, 2211–2219. [Google Scholar] [CrossRef] [Green Version]
- Lister, D.L.; Thaw, S.; Bower, M.A.; Jones, H.; Charles, M.P.; Jones, G.; Smith, L.M.; Howe, C.J.; Brown, T.A.; Jones, M.K. Latitudinal variation in a photoperiod response gene in European barley: Insight into the dynamics of agricultural spread from ‘historic’ specimens. J. Archaeol. Sci. 2009, 36, 1092–1098. [Google Scholar] [CrossRef]
- Hyles, J.; Maxwell, T.; Bloomfield, M.T.; Hunt, J.R.; Trethowan, R.M.; Trevaskis, B. Phenology and related traits for wheat adaptation. Heredity 2020, 125, 417–430. [Google Scholar] [CrossRef]
- Miralles, D.J.; Richards, R.A.; Slafer, G.A. Duration of the stem elongation period influences the number of fertile florets in wheat and barley. Funct. Plant Biol. 2000, 27, 931–940. [Google Scholar] [CrossRef]
- Borràs-Gelonch, G.; Rebetzke, G.J.; Richards, R.A.; Romagosa, I. Genetic control of duration of pre-anthesis phases in wheat (Triticum aestivum L.) and relationships to leaf appearance, tillering, and dry matter accumulation. J. Exp. Bot. 2012, 63, 69–89. [Google Scholar] [CrossRef] [Green Version]
- Riley, T.J.; Freimuth, G. Field systems and frost drainage in the prehistoric agriculture of the upper Great Lakes. Am. Antiq. 1979, 44, 271–285. [Google Scholar] [CrossRef]
- Celestina, C.; Bloomfield, M.T.; Stefanova, K.; Hunt, J.R. Use of spike moisture content to define physiological maturity and quantify progress through grain development in wheat and barley. Crop Pasture Sci. 2021, 72, 95–104. [Google Scholar] [CrossRef]
- Falk, D. Pre-domesticating wild relatives as new sources of novel genetic diversity. In Enhancing Crop Genepool Use: Capturing Wild Relative and Landrace Diversity for Crop Improvement; Maxted, N., Dulloo, M.E., Ford-Lloyd, B.V., Eds.; CABI: Wallingford, UK; Boston, MA, USA, 2016; pp. 10–19. [Google Scholar]
- Uauy, C.; Distelfeld, A.; Fahima, T.; Blechl, A.; Dubcovsky, J. A NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 2006, 314, 1298–1301. [Google Scholar] [CrossRef] [Green Version]
- Uauy, C.; Brevis, J.C.; Dubcovsky, J. The high grain protein content gene Gpc-B1 accelerates senescence and has pleiotropic effects on protein content in wheat. J. Exp. Bot. 2006, 57, 2785–2794. [Google Scholar] [CrossRef] [PubMed]
- Distelfeld, A.; Cakmak, I.; Peleg, Z.; Ozturk, L.; Yazici, A.M.; Budak, H.; Saranga, Y.; Fahima, T. Multiple QTL-effects of wheat Gpc-B1 locus on grain protein and micronutrient concentrations. Physiol. Plant. 2007, 129, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Asplund, L.; Hagenblad, J.; Leino, M.W. Re-evaluating the history of the wheat domestication gene NAM-B1 using historical plant material. J. Archaeol. Sci. 2010, 37, 2303–2307. [Google Scholar] [CrossRef]
- Johnson, D.L. The Nature of Nomadism: A Comparative Study of Pastoral Migrations in Southwestern Asia and Northern Africa; Department of Geography, University of Chicago: Chicago, IL, USA, 1969. [Google Scholar]
- Khazanov, A.M. Nomads and the Outside World; Cambridge University Press: Cambridge, UK, 1984. [Google Scholar]
- Marx, E. Bedouin of the Negev; Manchester University Press: Manchester, UK, 1967. [Google Scholar]
- Zuckerman, S. The breeding seasons of mammals in captivity. Proc. Zool. Soc. Lond. 1952, 122, 827–950. [Google Scholar] [CrossRef]
- Hafez, E.S.E. Studies on the breeding season and reproduction of the ewe. J. Agric. Sci. 1952, 42, 189–231. [Google Scholar] [CrossRef]
- Setchell, B. Domestication and reproduction. Anim. Reprod. Sci. 1992, 28, 195–202. [Google Scholar] [CrossRef]
- Rosa, H.J.D.; Bryant, M. Seasonality of reproduction in sheep. Small Rumin. Res. 2003, 48, 155–171. [Google Scholar] [CrossRef]
- Delgadillo, J.A. Environmental and social cues can be used in combination to develop sustainable breeding techniques for goat reproduction in the subtropics. Animal 2011, 5, 74–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karsch, F.J.; Bittman, E.L.; Foster, D.L.; Goodman, R.L.; Legan, S.J.; Robinson, J.E. Neuroendocrine basis of seasonal reproduction. Recent Prog. Horm. Res. 1984, 40, 185–232. [Google Scholar] [CrossRef]
- Berteaux, D.; Micol, T. Population studies and reproduction of the feral cattle (Bos taurus) of Amsterdam Island, Indian Ocean. J. Zool. 1992, 228, 265–276. [Google Scholar] [CrossRef]
- Reinhardt, C.; Reinhardt, A.; Reinhardt, V. Social behaviour and reproductive performance in semi-wild Scottish Highland cattle. Appl. Anim. Behav. Sci. 1986, 15, 125–136. [Google Scholar] [CrossRef]
- Balasse, M.; Gillis, R.; Živaljević, I.; Berthon, R.; Kovačiková, L.; Fiorillo, D.; Arbogast, R.M.; Bălăşescu, A.; Bréhard, S.; Nyerges, É.Á.; et al. Seasonal calving in European Prehistoric cattle and its impacts on milk availability and cheese-making. Sci. Rep. 2021, 11, 8185. [Google Scholar] [CrossRef]
- Hall, S.J.G.; Hall, J.G. Inbreeding and population dynamics of the Chillingham cattle (Bos taurus). J. Zool. 1988, 216, 479–493. [Google Scholar] [CrossRef]
- Orihuela, A. Some factors affecting the behavioural manifestation of oestrus in cattle: A review. Appl. Anim. Behav. Sci. 2000, 70, 1–16. [Google Scholar] [CrossRef]
- Morrison, M. Evidence for herdsmen and animal husbandry in the Nuzi documents. In Studies on the Civilization and Culture of Nuzi and the Hurrians in Honor of Ernest R. Lacheman; Owens, D., Morrison, M., Eds.; Eisenbrauns: Winona Lake, IN, USA, 1981; pp. 257–296. [Google Scholar]
- Barth, F. Nomads of South Persia: The Basseri tribe of the Khamseh Confederacy; Universitetsforlaget: Oslo, Norway, 1965. [Google Scholar]
- Hadjikoumis, A.; Vigne, J.-D.; Simmons, A.; Guilaine, J.; Fiorillo, D.; Balasse, M. Autumn/winter births in traditional and Pre-Pottery Neolithic caprine husbandry in Cyprus: Evidence from ethnography and stable isotopes. J. Anthr. Archaeol. 2018, 53, 102–111. [Google Scholar] [CrossRef]
- Abu-Rabia, A. The Negev Bedouin and Livestock Rearing: Social, Economic and Political Aspects; Berg: Oxford, UK, 1994. [Google Scholar]
- Ginguld, M.; Perevolotsky, A.; Ungar, E.D. Living on the margins: Livelihood strategies of Bedouin herd-owners in the northern Negev, Israel. Hum. Ecol. 1997, 25, 567–591. [Google Scholar] [CrossRef]
- Perevolotsky, A.; Perevolotsky, A. Subsistence Patterns of the Jebaliyah Bedouin in the High Mountain Region of Southern Sinai; SPNI: Tel Aviv, Israel, 1979; (In Hebrew with an English Summary). [Google Scholar]
- Perevolotsky, A.; Perevolotsky, A.; Noy-Meir, I. Environmental adaptation and Economic change in a pastoral mountain society: The case of the Jabaliyah Bedouin of the Mt. Sinai Region. Mt. Res. Dev. 1989, 9, 153. [Google Scholar] [CrossRef]
- Tapper, R. Pasture and Politics: Economics, Conflict and Ritual among Shahsevan Nomads of Northwestern Iran; Academic Press: London, UK, 1979. [Google Scholar]
- Black-Michaud, J. Sheep and Land: The Economics of Power in a Tribal Society; Cambridge University Press: Cambridge, UK, 1986. [Google Scholar]
- Lancaster, W.; Lancaster, F. Limitations on sheep and goat herding in the eastern Badia of Jordan: An ethno-archaeological enquiry. Levant 1991, 23, 125–138. [Google Scholar] [CrossRef]
- Widell, M. Destined for Slaughter: Identifying Seasonal Breeding Patterns in Sheep and Goats in Early Babylonia. J. Near East. Stud. 2020, 79, 209–223. [Google Scholar] [CrossRef]
- Tornero, C.; Balasse, M.; Molist, M.; Saña, M. Seasonal reproductive patterns of early domestic sheep at Tell Halula (PPNB, Middle Euphrates Valley): Evidence from sequential oxygen isotope analyses of tooth enamel. J. Archaeol. Sci. Rep. 2016, 6, 810–818. [Google Scholar] [CrossRef]
- Dahl, G.; Hjort, A. Having Herds: Pastoral Herd Growth and Household Economy; Department of Social Anthropology, University of Stockholm: Stockholm, Sweden, 1976. [Google Scholar]
- Postgate, J.; Payne, S. Some Old Babylonian shepherds and their flocks. J. Semit. Stud. 1975, 20, 1–21. [Google Scholar] [CrossRef]
- Henton, E. The combined use of oxygen isotopes and microwear in sheep teeth to elucidate seasonal management of domestic herds: The case study of Çatalhöyük, central Anatolia. J. Archaeol. Sci. 2012, 39, 3264–3276. [Google Scholar] [CrossRef]
- Tornero, C.; Balasse, M.; Bréhard, S.; Carrère, I.; Fiorillo, D.; Guilaine, J.; Vigne, J.-D.; Manen, C. Early evidence of sheep lambing de-seasoning in the western Mediterranean in the sixth millennium BCE. Sci. Rep. 2020, 10, 12798. [Google Scholar] [CrossRef]
- Blaise, E.; Balasse, M. Seasonality and season of birth of modern and late Neolithic sheep from south-eastern France using tooth enamel δ18O analysis. J. Archaeol. Sci. 2011, 38, 3085–3093. [Google Scholar] [CrossRef] [Green Version]
- Balasse, M.; Boury, L.; Ughetto-Monfrin, J.; Tresset, A. Stable isotope insights (δ18O, δ13C) into cattle and sheep husbandry at Bercy (Paris, France, 4th millennium BC): Birth seasonality and winter leaf foddering. Environ. Archaeol. 2012, 17, 29–44. [Google Scholar] [CrossRef]
- Balasse, M.; Bălăşescu, A.; Janzen, A.; Ughetto-Monfrin, J.; Mirea, P.; Andreescu, R. Early herding at Măgura-Boldul lui Moş Ivănuş (early sixth millennium BC, Romania): Environments and seasonality from stable isotope analysis. Eur. J. Archaeol. 2013, 16, 221–246. [Google Scholar] [CrossRef]
- Balasse, M.; Tresset, A.; Bălăşescu, A.; Blaise, E.; Tornero, C.; Gandois, H.; Fiorillo, D.; Nyerges, É.Á.; Frémondeau, D.; Banffy, E.; et al. Animal Board Invited Review: Sheep birth distribution in past herds: A review for prehistoric Europe (6th to 3rd millennia BC). Animal 2017, 11, 2229–2236. [Google Scholar] [CrossRef] [Green Version]
- Balasse, M.; Renault-Fabregon, L.; Gandois, H.; Fiorillo, D.; Gorczyk, J.; Bacvarov, K.; Ivanova, M. Neolithic sheep birth distribution: Results from Nova Nadezhda (sixth millennium BC, Bulgaria) and a reassessment of European data with a new modern reference set including upper and lower molars. J. Archaeol. Sci. 2020, 118, 105139. [Google Scholar] [CrossRef]
- Tornero, C.; Bălăşescu, A.; Ughetto-Monfrin, J.; Voinea, V.; Balasse, M. Seasonality and season of birth in early Eneolithic sheep from Cheia (Romania): Methodological advances and implications for animal economy. J. Archaeol. Sci. 2013, 40, 4039–4055. [Google Scholar] [CrossRef]
- Chazin, H. Multi-season reproduction and pastoralist production strategies: New approaches to birth seasonality from the South Caucasus region. J. Field Archaeol. 2021, 46, 448–460. [Google Scholar] [CrossRef]
- Weinberg, P. On the status and biology of the wild goat in Daghestan (Russia). J. Mt. Ecol. 2001, 6, 31–40. [Google Scholar]
- Langbein, J.; Streich, J.; Scheibe, K.M. Characteristic activity patterns of female mouflons (Ovis orientalis musimon) in the lambing period. Appl. Anim. Behav. Sci. 1998, 58, 281–292. [Google Scholar] [CrossRef]
- Ciuti, S.; Pipia, A.; Grignolio, S.; Ghiandai, F.; Apollonio, M. Space use, habitat selection and activity patterns of female Sardinian mouflon (Ovis orientalis musimon) during the lambing season. Eur. J. Wildl. Res. 2009, 55, 589–595. [Google Scholar] [CrossRef]
- Chemineau, P.; Guillaume, D.; Migaud, M.; Thiéry, J.C.; Pellicer-Rubio, M.T.; Malpaux, B. Seasonality of reproduction in mammals: Intimate regulatory mechanisms and practical implications. Reprod. Domest. Anim. 2008, 43, 40–47. [Google Scholar] [CrossRef]
- Posbergh, C.J.; Thonney, M.; Huson, H.J. Genomic approaches identify novel gene associations with out of season lambing in sheep. J. Hered. 2019, 110, 577–586. [Google Scholar] [CrossRef]
- Towers, J.; Jay, M.; Mainland, I.; Nehlich, O.; Montgomery, J. A calf for all seasons? The potential of stable isotope analysis to investigate prehistoric husbandry practices. J. Archaeol. Sci. 2011, 38, 1858–1868. [Google Scholar] [CrossRef] [Green Version]
- Gron, K.J.; Montgomery, J.; Rowley-Conwy, P. Cattle management for dairying in Scandinavia’s earliest Neolithic. PLoS ONE 2015, 10, e0131267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Towers, J.; Mainland, I.; Montgomery, J.; Bond, J. Calving seasonality at Pool, Orkney during the first millennium AD: An investigation using intra-tooth isotope ratio analysis of cattle molar enamel. Environ. Archaeol. 2017, 22, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Gillis, R.; Kendall, I.; Roffet-Salque, M.; Zanon, M.; Anders, A.; Arbogast, R.M.; Bogucki, P.; Brychova, V.; Casanova, E.; Claßen, E.; et al. Forest Ecosystems and Evolution of Cattle Husbandry Practices of the Earliest Central European Farming Societies. Res. Sq. Prepr. 2022, 1–26, PREPRINT (Version 1). [Google Scholar] [CrossRef]
- deNicolo, G. Accelerated and Out-of-Season Lamb Production in New Zealand. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 2007. [Google Scholar]
- Stępień, M. Animal Husbandry in the Ancient Near East: A Prosopographic Study of Third Millennium Umma; CDL Press: Bethesda, MD, USA, 1996. [Google Scholar]
- Evershed, R.P.; Payne, S.; Sherratt, A.G.; Copley, M.S.; Coolidge, J.; Urem-Kotsu, D.; Kotsakis, K.; Özdoğan, M.; Özdoğan, A.E.; Nieuwenhuyse, O.; et al. Earliest date for milk use in the Near East and southeastern Europe linked to cattle herding. Nature 2008, 455, 528–531. [Google Scholar] [CrossRef]
- Debono Spiteri, C.; Gillis, R.E.; Roffet-Salque, M.; Navarro, L.C.; Guilaine, J.; Manen, C.; Muntoni, I.M.; Segui, M.S.; Urem-Kotsou, D.; Whelton, H.L.; et al. Regional asynchronicity in dairy production and processing in early farming communities of the northern Mediterranean. Proc. Natl. Acad. Sci. USA 2016, 113, 13594–13599. [Google Scholar] [CrossRef] [Green Version]
- Dunne, J.; di Lernia, S.; Chłodnicki, M.; Kherbouche, F.; Evershed, R. Timing and pace of dairying inception and animal husbandry practices across Holocene North Africa. Quat. Int. 2018, 471, 147–159. [Google Scholar] [CrossRef]
- Hendy, J.; Colonese, A.C.; Franz, I.; Fernandes, R.; Fischer, R.; Orton, D.; Lucquin, A.; Spindler, L.; Anvari, J.; Stroud, E.; et al. Ancient proteins from ceramic vessels at Çatalhöyük West reveal the hidden cuisine of early farmers. Nat. Commun. 2018, 9, 4064. [Google Scholar] [CrossRef]
- Simoons, F.J. The antiquity of dairying in Asia and Africa. Geogr. Rev. 1971, 61, 431. [Google Scholar] [CrossRef]
- Bleasdale, M.; Richter, K.K.; Janzen, A.; Brown, S.; Scott, A.; Zech, J.; Wilkin, S.; Wang, K.; Schiffels, S.; Desideri, J.; et al. Ancient proteins provide evidence of dairy consumption in eastern Africa. Nat. Commun. 2021, 12, 632. [Google Scholar] [CrossRef]
- Ryder, M.L. Sheep and Man; Duckworth: London, UK, 1983. [Google Scholar]
- Salque, M.; Bogucki, P.I.; Pyzel, J.; Sobkowiak-Tabaka, I.; Grygiel, R.; Szmyt, M.; Evershed, R.P. Earliest evidence for cheese making in the sixth millennium BC in northern Europe. Nature 2013, 493, 522–525. [Google Scholar] [CrossRef] [PubMed]
- Charlton, S.; Ramsøe, A.; Collins, M.; Craig, O.E.; Fischer, R.; Alexander, M.; Speller, C.F. New insights into Neolithic milk consumption through proteomic analysis of dental calculus. Archaeol. Anthr. Sci. 2019, 11, 6183–6196. [Google Scholar] [CrossRef] [Green Version]
- Cannas, A.; Nudda, A.; Pulina, G. Nutritional strategies to improve lactation persistency in dairy ewes. In Proceedings of the 8th Great Lakes Dairy Sheep Symposium, Ithaca, NY, USA, 7–9 November 2002; Cornell University: Ithaca, NY, USA, 2002. [Google Scholar]
- Pulina, G.; Nudda, A.; Macciotta, N.P.P.; Battacone, G.; Rassu, S.P.G.; Cannas, A. Non-nutritional factors affecting lactation persistency in dairy ewes: A review. Ital. J. Anim. Sci. 2007, 6, 115–141. [Google Scholar] [CrossRef] [Green Version]
- Goetsch, A.; Zeng, S.; Gipson, T. Factors affecting goat milk production and quality. Small Rumin. Res. 2011, 101, 55–63. [Google Scholar] [CrossRef]
- Abecia, J.A.; Garcia, A.; Castillo, L.; Palacios, C. The effects of weather on milk production in dairy sheep vary by month of lambing and lactation phase. J. Anim. Behav. Biometeorol. 2017, 5, 56–63. [Google Scholar] [CrossRef]
- Smith, S.E. The Environmental adaptation of nomads in the West African Sahel: A key to understanding prehistoric pastoralists. In The Nomadic Alternative; Barfield, T.J., Ed.; Prentice Hall: Englewood Cliffs, NJ, USA, 1978; pp. 75–96. [Google Scholar]
- Turner, C.W.; Ragsdale, A.C.; Brody, S. How the advance of the period of lactation affects the milk flow. J. Dairy Sci. 1923, 6, 527–531. [Google Scholar] [CrossRef]
- Wood, P.D.P. Algebraic model of the lactation curve in cattle. Nature 1967, 216, 164–165. [Google Scholar] [CrossRef]
- Wood, P.D.P. Algebraic models of the lactation curves for milk, fat and protein production, with estimates of seasonal variation. Anim. Sci. 1976, 22, 35–40. [Google Scholar] [CrossRef]
- Wood, P.D.P. Factors affecting the shape of the lactation curve in cattle. Anim. Sci. 1969, 11, 307–316. [Google Scholar] [CrossRef]
- García, S.; Holmes, C. Lactation curves of autumn- and spring-calved cows in pasture-based dairy systems. Livest. Prod. Sci. 2001, 68, 189–203. [Google Scholar] [CrossRef]
- Auldist, M.J.; O’Brien, G.; Cole, D.; Macmillan, K.L.; Grainger, C. Effects of varying lactation length on milk production capacity of cows in pasture-based dairying systems. J. Dairy Sci. 2007, 90, 3234–3241. [Google Scholar] [CrossRef]
- Carta, A.; Sanna, S.; Casu, S. Estimating lactation curves and seasonal effects for milk, fat and protein in Sarda dairy sheep with a test day model. Livest. Prod. Sci. 1995, 44, 37–44. [Google Scholar] [CrossRef]
- Assan, N. Significance of litter size, duration of dry period and stage of pregnacy on milk yield and composition in dairy animals. Sci. J. Rev. 2014, 3, 993–1003. [Google Scholar]
- Zobel, G.; Weary, D.; Leslie, K.; Von Keyserlingk, M. Invited review: Cessation of lactation: Effects on animal welfare. J. Dairy Sci. 2015, 98, 8263–8277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Quellec, J.-L. Provoking lactation by the insufflation technique as documented by the rock images of the Sahara. Anthropozoologica 2011, 46, 65–125. [Google Scholar] [CrossRef]
- Jordan, D.L.; Erb, R.; Malven, P.; Callahan, C.; Veenhuizen, E. Artificial induction of lactation in cattle: Effect of modified treatments on milk yield, fertility, and hormones in blood plasma and milk. Theriogenology 1981, 16, 315–329. [Google Scholar] [CrossRef]
- Magliaro, A.; Kensinger, R.; Ford, S.; O’Connor, M.; Muller, L.; Graboski, R. Induced lactation in nonpregnant cows: Profitability and response to bovine somatotropin. J. Dairy Sci. 2004, 87, 3290–3297. [Google Scholar] [CrossRef] [Green Version]
- Grummer, R.; Rastani, R. Why reevaluate dry period length? J. Dairy Sci. 2004, 87, E77–E85. [Google Scholar] [CrossRef] [Green Version]
- Belanche, A.; Martín-García, A.I.; Fernández-Álvarez, J.; Pleguezuelos, J.; Mantecón, Á.R.; Yáñez-Ruiz, D.R. Optimizing management of dairy goat farms through individual animal data interpretation: A case study of smart farming in Spain. Agric. Syst. 2019, 173, 27–38. [Google Scholar] [CrossRef]
- Gross, J.; Bruckmaier, R. Invited review: Metabolic challenges and adaptation during different functional stages of the mammary gland in dairy cows: Perspectives for sustainable milk production. J. Dairy Sci. 2019, 102, 2828–2843. [Google Scholar] [CrossRef] [Green Version]
- Kok, A.; Chen, J.; Kemp, B.; van Knegsel, A.T.M. Review: Dry period length in dairy cows and consequences for metabolism and welfare and customised management strategies. Animal 2019, 13, s42–s51. [Google Scholar] [CrossRef] [Green Version]
- Brody, S.; Ragsdale, A.C.; Turner, C.W. The rate of growth of the dairy cow: III. The relation between growth in weight and increase of milk secretion with age. J. Gen. Physiol. 1923, 6, 21–30. [Google Scholar] [CrossRef]
- Brody, S.; Ragsdale, A.C.; Turner, C.W. The rate of growth of the dairy cow: IV. Growth and senescence as measured by the rise and fall of milk secretion with age. J. Gen. Physiol. 1923, 6, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Payne, S. Kill-off Patterns in Sheep and Goats: The mandibles from Aşvan Kale. Anatol. Stud. 1973, 23, 281–303. [Google Scholar] [CrossRef]
- Daly, K.G.; Delser, P.M.; Mullin, V.E.; Scheu, A.; Mattiangeli, V.; Teasdale, M.D.; Hare, A.J.; Burger, J.; Verdugo, M.P.; Collins, M.J.; et al. Ancient goat genomes reveal mosaic domestication in the Fertile Crescent. Science 2018, 361, 85–88. [Google Scholar] [CrossRef] [Green Version]
- Ryder, M.L. A study of the coat of the Mouflon Ovis musimon with special reference to seasonal change. Proc. Zool. Soc. Lon. 1960, 135, 387–408. [Google Scholar] [CrossRef]
- Ciani, E.; Lasagna, E.; D’Andrea, M.; Alloggio, I.; Marroni, F.; Ceccobelli, S.; Bermejo, J.V.D.; Sarti, F.M.; Kijas, J.; Lenstra, J.A.; et al. Merino and Merino-derived sheep breeds: A genome-wide intercontinental study. Genet. Sel. Evol. 2015, 47, 64. [Google Scholar] [CrossRef] [Green Version]
- Ciani, E.; Mastrangelo, S.; Da Silva, A.; Marroni, F.; Ferenčaković, M.; Ajmone-Marsan, P.; Baird, H.; Barbato, M.; Colli, L.; Delvento, C.; et al. On the origin of European sheep as revealed by the diversity of the Balkan breeds and by optimizing population-genetic analysis tools. Genet. Sel. Evol. 2020, 52, 25. [Google Scholar] [CrossRef]
- Gootwine, E. Chapter 10: Genetics and breeding of sheep and goats. In Animal Agriculture; Bazer, F.W., Lamb, G.C., Wu, G., Eds.; Academic Press: London, UK, 2020; pp. 183–198. [Google Scholar] [CrossRef]
- Ryder, M.L. The interaction between biological and technological change during the development of different fleece types in sheep. Anthropozoologica 1992, 16, 131–140. [Google Scholar]
- Gleba, M. From textiles to sheep: Investigating wool fibre development in pre-Roman Italy using scanning electron microscopy (SEM). J. Archaeol. Sci. 2012, 39, 3643–3661. [Google Scholar] [CrossRef]
- Rast-Eicher, A. Bronze and Iron Age wools in Europe. In Wool Economy in the Ancient Near East and the Aegean: From the Beginnings of Sheep Husbandry to Institutional Textile Industry; Breniquet, C., Michel, C., Eds.; Ancient Textiles, Series; Oxbow Books: Oxford, UK; Philadelphia, PA, USA, 2014. [Google Scholar]
- Stiner, M.C.; Buitenhuis, H.; Duru, G.; Kuhn, S.L.; Mentzer, S.M.; Munro, N.D.; Pöllath, N.; Quade, J.; Tsartsidou, G.; Özbaşaran, M. A forager–herder trade-off, from broad-spectrum hunting to sheep management at Aşıklı Höyük, Turkey. Proc. Natl. Acad. Sci. USA 2014, 111, 8404–8409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saña, M.; Tornero, C. Use of animal fibres during the Neolithisation in the Middle Euphrates Valley (Syria): An archaeozoological approach. Paléorient 2012, 38, 79–91. [Google Scholar] [CrossRef]
- Rooijakkers, C.T. Spinning animal fibres at late Neolithic Tell Sabi Abyad, Syria? Paléorient 2012, 38, 93–109. [Google Scholar] [CrossRef]
- Breniquet, C. The archaeology of wool in early Mesopotamia: Sources, methods, perspectives. In Wool Economy in the Ancient Near East and the Aegean: From the Beginnings of Sheep Husbandry to Institutional Textile Industry; Breniquet, C., Michel, C., Eds.; Oxbow Books: Oxford, UK, 2014; pp. 52–78. [Google Scholar]
- Jackson, N.; Maddocks, I.G.; Watts, J.E.; Scobie, D.; Mason, R.S.; Gordon-Thomson, C.; Stockwell, S.; Moore, G.P. Evolution of the sheep coat: The impact of domestication on its structure and development. Genet. Res. 2020, 102, e4. [Google Scholar] [CrossRef]
- Sherratt, A. Plough and pastoralism: Aspects of the secondary products revolution. In Pattern of the Past: Studies in Honour of David Clarke; Hodder, I., Isaac, G., Hammond, N., Eds.; Cambridge University Press: Cambridge, UK, 1981; pp. 261–306. [Google Scholar]
- Sherratt, A. The secondary exploitation of animals in the Old World. World Archaeol. 1983, 15, 90–104. [Google Scholar] [CrossRef]
- Greenfield, H.J. The Secondary Products Revolution: The past, the present and the future. World Archaeol. 2010, 42, 29–54. [Google Scholar] [CrossRef]
- Breniquet, C. Early wool of Mesopotamia, C. 7000–3000 BC: Between prestige and economy. In The Competition of Fibres: Early Textile Production in Western Asia, South-East and Central Europe (10,000-500 BCE); Schier, W., Pollock, S., Eds.; Oxbow Books: Oxford, UK, 2020; pp. 17–26. [Google Scholar]
- Price, M.; Rowan, Y.M.; Kersel, M.M.; Makarewicz, C.A. Fodder, pasture, and the development of complex society in the Chalcolithic: Isotopic perspectives on animal husbandry at Marj Rabba. Archaeol. Anthr. Sci. 2020, 12, 95. [Google Scholar] [CrossRef]
- Fall, P.L.; Lines, L.; Falconer, S.E. Seeds of civilization: Bronze Age rural economy and ecology in the Southern Levant. Ann. Assoc. Am. Geogr. 1998, 88, 107–125. [Google Scholar] [CrossRef]
- Marston, J.M.; Çakırlar, C.; Luke, C.; Kováčik, P.; Slim, F.G.; Shin, N.; Roosevelt, C.H. Agropastoral economies and land use in Bronze Age western Anatolia. Environ. Archaeol. 2021, 1–15. [Google Scholar] [CrossRef]
- Hilzheimer, M. Sheep. Antiquity 1936, 10, 195–206. [Google Scholar] [CrossRef]
- Ryder, M. L Sheep–Hilzheimer 45 years on. Antiquity 1982, 46, 15–23. [Google Scholar] [CrossRef]
- Algaze, G. Ancient Mesopotamia at the Dawn of Civilization: The Evolution of an Urban Landscape; Chicago University Press: Chicago, IL, USA, 2008. [Google Scholar]
- Waetzoldt, H. Unterzuchungen zur Neusumerischen Textilindustrie; Studi Economici e Tecnologici I: Rome, Italy, 1972. [Google Scholar]
- Steinkeller, P. Sheep and goat terminology in Ur III sources from Drehem. In Domestic Animals of Mesopotamia; Postgate, J., Powell, M., Eds.; University of Cambridge: Cambridge, UK, 1995; Volume 7, pp. 49–70. [Google Scholar]
- Wisti Lassen, A. The trade in wool in Old Assyrian Anatolia. Jaarber. Oriente Lux 2010, 42, 159–179. [Google Scholar]
- Breniquet, C. Weaving in Mesopotamia during the Bronze Age: Archaeology, techniques, iconography. In Textile Terminologies in the Ancient Near East and Mediterranean from the Third to the First Millennnia BC; Michel, C., Nosch, M.L., Eds.; Oxbow Books: Oxford, UK, 2010; pp. 52–67. [Google Scholar]
- Ryder, M.L. The human development of different fleece types in sheep. In Animals and Man in the Past: Essays in Honour of A.T. Clason Emeritus Professor of Archaeozoology Rijksuniversiteit Groningen, the Netherlands; Buitenhuis, H., Prummel, W., Eds.; Archaeological Research and Consultancy: Groningen, The Netherlands, 2001; pp. 67–73. [Google Scholar]
- Demars, J.; Cano, M.; Drouilhet, L.; Plisson-Petit, F.; Bardou, P.; Fabre, S.; Servin, B.; Sarry, J.; Woloszyn, F.; Mulsant, P.; et al. Genome-wide identification of the mutation underlying fleece variation and discriminating ancestral hairy species from modern woolly sheep. Mol. Biol. Evol. 2017, 34, 1722–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabatini, S.; Bergerbrant, S.; Brandt, L.; Margaryan, A.; Allentoft, M.E. Approaching sheep herds origins and the emergence of the wool economy in continental Europe during the Bronze Age. Archaeol. Anthr. Sci. 2019, 11, 4909–4925. [Google Scholar] [CrossRef] [Green Version]
- Chessa, B.; Pereira, F.; Arnaud, F.; Amorim, A.; Goyache, F.; Mainland, I.; Kao, R.R.; Pemberton, J.M.; Beraldi, D.; Stear, M.J.; et al. Revealing the history of sheep domestication using retrovirus integrations. Science 2009, 324, 532–536. [Google Scholar] [CrossRef] [Green Version]
- Ryder, M.L. The Evolution of the Fleece. Sci. Am. 1987, 256, 112–119. [Google Scholar] [CrossRef]
- Deng, J.; Xie, X.-L.; Wang, D.-F.; Zhao, C.; Lv, F.-H.; Li, X.; Yang, J.; Yu, J.-L.; Shen, M.; Gao, L.; et al. Paternal origins and migratory episodes of domestic sheep. Curr. Biol. 2020, 30, 4085–4095.e6. [Google Scholar] [CrossRef]
- Brunson, K.; He, N.; Dai, X. Sheep, cattle, and specialization: New zooarchaeological perspectives on the Taosi Longshan. Int. J. Osteoarchaeol. 2016, 26, 460–475. [Google Scholar] [CrossRef]
- Cai, D.; Zhang, N.; Shao, X.; Sun, W.; Zhu, S.; Yang, D.Y. New ancient DNA data on the origins and spread of sheep and cattle in northern China around 4000 BP. Asian Archaeol. 2018, 2, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Hermes, T.R.; Tishkin, A.A.; Kosintsev, P.A.; Stepanova, N.F.; Krause-Kyora, B.; Makarewicz, C.A. Mitochondrial DNA of domesticated sheep confirms pastoralist component of Afanasievo subsistence economy in the Altai Mountains (3300–2900 cal BC). Archaeol. Res. Asia 2020, 24, 100232. [Google Scholar] [CrossRef]
- Taylor, W.T.T.; Pruvost, M.; Posth, C.; Rendu, W.; Krajcarz, M.T.; Abdykanova, A.; Brancaleoni, G.; Spengler, R.; Hermes, T.; Schiavinato, S.; et al. Evidence for early dispersal of domestic sheep into Central Asia. Nat. Hum. Behav. 2021, 5, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Pulak, C. The Uluburun shipwreck and Late Bronze Age trade. In Beyond Babylon: Art, Trade, and Diplomacy in the Second Millennium BC; Aruz, J., Benzel, K., Evans, J.M., Eds.; Metropolitan Museum of Art: New York, NY, USA, 2008; pp. 288–310. [Google Scholar]
- Sherratt, A. The Trans-Eurasian Exchange: The prehistory of Chinese relations with the West. In Contact and Exchange in the Ancient World; Mair, V.H., Ed.; University of Hawaii Press: Honolulu, HI, USA, 2006; pp. 30–61. [Google Scholar] [CrossRef]
- Broodbank, C. The Making of the Middle Sea: A History of the Mediterranean from the Beginning to the Emergence of the Classical World; Thames & Hudson: London, UK, 2013. [Google Scholar]
- Vandkilde, H. Bronzization: The Bronze Age as pre-modern globalization. Praehistorische Z. 2016, 91, 103–123. [Google Scholar] [CrossRef]
- Dong, G.; Du, L.; Wei, W. The impact of early trans-Eurasian exchange on animal utilization in northern China during 5000–2500 BP. Holocene 2021, 31, 294–301. [Google Scholar] [CrossRef]
- Ryder, M.L. Coat structure and seasonal shedding in goats. Anim. Sci. 1966, 8, 289–302. [Google Scholar] [CrossRef]
- O’Connell, D.; Scobie, D.R.; Hickey, S.; Sumner, R.M.W.; Pearson, A.J. Selection for yearling fleece weight and its effect on fleece shedding in New Zealand Wiltshire sheep. Anim. Prod. Sci. 2012, 52, 456–462. [Google Scholar] [CrossRef]
- Vargas Jurado, N.; Leymaster, K.; Kuehn, L.; Lewis, R. Estimating heritability of wool shedding in a cross-bred ewe population. J. Anim. Breed. Genet. 2016, 133, 396–403. [Google Scholar] [CrossRef]
- Lopez, R. The origin of the Merino sheep. Jew. Soc. Stud. 1953, 5, 161–168. [Google Scholar]
- Klein, J. The Mesta: A Study in Spanish Economic History, 1273–1836; Kennikat Press: Port Washington, NY, USA, 1964. [Google Scholar]
- Pollard, S. Fixed capital in the Industrial Revolution in Britain. J. Econ. Hist. 1964, 24, 299–314. [Google Scholar] [CrossRef]
- Berg, M.; Hudson, P. Rehabilitating the industrial revolution. Econ. Hist. Rev. 1992, 45, 24–50. [Google Scholar] [CrossRef]
- Bruland, K. Industrialisation and technological change. In The Cambridge Economic History of Modern Britain; Floud, R., Johnson, P., Eds.; Cambridge University Press: Cambridge, UK, 2004; Volume 1, pp. 117–146. [Google Scholar] [CrossRef]
- Rossi, C.; Ruß-Popa, G.; Mattiangeli, V.; McDaid, F.; Hare, A.J.; Davoudi, H.; Laleh, H.; Lorzadeh, Z.; Khazaeli, R.; Fathi, H.; et al. Exceptional ancient DNA preservation and fibre remains of a Sasanian saltmine sheep mummy in Chehrābād, Iran. Biol. Lett. 2021, 17, 20210222. [Google Scholar] [CrossRef]
- Ferguson, K.; Carter, H.; Hardy, M.H. Studies of comparative fleece growth in sheep I. The quantitative nature of inherent differences in wool-growth rate. Aust. J. Biol. Sci. 1949, 2, 42–82. [Google Scholar] [CrossRef]
- Morris, L.R. Photoperiodicity of seasonal rhythm of wool growth in sheep. Nature 1961, 190, 102–103. [Google Scholar] [CrossRef] [PubMed]
- Slee, J.; Carter, H.B. A comparative study of fleece growth in Tasmanian Fine Merino and Wiltshire Horn ewes. J. Agric. Sci. 1961, 57, 11–19. [Google Scholar] [CrossRef]
- Bennett, J.W.; Hutchinson, J.C.D.; Wodzicka-Tomaszewska, M. Annual rhythm of wool growth. Nature 1962, 194, 651–652. [Google Scholar] [CrossRef]
- Hart, D.S.; Bennett, J.W.; Hutchinson, J.C.D.; Wodzicka-Tomaszewska, M. Reversed photoperiodic seasons and wool growth. Nature 1963, 198, 310–311. [Google Scholar] [CrossRef]
- Elsherbiny, A.A.; Eloksh, H.A.; Elsheikh, A.S.; Khalil, M.H. Effect of light and temperature on wool growth. J. Agric. Sci. 1978, 90, 329–334. [Google Scholar] [CrossRef]
- Gonzalez, E.B.; Sacchero, D.M.; Easdale, M.H. Environmental influence on Merino sheep wool quality through the lens of seasonal variations in fibre diameter. J. Arid Environ. 2020, 181, 104248. [Google Scholar] [CrossRef]
- Elsherbiny, A.; Markotić, B. Seasonality in wool growth under semi-arid conditions. J. Agric. Sci. 1974, 83, 443–446. [Google Scholar] [CrossRef]
- Matsoukis, A.; Chronopoulou-Sereli, A.; Stratakos, G. A comparative study of simple regression models to estimate fibre length growth in Chios sheep from common meteorological variables. Curr. Agric. Res. J. 2020, 8, 187–192. [Google Scholar] [CrossRef]
- Brochier, J.É. The use and abuse of culling profiles in recent zooarchaeological studies: Some methodological comments on “frequency correction” and its consequences. J. Archaeol. Sci. 2013, 40, 1416–1420. [Google Scholar] [CrossRef]
- Gerbault, P.; Gillis, R.; Vigne, J.-D.; Tresset, A.; Bréhard, S.; Thomas, M.G. Statistically robust representation and comparison of mortality profiles in archaeozoology. J. Archaeol. Sci. 2016, 71, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Gifford-Gonzalez, D. An Introduction to Zooarchaeology; Springer: Cham, Switzerland, 2018; pp. 480–481. [Google Scholar] [CrossRef]
- Barfield, T.J. The Central Asian Arabs of Afghanistan: Pastoral Nomadism in Transition; University of Texas Press: Austin, TX, USA, 1981. [Google Scholar]
- Gillis, R.E.; Gaastra, J.S.; Linden, M.V.; Vigne, J.-D. A species specific investigation into sheep and goat husbandry during the early European Neolithic. Environ. Archaeol. 2019, 27, 8–19. [Google Scholar] [CrossRef]
- Arbuckle, B.S. Early history of animal domestication in Southwest Asia. In Oxford Research Encyclopedia of Environmental Science; Shugart, H.H., Ed.; Oxford University Press: Oxford, UK, 2018. [Google Scholar] [CrossRef]
- Sierra, A.; Bréhard, S.; Montes, L.; Utrilla, P.; Saña, M. Sheep exploitation and husbandry in first farming societies: From production to consumption in Central Pyrenees in the Early Neolithic. Archaeol. Anthr. Sci. 2019, 11, 5813–5829. [Google Scholar] [CrossRef]
- Gourichon, L.; Horwitz, L.K. An Inter-Regional Comparison of Animal Domestication in the Northern and Southern Levant. Food Hist. 2021, 19, 53–78. [Google Scholar] [CrossRef]
- Stiner, M.C.; Munro, N.D.; Buitenhuis, H.; Duru, G.; Özbaşaran, M. An endemic pathway to sheep and goat domestication at Aşıklı Höyük (Central Anatolia, Turkey). Proc. Natl. Acad. Sci. USA 2022, 119, e2110930119. [Google Scholar] [CrossRef]
- Green, M.W. Animal Husbandry at Uruk in the Archaic Period. J. Near East. Stud. 1980, 39, 1–35. [Google Scholar] [CrossRef]
- Van de Mieroop, M. Sheep and goat herding according to the Old Babylonian texts from Ur. In Domestic Animals of Mesopotamia, Part I: Bulletin on Sumerian Agriculture; Powell, M., Postgate, J., Eds.; Sumerian Agriculture Group: Cambridge, UK, 1993; Volume 7, pp. 161–182. [Google Scholar]
- Rappaport, R.A. Ritual regulation of environmental relations among a New Guinea people. Ethnology 1967, 6, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Ekroth, G.; Wallensten, J. (Eds.) Bones, Behaviour and Belief: The Zooarchaeological Evidence as a Source for Ritual Practice in Ancient Greece and Beyond; Svenska Institutet i Athen: Stockholm, Sweden, 2013. [Google Scholar]
- Porter, A.; Schwartz, G. (Eds.) Sacred Killing: The Archaeology of Sacrifice in the Ancient Near East; Eisenbrauns: Winona Lake, IN, USA, 2012. [Google Scholar]
- Henton, E.; McCorriston, J.; Martin, L.; Oches, E.A. Seasonal aggregation and ritual slaughter: Isotopic and dental microwear evidence for cattle herder mobility in the Arabian Neolithic. J. Anthr. Archaeol. 2014, 33, 119–131. [Google Scholar] [CrossRef]
- Maher, E.F. Lambs to the Slaughter: Cultic Orientations at Philistine Ekron in the 7th Century BCE. In Material Culture Matters: Essays on the Archaeology of the Southern Levant in Honor of Seymour Gitin; Spencer, J.R., Mullins, R.A., Brody, A.J., Eds.; Eisenbrauns: Winona Lake, IN, USA, 2014; pp. 111–130. [Google Scholar]
- Marom, N.; Garfinkel, Y.; Bar-Oz, G. Times in between: A zooarchaeological analysis of ritual in Neolithic Sha’ar Hagolan. Quat. Int. 2018, 464, 216–225. [Google Scholar] [CrossRef]
- O’Connor, T.P. On the difficulty of detecting seasonal slaughtering of sheep. Environ. Archaeol. 1998, 3, 5–11. [Google Scholar] [CrossRef]
- Helmer, D.; Gourichon, L.; Vila, E. The development of the exploitation of products from Capra and Ovis (meat, milk and fleece) from the PPNB to the Early Bronze in the northern Near East (8700 to 2000 BC cal.). Anthropozoologica 2007, 42, 41–69. [Google Scholar]
- Sasson, A. Animal Husbandry in Ancient Israel: A Zooarchaeological Perspective on Livestock Exploitation, Herd Management and Economic Strategies; Routledge: London, UK, 2016. [Google Scholar]
- Daly, K.G.; Mattiangeli, V.; Hare, A.J.; Davoudi, H.; Fathi, H.; Doost, S.B.; Amiri, S.; Khazaeli, R.; Decruyenaere, D.; Nokandeh, J.; et al. Herded and hunted goat genomes from the dawn of domestication in the Zagros Mountains. Proc. Natl. Acad. Sci. USA 2021, 118, e2100901118. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.M. Agricultural Innovation in the Early Islamic World: The Diffusion of Crops and Farming Techniques, 700–1100; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Decker, M. Tilling the Hateful Earth: Agricultural Production and Trade in the Late Antique East; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Steger, M.B. Globalization: A Very Short Introduction; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Harris, D.R. An evolutionary continuum of people-plant interaction. In Foraging and Farming: The Evolution of Plant Exploitation; Harris, D.R., Hillman, G.C., Eds.; Unwin and Hyman: London, UK, 1989; pp. 11–26. [Google Scholar]
- Bar-Yosef, O. On the nature of transitions: The Middle to Upper Palaeolithic and the Neolithic Revolution. Camb. Archaeol. J. 1998, 8, 141–163. [Google Scholar] [CrossRef]
- Snir, A.; Nadel, D.; Groman-Yaroslavski, I.; Melamed, Y.; Sternberg, M.; Bar-Yosef, O.; Weiss, E. The origin of cultivation and proto-weeds, long before Neolithic farming. PLoS ONE 2015, 10, e0131422. [Google Scholar] [CrossRef] [Green Version]
- Mahaut, L.; Cheptou, P.-O.; Fried, G.; Munoz, F.; Storkey, J.; Vasseur, F.; Violle, C.; Bretagnolle, F. Weeds: Against the rules? Trends Plant Sci. 2020, 25, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, M.M.; Lochab, A.K.; Turley, N.E.; Johnson, M.T.J. Plant domestication slows pest evolution. Ecol. Lett. 2015, 18, 907–915. [Google Scholar] [CrossRef]
- Palmer, C. An exploration of the effects of crop rotation regime on modem weed floras. Environ. Archaeol. 1998, 2, 35–48. [Google Scholar] [CrossRef]
- Bogaard, A. Neolithic Farming in Central Europe; Routledge: London, UK, 2004. [Google Scholar]
- Bogaard, A.; Fraser, R.; Heaton, T.H.; Wallace, M.; Vaiglova, P.; Charles, M.; Jones, G.; Evershed, R.P.; Styring, A.K.; Andersen, N.H.; et al. Crop manuring and intensive land management by Europe’s first farmers. Proc. Natl. Acad. Sci. USA 2013, 110, 12589–12594. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.; Charles, M.; Bogaard, A.; Hodgson, J. Crops and weeds: The role of weed functional ecology in the identification of crop husbandry methods. J. Archaeol. Sci. 2010, 37, 70–77. [Google Scholar] [CrossRef]
- Bogaard, A.; Jacomet, S.; Schibler, J. Towards an integrated bioarchaeological perspective on the central European Neolithic: Understanding the pace and rhythm of social processes through comparative discussion of the western loess belt and Alpine foreland. In The Neolithic of Europe: Papers in Honour of Alasdair Whittle; Bickle, P., Cummings, V., Hofmann, D., Pollard, J., Eds.; Oxbow Books: Oxford, UK; Philadelphia, PA, USA, 2017; pp. 120–144. [Google Scholar] [CrossRef]
- Childe, V.G. What Happened in History; Penguin Books: London, UK, 1954. [Google Scholar]
- Kuijt, I. Demography and storage systems during the Southern Levantine Neolithic demographic transition. In The Neolithic Demographic Transition and its Consequences; Bocquet-Appel, J.P., Bar-Yosef, O., Eds.; Springer: Dordrecht, The Netherlands, 2008. [Google Scholar] [CrossRef]
- Kuijt, I. The Neolithic refrigerator on a Friday night: How many people are coming to dinner and just what should I do with the slimy veggies in the back of the fridge? Environ. Archaeol. 2015, 20, 321–336. [Google Scholar] [CrossRef]
- de Saulieu, G.; Testart, A. Innovations, food storage and the origins of agriculture. Environ. Archaeol. 2015, 20, 314–320. [Google Scholar] [CrossRef]
- Testart, A. The significance of food storage among hunter-gatherers: Residence patterns, population densities, and social inequalities. Curr. Anthropol. 1982, 23, 523–537. [Google Scholar] [CrossRef]
- Kuijt, I.; Finlayson, B. Evidence for food storage and predomestication granaries 11,000 years ago in the Jordan Valley. Proc. Natl. Acad. Sci. USA 2009, 106, 10966–10970. [Google Scholar] [CrossRef] [Green Version]
- Colledge, S.; Conolly, J.; Finlayson, B.; Kuijt, I. New insights on plant domestication, production intensification, and food storage: The archaeobotanical evidence from PPNA Dhra‘. Levant 2018, 50, 14–31. [Google Scholar] [CrossRef]
- Belfer-Cohen, A.; Bar-Yosef, O. Early sedentism in the Near East: A bumpy ride to village life. In Life in Neolithic Farming Communities: Social Organization, Identity and Differentiation; Kuijt, I., Ed.; Kluwer Academic: New York, NY, USA, 2000; pp. 19–37. [Google Scholar]
- Ladizinsky, G. Plant Evolution under Domestication; Kluwer Academic: Dordrecht, The Netherlands, 1998; pp. 32–39. [Google Scholar] [CrossRef]
- Bar-Yosef, O.; Belfer-Cohen, A. The origins of sedentism and farming communities in the Levant. J. World Prehistory 1989, 3, 447–498. [Google Scholar] [CrossRef]
- Yesner, D.R. Seasonality and resource “stress” among hunter-gatherers: Archaeological signatures. In Key Issues in Hunter-Gatherer Research; Burch, E.S., Ellanna, L.J., Eds.; Routledge: London, UK, 2020; pp. 151–167. [Google Scholar]
- Clauss, M.; Hume, I.D.; Hummel, J. Evolutionary adaptations of ruminants and their potential relevance for modern production systems. Animal 2010, 4, 979–992. [Google Scholar] [CrossRef] [Green Version]
- PERLIFE: Engineering Perennial Barley. European Research Council Grant 101002085. Available online: https://cordis.europa.eu/project/id/101002085 (accessed on 20 March 2022).
- Windes, S.; Carrijo, D.; Curwen-McAdams, C.; Hayes, P. Improving the sustainability of malting barley production: Prospects for perennial and annual growth habit varieties. Crop Sci. 2019, 59, 2289–2296. [Google Scholar] [CrossRef]
- Lister, D.L.; Bower, M.A.; Jones, M.K. Herbarium specimens expand the geographical and temporal range of germplasm data in phylogeographic studies. Taxon 2010, 59, 1321–1323. [Google Scholar] [CrossRef]
- Ellis, E.C.; Kaplan, J.O.; Fuller, D.Q.; Vavrus, S.; Goldewijk, K.K.; Verburg, P.H. Used planet: A global history. Proc. Natl. Acad. Sci. USA 2013, 110, 7978–7985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquer, L.; Gaillard, M.-J.; Sugita, S.; Poska, A.; Trondman, A.-K.; Mazier, F.; Nielsen, A.B.; Fyfe, R.M.; Jönsson, A.M.; Smith, B.; et al. Quantifying the effects of land use and climate on Holocene vegetation in Europe. Quat. Sci. Rev. 2017, 171, 20–37. [Google Scholar] [CrossRef] [Green Version]
- Rosen, S.A. Basic Instabilities? Climate and culture in the Negev over the long term. Geoarchaeology 2017, 32, 6–22. [Google Scholar] [CrossRef]
- Rosen, S.A. Trade through the desert: A long-term perspective on goods, animals, and polities in the Negev. Chungara 2019, 51, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Rosen, S.A. Hunting to herding to trading to warfare: A chronology of animal exploitation in the Negev. In Animals and Human Society in Asia; Kowner, R., Bar-Oz, G., Biran, M., Shahar, M., Shelach-Lavi, G., Eds.; Palgrave Macmillan: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Borcan, O.; Olsson, O.; Putterman, L. Transition to agriculture and first state presence: A global analysis. Explor. Econ. Hist. 2021, 82, 101404. [Google Scholar] [CrossRef]
- Chase-Dunn, C. Globalization: A world-systems perspective. J. World-Syst. Res. 1999, 5, 187–215. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, K.H.; Williamson, J.G. When did globalisation begin? Eur. Rev. Econ. Hist. 2002, 6, 23–50. [Google Scholar] [CrossRef] [Green Version]
- Gills, B.K.; Thompson, W. (Eds.) Globalization and Global History; Routledge: London, UK; New York, NY, USA, 2006. [Google Scholar]
- McKeown, A. Periodizing Globalization. Hist. Work. J. 2007, 63, 218–230. [Google Scholar] [CrossRef]
- Nederveen Pieterse, J. Periodizing globalization: Histories of globalization. New Glob. Stud. 2012, 6. [Google Scholar] [CrossRef]
- Wallerstein, E. The Modern World-System, Vol. 1: Capitalist Agriculture and the Origins of the European World-Economy in the Sixteenth Century; Academic Press: New York, NY, USA, 1974. [Google Scholar]
- Frank, A.G.; Gills, B.K. (Eds.) The World System: Five Hundred Years or Five Thousand? Routledge: London, UK, 1993. [Google Scholar]
- Woolf, G. World-systems analysis and the Roman empire. J. Rom. Archaeol. 1990, 3, 44–58. [Google Scholar] [CrossRef]
- Hingley, R. Globalizing Roman culture: Unity, diversity and Empire; Routledge: London, UK, 2005. [Google Scholar] [CrossRef]
- LaBianca, O.S.; Arnold Scham, S. Connectivity in Antiquity: Globalization as a Long-Term Historical Process; Routledge: London, UK, 2006. [Google Scholar] [CrossRef]
- Jennings, J. Globalizations and the Ancient World; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Pitts, M.; Versluys, M.J. (Eds.) Globalisation and the Roman World: World History, Connectivity and Material Culture; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Hodos, T. (Ed.) The Routledge Handbook of Archaeology and Globalization; Routledge: London, UK, 2017. [Google Scholar]
- Boivin, N.; Frachetti, M.D. Globalization in Prehistory: Contact, Exchange, and the ‘People without History’; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]
- Autiero, S.; Cobb, M.A. (Eds.) Globalization and Transculturality from Antiquity to the Pre-Modern World; Routledge: London, UK, 2021. [Google Scholar] [CrossRef]
- Jones, M.; Hunt, H.; Lightfoot, E.; Lister, D.; Liu, X.; Motuzaite-Matuzeviciute, G. Food globalization in prehistory. World Archaeol. 2011, 43, 665–675. [Google Scholar] [CrossRef]
- Liu, X.; Jones, M.K. Food globalisation in prehistory: Top down or bottom up? Antiquity 2014, 88, 956–963. [Google Scholar] [CrossRef]
- Boivin, N.L.; Fuller, D.Q.; Crowther, A. Old World globalization and food exchanges. In Archaeology of Food: An Encyclopedia, Volume 2: L-Z; Metheny, K.B., Beaudry, M.C., Eds.; Rowman & Littlefield: Lanham, MD, USA, 2015; pp. 350–356. [Google Scholar]
- Jones, M.; Hunt, H.; Kneale, C.; Lightfoot, E.; Lister, D.; Liu, X.; Motuzaite-Matuzeviciute, G. Food globalisation in prehistory: The agrarian foundations of an interconnected continent. J. Br. Acad. 2016, 4, 73–87. [Google Scholar] [CrossRef]
- Lee, G.A. The spread of domesticated plant resources in prehistoric northeast Asia. In The Routledge Handbook of Archaeology and Globalization; Hodos, T., Ed.; Routledge: London, UK, 2017; pp. 394–412. [Google Scholar]
- Van der Veen, M.; Morales, J. Food Globalisation and the Red Sea: New Evidence from the Ancient Ports at Quseir al-Qadim, Egypt. In Human Interaction with the Environment in the Red Sea; Agius, D.A., Khalil, E., Scerri, E.M.L., Williams, A., Eds.; Brill: Leiden, The Netherlands, 2017; pp. 254–289. [Google Scholar]
- Jacobs, C.; Berglund, M.; Kurnik, B.; Dworak, T.; Marras, S.; Mereu, V.; Michetti, M. Climate Change Adaptation in the Agriculture Sector in Europe (No. 4/2019); European Environment Agency (EEA); Publications Office of the European Union: Luxembourg, 2019; Available online: https://euroseeds.eu/app/uploads/2019/09/Climate-change-adaptation-in-the-agriculture-sector-in-Europe.pdf (accessed on 20 March 2022).



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuks, D.; Lister, D.L.; Distelfeld, A.; Marom, N. A Time to Sow, a Time to Reap: Modifications to Biological and Economic Rhythms in Southwest Asian Plant and Animal Domestication. Agronomy 2022, 12, 1368. https://doi.org/10.3390/agronomy12061368
Fuks D, Lister DL, Distelfeld A, Marom N. A Time to Sow, a Time to Reap: Modifications to Biological and Economic Rhythms in Southwest Asian Plant and Animal Domestication. Agronomy. 2022; 12(6):1368. https://doi.org/10.3390/agronomy12061368
Chicago/Turabian StyleFuks, Daniel, Diane L. Lister, Assaf Distelfeld, and Nimrod Marom. 2022. "A Time to Sow, a Time to Reap: Modifications to Biological and Economic Rhythms in Southwest Asian Plant and Animal Domestication" Agronomy 12, no. 6: 1368. https://doi.org/10.3390/agronomy12061368