
Foliar Brassinolide Sprays Ameliorate Post-Silking Heat Stress on the Accumulation and Remobilization of Biomass and Nitrogen in Fresh Waxy Maize


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Yield Determination
2.3. Accumulation and Remobilization of Biomass and Nitrogen
- Post-silking biomass (nitrogen) accumulation = biomass (nitrogen) accumulation at the fresh stage − biomass (nitrogen) accumulation at the silking stage;
- Biomass (nitrogen) remobilization amount from pre-silking leaf (stem) to grain (REP) = biomass (nitrogen) accumulation amount in leaf (stem) at the silking stage − biomass (nitrogen) accumulation amount in leaf (stem) at the fresh stage;
Biomass (nitrogen) remobilization efficiency (%, REE) =100 × biomass (nitrogen) remobilization from pre-silking leaf (stem) to grain biomass (nitrogen) accumulation in leaf (stem) at the silking stage - Remobilization amount of biomass (nitrogen) of post-silking vegetative organ photosynthate = grain dry weight at the fresh stage−biomass (nitrogen) remobilization from pre-silking vegetative organs to grain;
Harvest index (HI, %) =100 × grain dry weight biomass at the fresh stage Nitrogen HI (NHI, %) =100 × nitrogen accumulation in grain at the fresh stage nitrogen accumulation in total plants at the fresh stage
2.4. Statistical Analysis
3. Results and Discussion
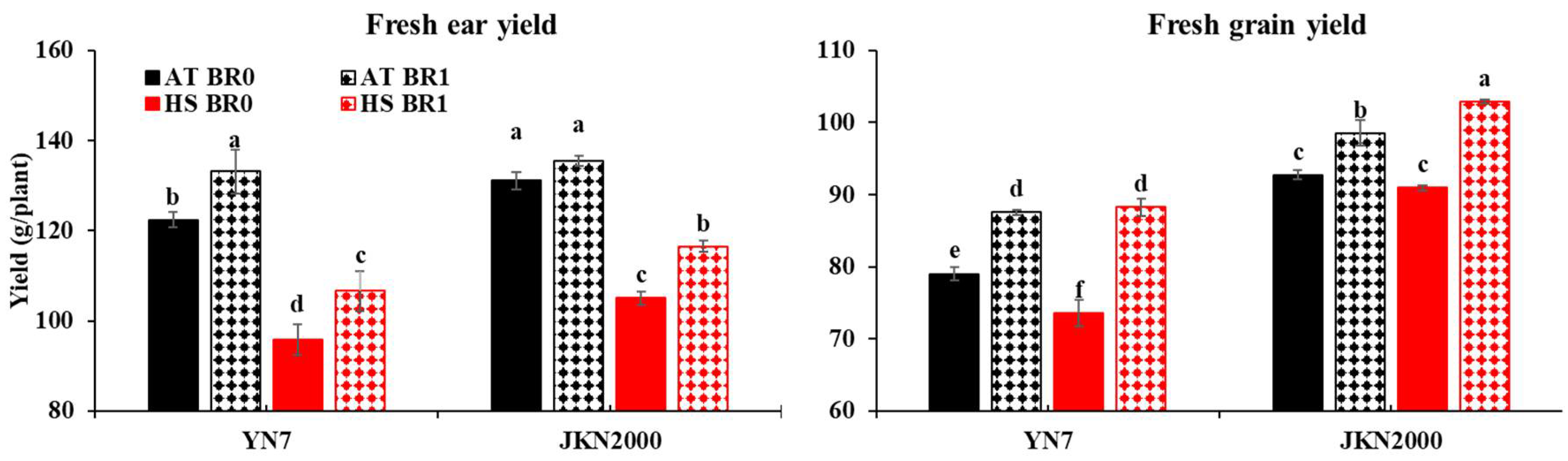
3.1. Ear and Grain Yield
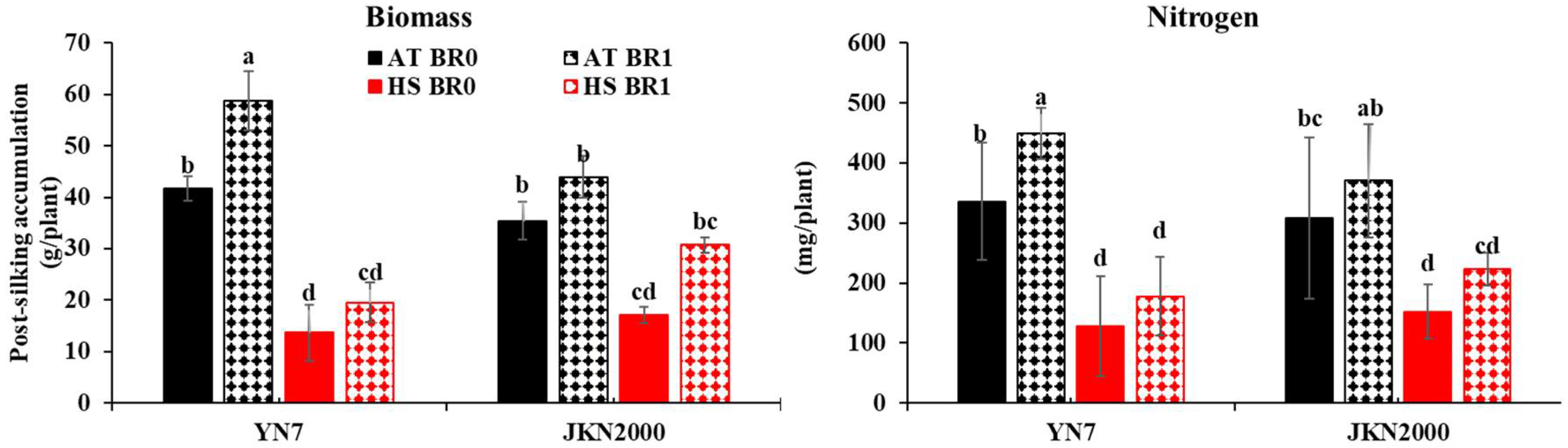
3.2. Post-Silking Biomass and Nitrogen Accumulation
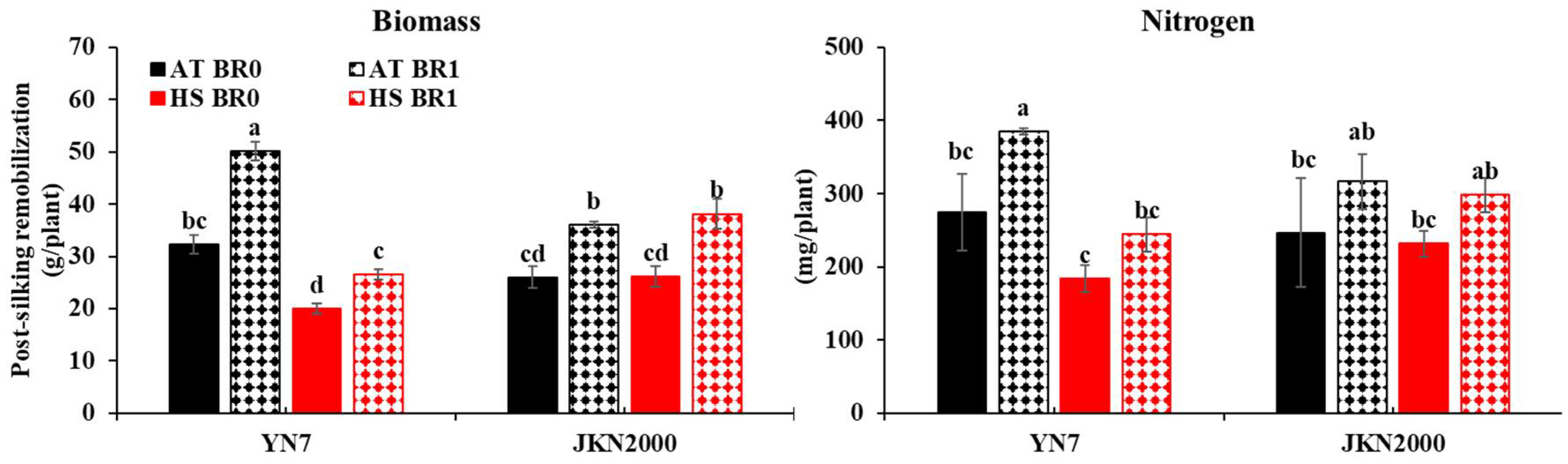
3.3. Remobilization of Post-Silking Biomass and Nitrogen
3.4. Remobilization of Pre-Silking Biomass and Nitrogen
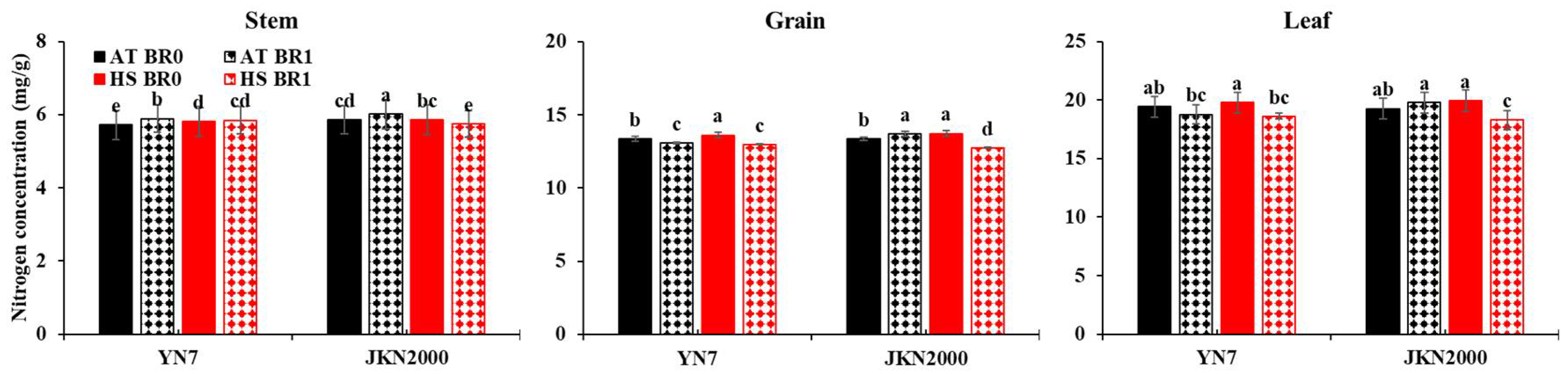
3.5. Nitrogen Concentration in Stem, Leaf, and Grain
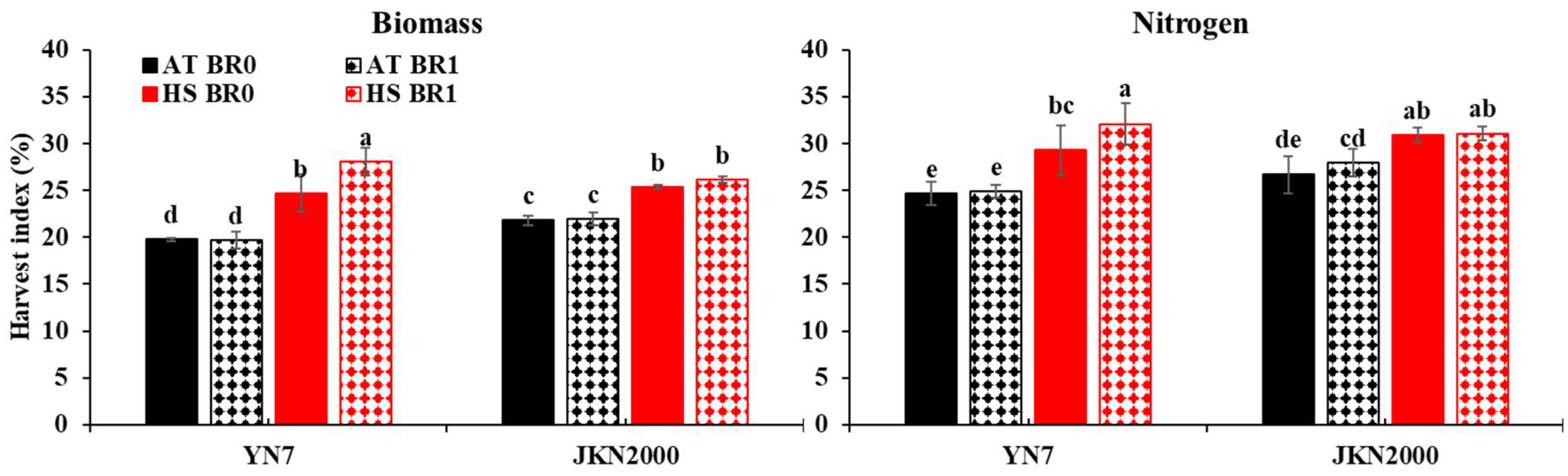
3.6. Harvest Index of Biomass and Nitrogen
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goraya, G.K.; Kaur, B.; Asthir, B.; Bala, S.; Kaur, G.; Farooq, M. Rapid injuries of high temperature in plants. J. Plant Biol. 2017, 60, 298–305. [Google Scholar] [CrossRef]
- Lohani, N.; Singh, M.B.; Bhalla, P.L. High temperature susceptibility of sexual reproduction in crop plants. J. Exp. Bot. 2020, 71, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Li, B.J.; Gao, K.; Ren, H.M.; Tang, W.Q. Molecular mechanisms governing plant responses to high temperatures. J. Integr. Plant Biol. 2018, 60, 757–779. [Google Scholar] [CrossRef]
- Tiwari, Y.K.; Yadav, S.K. High temperature stress tolerance in maize (Zea mays L.): Physiological and molecular mechanisms. J. Plant Biol. 2019, 62, 93–102. [Google Scholar] [CrossRef]
- Wei, S.B.; Liu, J.; Li, T.T.; Wang, X.Y.; Peng, A.C.; Chen, C.Q. Effect of high-temperature events when heading into the maturity period on summer maize (Zea mays L.) yield in the Huang-Huai-Hai region, China. Atmosphere 2020, 11, 1291. [Google Scholar] [CrossRef]
- Tian, B.; Talukder, S.K.; Fu, J.M.; Fritz, A.K.; Trick, H.N. Expression of a rice soluble starch synthase gene in transgenic wheat improves the grain yield under heat stress conditions. Vitr. Cell. Dev. Biol. Plant 2018, 54, 216–227. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.R.; Goswami, S.; Shamim, M.; Mishra, U.; Jain, M.; Singh, K.; Singh, J.P.; Dubey, K.; Singh, S.; Rai, G.K.; et al. Biochemical defense response: Characterizing the plasticity of source and sink in spring wheat under terminal heat stress. Front. Plant Sci. 2017, 8, 1603. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.X.; Howell, S.H. Heat stress responses and thermotolerance in maize. Int. J. Mol. Sci. 2021, 22, 948. [Google Scholar] [CrossRef]
- Schaarschmidt, S.; Lawas, L.M.F.; Kopka, J.; Jagadish, S.V.K.; Zuther, E. Physiological and molecular attributes contribute to high night temperature tolerance in cereals. Plant Cell Environ. 2021, 44, 2034–2048. [Google Scholar] [CrossRef]
- Impa, S.M.; Sunoj, V.S.J.; Krassovskaya, I.; Bheemanahalli, R.; Obata, T.; Jagadish, S.V.K. Carbon balance and source-sink metabolic changes in winter wheat exposed to high night-time temperature. Plant Cell Environ. 2019, 42, 1233–1246. [Google Scholar] [CrossRef]
- Shirdelmoghanloo, H.; Cozzolino, D.; Lohraseb, I.; Collins, N.C. Truncation of grain filling in wheat (Triticum aestivum) triggered by brief heat stress during early grain filling: Association with senescence responses and reductions in stem reserves. Funct. Plant Biol. 2016, 43, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Huang, T.Q.; Ding, M.Q.; Lu, D.L.; Lu, W.P. High temperature during grain filling impacts on leaf senescence in waxy maize. Agron. J. 2017, 109, 906–916. [Google Scholar] [CrossRef]
- Ercoli, L.; Arduini, I.; Mariotti, M.; Masoni, A. Post-anthesis dry matter and nitrogen dynamics in durum wheat as affected by nitrogen and temperature during grain filling. Cereal Res. Commun. 2010, 38, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Larmure, A.; Munier-Jolain, N.G. High temperatures during the seed-filling period decrease seed nitrogen amount in pea (Pisum sativum L.): Evidence for a sink limitation. Front. Plant Sci. 2019, 10, 1608. [Google Scholar] [CrossRef]
- Zhen, F.X.; Zhou, J.J.; Mahmood, A.; Wang, W.; Chang, X.N.; Liu, B.; Liu, L.L.; Cao, W.X.; Zhu, Y.; Tang, L. Quantifying the effects of short-term heat stress at booting stage on nonstructural carbohydrates remobilization in rice. Crop J. 2020, 8, 194–212. [Google Scholar] [CrossRef]
- Li, S.M.; Zheng, H.X.; Lin, L.; Wang, F.; Sui, N. Roles of brassinosteroids in plant growth and abiotic stress response. Plant Growth Regul. 2021, 93, 29–38. [Google Scholar] [CrossRef]
- Yang, J.C.; Miao, W.Q.; Chen, J. Roles of jasmonates and brassinosteroids in rice responses to high temperature stress—A review. Crop J. 2021, 9, 977–985. [Google Scholar] [CrossRef]
- Sharma, I.; Kaur, N.; Pati, P.K. Brassinosteroids: A promising option in deciphering remedial strategies for abiotic stress tolerance in rice. Front. Plant Sci. 2017, 8, 2151. [Google Scholar] [CrossRef]
- Lv, J.H.; Dong, T.Y.; Zhang, Y.P.; Ku, Y.; Zheng, T.; Jia, H.F.; Fang, J.G. Metabolomic profiling of brassinolide and abscisic acid in response to high-temperature stress. Plant Cell Rep. 2022, 41, 935–946. [Google Scholar] [CrossRef]
- Chen, Y.H.; Chen, H.Z.; Xiang, J.; Zhang, Y.K.; Wang, Z.G.; Zhu, D.F.; Wang, J.K.; Zhang, Y.P.; Wang, Y.L. Rice spikelet formation inhibition caused by decreased sugar utilization under high temperature is associated with brassinolide decomposition. Environ. Exp. Bot. 2021, 190, 104585. [Google Scholar] [CrossRef]
- Yan, Y.N.; Wang, L.F.; Lu, D.L. Effects of spraying exogenous cytokinin or spermine on the starch physicochemical properties of waxy maize exposed to post-silking high temperature. J. Cereal Sci. 2020, 95, 103040. [Google Scholar] [CrossRef]
- Yang, H.; Lu, D.L.; Shen, X.; Cai, X.M.; Lu, W.P. Heat stress at different grain Filling stages affects fresh waxy maize grain yield and quality. Cereal Chem. 2015, 92, 258–264. [Google Scholar] [CrossRef]
- Wang, H.Q.; Liu, P.; Zhang, J.W.; Zhao, B.; Ren, B.Z. Endogenous hormones inhibit differentiation of young ears in maize (Zea mays L.) under heat stress. Front. Plant Sci. 2020, 11, 553046. [Google Scholar] [CrossRef]
- Lu, D.L.; Sun, X.L.; Yan, F.B.; Wang, X.; Xu, R.C.; Lu, W.P. Effects of heat stress at different grain-filling phases on the grain yield and quality of waxy maize. Cereal Chem. 2014, 91, 189–194. [Google Scholar] [CrossRef]
- Lu, D.L.; Cai, X.M.; Yan, F.B.; Sun, X.L.; Wang, X.; Lu, W.P. Effects of high temperature after pollination on physicochemical properties of waxy maize flour during grain development. J. Sci. Food Agric. 2014, 94, 1416–1421. [Google Scholar] [CrossRef]
- Gao, Z.; Liang, X.G.; Zhang, L.; Lin, S.; Zhao, X.; Zhou, L.L.; Shen, S.; Zhou, S.L. Spraying exogenous 6-benzyladenine and brassinolide at maize yield by enhancing source and sink capacity tasseling increases. Field Crops Res. 2017, 211, 1–9. [Google Scholar] [CrossRef]
- Kothari, A.; Lachowiec, J. Roles of brassinosteroids in mitigating heat stress damage in cereal crops. Int. J. Mol. Sci. 2021, 22, 2706. [Google Scholar] [CrossRef]
- Pantoja-Benavides, A.D.; Garces-Varon, G.; Restrepo-Diaz, H. Foliar growth regulator sprays induced tolerance to combined heat stress by enhancing physiological and biochemical responses in rice. Front. Plant Sci. 2021, 12, 702892. [Google Scholar] [CrossRef]
- Chen, Y.L.; Xiao, C.X.; Chen, X.C.; Li, Q.; Zhang, J.; Chen, F.J.; Yuan, L.X.; Mi, G.H. Characterization of the plant traits contributed to high grain yield and high grain nitrogen concentration in maize. Field Crops Res. 2014, 159, 1–9. [Google Scholar] [CrossRef]
- Osman, R.; Zhu, Y.; Cao, W.X.; Ding, Z.F.; Wang, M.; Liu, L.L.; Tang, L.; Liu, B. Modeling the effects of extreme high-temperature stress at anthesis and grain filling on grain protein in winter wheat. Crop J. 2021, 9, 889–900. [Google Scholar] [CrossRef]
- Ben-Asher, J.; Garcia, A.G.Y.; Hoogenboom, G. Effect of high temperature on photosynthesis and transpiration of sweet corn (Zea mays L. var. rugosa). Photosynthetica 2008, 46, 595–603. [Google Scholar] [CrossRef]
- Yang, H.; Gu, X.T.; Ding, M.Q.; Lu, W.P.; Lu, D.L. Weakened carbon and nitrogen metabolisms under post-silking heat stress reduce the yield and dry matter accumulation in waxy maize. J. Integr. Agr. 2020, 19, 78–88. [Google Scholar] [CrossRef]
- Vignjevic, M.; Wang, X.; Olesen, J.E.; Wollenweber, B. Traits in spring wheat cultivars associated with yield loss caused by a heat stress episode after anthesis. J. Agron. Crop Sci. 2015, 201, 32–48. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lim, S.S.; Kwak, J.H.; Lee, D.S.; Lee, S.M.; Ro, H.M.; Choi, W.J. Dry matter and nitrogen accumulation and partitioning in rice (Oryza sativa L.) exposed to experimental warming with elevated CO2. Plant Soil 2011, 342, 59–71. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Guan, D.H.; Pan, H.B.; Li, J.M.; Duan, L.S.; Zhang, M.C.; Li, Z.H. Effect of brassinolide on leaf photosynthetic function and yield in spring maize filling stage. Acta Agron. Sin. 2015, 41, 1557–1563. [Google Scholar] [CrossRef]
- Zang, J.P.; Zhao, A.J.; Zhao, Y.L.; Yan, Q.D.; Feng, J.J.; Zhang, H.L.; Wang, F.R.; Dong, J.G. The influence of brassinosteroid on the light catching, CO2 fixation and the translocation of organic compounds in maize leaves. Sci. Agric. Sin. 2017, 50, 4228–4234. [Google Scholar]
- Otie, V.; Ping, A.; Udo, I.; Eneji, E. Brassinolide effects on maize (Zea mays L.) growth and yield under waterlogged conditions. J. Plant Nutr. 2019, 42, 954–969. [Google Scholar] [CrossRef]
- Otie, V.; Ping, A.; Eneji, E. Interactive effect of brassinolide and lime on growth and yield of maize (Zea mays L.) on acid soils of South-East Nigeria. Commun. Soil Sci. Plant Anal. 2018, 49, 2918–2931. [Google Scholar] [CrossRef]
- Cheng, W.G.; Sakai, H.; Yagi, K.; Hasegawa, T. Combined effects of elevated CO2 and high night temperature on carbon assimilation, nitrogen absorption, and the allocations of C and N by rice (Oryza sativa L.). Agric. For. Meteorol. 2010, 150, 1174–1181. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, H.D.; Zhao, C.R.; Chen, G.H.; Zou, Y.B. Amino acid content in rice grains is affected by high temperature during the early grain-filling period. Sci. Rep. 2019, 9, 2700. [Google Scholar] [CrossRef]
- Rezaei, E.E.; Webber, H.; Gaiser, T.; Naab, J.; Ewert, F. Heat stress in cereals: Mechanisms and modelling. Eur. J. Agron. 2015, 64, 98–113. [Google Scholar] [CrossRef]
- Mu, X.H.; Chen, Q.W.; Chen, F.J.; Yuan, L.X.; Mi, G.H. Dynamic remobilization of leaf nitrogen components in relation to photosynthetic rate during grain filling in maize. Plant Physiol. Biochem. 2018, 129, 27–34. [Google Scholar] [CrossRef]
- Kong, L.A.; Xie, Y.; Hu, L.; Feng, B.; Li, S.D. Remobilization of vegetative nitrogen to developing grain in wheat (Triticum aestivum L.). Field Crops Res. 2016, 196, 134–144. [Google Scholar] [CrossRef]
- Liu, Q.H.; Ma, H.; Sun, Z.W.; Lin, X.Q.; Zhou, X.B. Translocation efficiencies and allocation of nitrogen, phosphorous and potassium in rice as affected by silicon fertilizer under high daytime temperature. J. Agron. Crop Sci. 2019, 205, 188–201. [Google Scholar] [CrossRef]
- Tahir, I.S.A.; Nakata, N. Remobilization of nitrogen and carbohydrate from stems of bread wheat in response to heat stress during grain filling. J. Agron. Crop Sci. 2005, 191, 106–115. [Google Scholar] [CrossRef]
- Ning, P.; Fritschi, F.B.; Li, C.J. Temporal dynamics of post-silking nitrogen fluxes and their effects on grain yield in maize under low to high nitrogen inputs. Field Crops Res. 2017, 204, 249–259. [Google Scholar] [CrossRef]
- Schittenhelm, S.; Langkamp-Wedde, T.; Kraft, M.; Kottmann, L.; Matschiner, K. Effect of two-week heat stress during grain filling on stem reserves, senescence, and grain yield of European winter wheat cultivars. J. Agron. Crop Sci. 2020, 206, 722–733. [Google Scholar] [CrossRef] [Green Version]
- Cosentino, S.L.; Sanzone, E.; Testa, G.; Patane, C.; Anastasi, U.; Scordia, D. Does post-anthesis heat stress affect plant phenology, physiology, grain yield and protein content of durum wheat in a semi-arid Mediterranean environment? J. Agron. Crop Sci. 2019, 205, 309–323. [Google Scholar] [CrossRef]
- Wang, Y.X.; Frei, M. Stressed food—The impact of abiotic environmental stresses on crop quality. Agric. Ecosyst. Environ. 2011, 141, 271–286. [Google Scholar] [CrossRef]
- Yamakawa, H.; Hakata, M. Atlas of rice grain filling-related metabolism under high temperature: Joint analysis of metabolome and transcriptome demonstrated inhibition of starch accumulation and induction of amino acid accumulation. Plant Cell Physiol. 2010, 51, 795–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, H.A.; Men, S.N.; Hussain, S.; Chen, Y.L.; Ali, S.; Zhang, S.; Zhang, K.P.; Li, Y.; Xu, Q.W.; Liao, C.Q.; et al. Interactive effects of drought and heat stresses on morphophysiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; Zhuang, Q.L.; Archontoulis, S.V.; Bernacchi, C.; Muller, C. Dissecting the nonlinear response of maize yield to high temperature stress with model-data integration. Glob. Chang. Biol. 2019, 25, 2470–2484. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hybrid | Temperature | BR | Stem | Leaf | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Biomass | Nitrogen | Biomass | Nitrogen | |||||||
| REP (g/plant) | REE (%) | REP (g/plant) | REE (%) | REP (g/plant) | REE (%) | REP (g/plant) | REE (%) | |||
| YN7 | AT | BR0 | −0.9 c | −1.6 d | 179.3 b | 36.4 cd | −1.4 a | −4.3 a | −51.0 b | −8.3 c |
| BR1 | −15.1 d | −27.9 c | 86.1 c | 17.5 e | −1.9 a | −5.8 a | −38.2 ab | −6.2 bc | ||
| HT | BR0 | 8.3 ab | 15.5 ab | 227.7 a | 46.2 ab | 2.0 a | 6.2 a | 4.9 ab | 0.8 abc | |
| BR1 | 10.5 a | 19.4 a | 237.8 a | 48.2 a | −0.8 a | −2.5 a | −13.4 ab | −2.2 abc | ||
| JKN2000 | AT | BR0 | 10.9 a | 16.5 ab | 256.4 a | 44.3 abc | −1.0 a | −2.5 a | −16.5 ab | −2.3 abc |
| BR1 | −0.2 c | −0.3 d | 180.9 b | 31.3 d | 2.3 a | 5.9 a | 25.9 ab | 3.5 ab | ||
| HT | BR0 | 7.5 ab | 11.4 bc | 236.6 a | 40.9 abc | 3.4 a | 8.9 a | 39.8 a | 5.5 a | |
| BR1 | 5.6 b | 8.4 c | 231.6 a | 40.0 bc | −1.8 a | −4.8 a | 5.2 ab | 0.7 abc | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Li, G.; Yang, H.; Lu, D. Foliar Brassinolide Sprays Ameliorate Post-Silking Heat Stress on the Accumulation and Remobilization of Biomass and Nitrogen in Fresh Waxy Maize. Agronomy 2022, 12, 1363. https://doi.org/10.3390/agronomy12061363
Zhang X, Li G, Yang H, Lu D. Foliar Brassinolide Sprays Ameliorate Post-Silking Heat Stress on the Accumulation and Remobilization of Biomass and Nitrogen in Fresh Waxy Maize. Agronomy. 2022; 12(6):1363. https://doi.org/10.3390/agronomy12061363
Chicago/Turabian StyleZhang, Xiaoyu, Guanghao Li, Huan Yang, and Dalei Lu. 2022. "Foliar Brassinolide Sprays Ameliorate Post-Silking Heat Stress on the Accumulation and Remobilization of Biomass and Nitrogen in Fresh Waxy Maize" Agronomy 12, no. 6: 1363. https://doi.org/10.3390/agronomy12061363