
Serine Hydroxymethyltransferase (SHMT) Gene Family in Wheat (Triticum aestivum L.): Identification, Evolution, and Expression Analysis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of the SHMT Family Genes
2.2. Phylogenetic Relationships, Gene Structures, and Domains Analysis
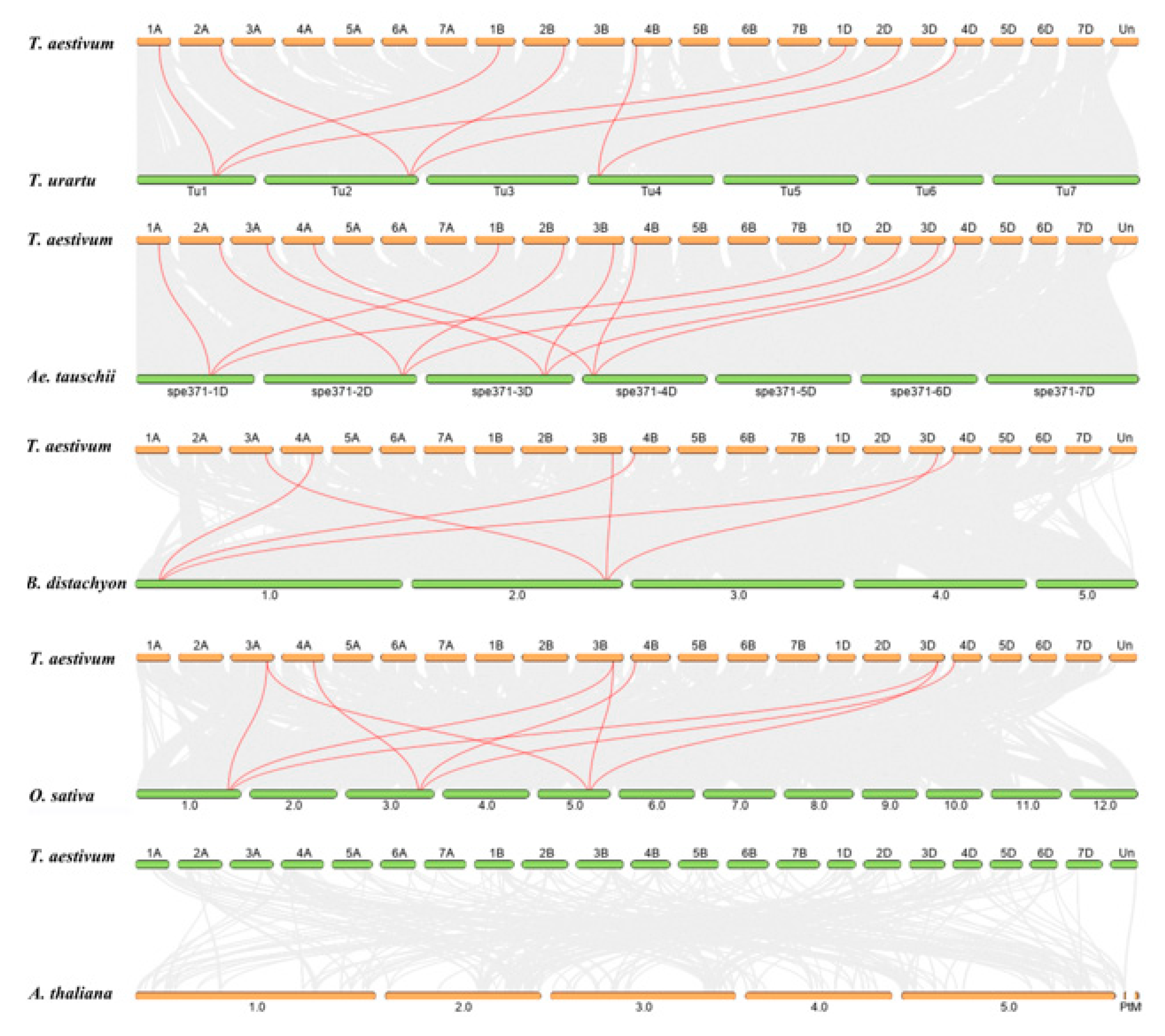
2.3. Chromosomal Location, Synteny, and Ka/Ks Analysis
2.4. GO Annotation and Protein Interactions Analysis
2.5. Cis-Element Analysis in the Promoter
2.6. Real-Time PCR Analysis
2.7. Stress Tolerance Assay in Yeast Cells
3. Results
3.1. Characteristics and Phylogenetic Analysis of TaSHMTs in T. aestivum
3.2. Chromosomal Distribution, Synteny, and Ka/Ks Analysis of TaSHMTs
3.3. Gene Structure and Conserved Motifs and Cis-Elements Analysis of TaSHMT Genes
3.4. Expression Patterns of TaSHMT Genes in Different Tissues
3.5. Expression Patterns of TaSHMT Genes under Abiotic Stress
3.6. TaSHMT2.1 Negatively Regulates Dehydration and Salt Stress Tolerance in Yeast Cells
3.7. GO Annotation and Protein–Protein Interactions of TaSHMTs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, P.K.; Balyan, H.S.; Sharma, S.; Kumar, R. Genetics of yield, abiotic stress tolerance and biofortification in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2020, 133, 1569–1602. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, K.; Sandoval, F.J.; Santiago, K.; Roje, S. One-carbon metabolism in plants: Characterization of a plastid serine hydroxymethyltransferase. Biochem. J. 2010, 430, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Mauve, C.; Lamothe Sibold, M.; Guérard, F.; Glab, N.; Hodges, M.; Jossier, M. Photorespiratory serine hydroxymethyltransferase 1 activity impacts abiotic stress tolerance and stomatal closure. Plant Cell Environ. 2019, 42, 2567–2583. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, N.H.; Liu, W.; Liu, G.; Selvaraj, G.; Wei, Y.; King, J. Transcriptional regulation of genes involved in the pathways of biosynthesis and supply of methyl units in response to powdery mildew attack and abiotic stresses in wheat. Plant Mol. Biol. 2007, 64, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Mcintyre, C.L.; Casu, R.E.; Rattey, A.; Dreccer, M.F.; Kam, J.W.; van Herwaarden, A.F.; Shorter, R.; Xue, G.P. Linked gene networks involved in nitrogen and carbon metabolism and levels of water-soluble carbohydrate accumulation in wheat stems. Funct. Integr. Genom. 2011, 11, 585–597. [Google Scholar] [CrossRef]
- Angelucci, F.; Morea, V.; Angelaccio, S.; Saccoccia, F.; Contestabile, R.; Ilari, A. The crystal structure of archaeal serine hydroxymethyltransferase reveals idiosyncratic features likely required to withstand high temperatures. Proteins Struct. Funct. Bioinform. 2014, 82, 3437–3449. [Google Scholar] [CrossRef]
- Mishra, P.; Jain, A.; Takabe, T.; Tanaka, Y.; Negi, M.; Singh, N.; Jain, N.; Mishra, V.; Maniraj, R.; Krishnamurthy, S.L.; et al. Heterologous expression of serine hydroxymethyltransferase-3 from rice confers tolerance to salinity stress in E. coli and arabidopsis. Front. Plant Sci. 2019, 10, 217. [Google Scholar] [CrossRef]
- Lakhssassi, N.; Patil, G.; Piya, S.; Zhou, Z.; Baharlouei, A.; Kassem, M.A.; Lightfoot, D.A.; Hewezi, T.; Barakat, A.; Nguyen, H.T.; et al. Genome reorganization of the GmSHMT gene family in soybean showed a lack of functional redundancy in resistance to soybean cyst nematode. Sci. Rep. 2019, 9, 1506. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.I.; Martin, R.; Castresana, C. Arabidopsis SHMT1, a serine hydroxymethyltransferase that functions in the photorespiratory pathway influences resistance to biotic and abiotic stress. Plant J. 2005, 41, 451–463. [Google Scholar] [CrossRef]
- Fang, C.; Zhang, P.; Li, L.; Yang, L.; Mu, D.; Yan, X.; Li, Z.; Lin, W. Serine hydroxymethyltransferase localised in the endoplasmic reticulum plays a role in scavenging H2O2 to enhance rice chilling tolerance. BMC Plant Biol. 2020, 20, 236. [Google Scholar] [CrossRef]
- Chen, J.; Huang, X.Y.; Salt, D.E.; Zhao, F.J. Mutation in OsCADT1 enhances cadmium tolerance and enriches selenium in rice grain. New Phytol. 2020, 226, 838–850. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Pan, X.; Wang, C.; Yun, F.; Huang, D.; Yao, Y.; Gao, R.; Ye, F.; Liu, X.; Liao, W. Genome-wide identification and expression analysis of serine hydroxymethyltransferase (SHMT) gene family in tomato (Solanum lycopersicum). PeerJ 2022, 10, e12943. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Ma, S.; Wang, M.; Wu, J.; Guo, W.; Chen, Y.; Li, G.; Wang, Y.; Shi, W.; Xia, G.; Fu, D.; et al. WheatOmics: A platform combining multiple omics data to accelerate functional genomics studies in wheat. Mol. Plant 2021, 14, 1965–1968. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Liu, H.; Yang, Y.; Zhang, L. Zinc finger-Homeodomain transcriptional factors (ZF-HDs) in wheat (Triticum aestivum L.): Identification, evolution, expression analysis and response to abiotic stresses. Plants 2021, 10, 593. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ohyanagi, H.; Tanaka, T.; Sakai, H.; Shigemoto, Y.; Yamaguchi, K.; Habara, T.; Fujii, Y.; Antonio, B.A.; Nagamura, Y.; Imanishi, T.; et al. The Rice Annotation Project Database (RAP-DB): Hub for Oryza sativa ssp. Japonica genome information. Nucleic Acids Res. 2006, 34, D741–D744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawton-Rauh, A. Evolutionary dynamics of duplicated genes in plants. Mol. Phylogenet. Evol. 2003, 29, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Gardestroem, P.; Henricson, D.; Ericson, I.U.U.S.; Edwards, G.E. The localization of serine hydroxymethyltransferase in leaves of C3 and C4 species. Physiol. Plant. 1985, 64, 29–33. [Google Scholar] [CrossRef]
- Khalil, S.R.M.; Ashoub, A.; Hussein, B.A.; Brüggemann, W.; Hussein, E.H.A.; Tawfik, M.S. Physiological and molecular evaluation of ten Egyptian barley cultivars under salt stress conditions. J. Crop Sci. Biotechnol. 2022, 25, 91–101. [Google Scholar] [CrossRef]
- Khalil, S.R.M.; Ashoub, A.; Hussein, B.A.; Brüggemann, W.; Hussein, E.H.A.; Tawfik, M.S. Physiological and molecular studies on wild barley (hordeum spontaneum) under salt stress. Plant Arch. 2020, 20, 9669–9681. [Google Scholar]
- Nambiar, D.; Berhane, T.K.; Shew, R.; Schwarz, B.; Duff, M.J.; Howell, E.E. In vivo titration of folate pathway enzymes. Appl. Environ. Microbiol. 2018, 84, e01139-18. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, R.; Gao, J.; Song, G.; Li, J.; Li, W.; Qi, Y.; Li, Y.; Li, G. CRISPR/Cas9-mediated genome editing for wheat grain quality improvement. Plant Biotechnol. J. 2021, 19, 1684–1686. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Transcript Name | Transcript ID | Genomic Position | Exon Number | ORF (bp) | Protein Length (aa) | Molecular Weight (kDa) | Isoelectric Point (pI) | GRAVY | Subcellular Localization | Group |
|---|---|---|---|---|---|---|---|---|---|---|---|
| TaSHMT1 | TaSHMT1.1 | TraesCS1A02G218700.1 | 1A:387359860:387363581:1 | 4 | 1608 | 535 | 58.24 | 8.13 | −0.22 | Mitochondrion | Ia |
| TaSHMT1 | TaSHMT1.2 | TraesCS1A02G218700.2 | 1A:387359860:387363581:1 | 4 | 1605 | 534 | 58.11 | 8.32 | −0.214 | Mitochondrion | Ia |
| TaSHMT2 | TaSHMT2.1 | TraesCS1B02G232200.1 | 1B:417264786:417268575:1 | 4 | 1599 | 532 | 58.05 | 8.12 | −0.253 | Mitochondrion | Ia |
| TaSHMT2 | TaSHMT2.2 | TraesCS1B02G232200.2 | 1B:417264786:417268575:1 | 4 | 1602 | 533 | 58.18 | 7.89 | −0.259 | Mitochondrion | Ia |
| TaSHMT3 | TaSHMT3.1 | TraesCS1D02G220400.1 | 1D:308179375:308183217:1 | 4 | 1602 | 533 | 58.03 | 7.9 | −0.241 | Mitochondrion | Ia |
| TaSHMT4 | TaSHMT4.1 | TraesCS2A02G493700.1 | 2A:726022665:726026360:1 | 4 | 1416 | 471 | 51.46 | 7.27 | −0.206 | Mitochondrion | Ia |
| TaSHMT5 | TaSHMT5.1 | TraesCS2B02G521700.1 | 2B:716779117:716782669:1 | 4 | 1416 | 471 | 51.45 | 7.27 | −0.191 | Mitochondrion | Ia |
| TaSHMT6 | TaSHMT6.1 | TraesCS2D02G493600.1 | 2D:591019752:591023458:1 | 4 | 1416 | 471 | 51.46 | 7.27 | −0.206 | Mitochondrion | Ia |
| TaSHMT7 | TaSHMT7.1 | TraesCS3A02G385600.1 | 3A:635148224:635152237:1 | 4 | 1758 | 585 | 63.64 | 7.3 | −0.379 | Mitochondrion | Ib |
| TaSHMT8 | TaSHMT8.1 | TraesCS3B02G417800.1 | 3B:654648169:654651767:1 | 5 | 1548 | 515 | 56.62 | 8.22 | −0.335 | Mitochondrion | Ib |
| TaSHMT9 | TaSHMT9.1 | TraesCS3D02G378700.1 | 3D:495915950:495920036:1 | 4 | 1752 | 583 | 63.37 | 7.17 | −0.362 | Mitochondrion | Ib |
| TaSHMT10 | TaSHMT10.1 | TraesCS4A02G246100.1 | 4A:556751829:556756496:−1 | 15 | 1533 | 510 | 56.13 | 8.5 | −0.275 | Mitochondrion | IIb |
| TaSHMT11 | TaSHMT11.1 | TraesCS4B02G069300.1 | 4B:62967286:62972392:1 | 15 | 1533 | 510 | 56.13 | 8.5 | −0.275 | Mitochondrion | IIb |
| TaSHMT12 | TaSHMT12.1 | TraesCS4D02G068100.1 | 4D:43300118:43305497:1 | 15 | 1533 | 510 | 56.13 | 8.5 | −0.275 | Mitochondrion | IIb |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Li, N.; Zhao, Y.; Kang, G.-Z.; Zhao, Y.-H.; Xu, H.-W. Serine Hydroxymethyltransferase (SHMT) Gene Family in Wheat (Triticum aestivum L.): Identification, Evolution, and Expression Analysis. Agronomy 2022, 12, 1346. https://doi.org/10.3390/agronomy12061346
Liu H, Li N, Zhao Y, Kang G-Z, Zhao Y-H, Xu H-W. Serine Hydroxymethyltransferase (SHMT) Gene Family in Wheat (Triticum aestivum L.): Identification, Evolution, and Expression Analysis. Agronomy. 2022; 12(6):1346. https://doi.org/10.3390/agronomy12061346
Chicago/Turabian StyleLiu, Hao, Na Li, Yuan Zhao, Guo-Zhang Kang, Yan-Hong Zhao, and Hua-Wei Xu. 2022. "Serine Hydroxymethyltransferase (SHMT) Gene Family in Wheat (Triticum aestivum L.): Identification, Evolution, and Expression Analysis" Agronomy 12, no. 6: 1346. https://doi.org/10.3390/agronomy12061346