
Effect of the Combined Treatments with LC2017 and Trichoderma atroviride Strain I-1237 on Disease Development and Defense Responses in Vines Infected by Lasiodiplodia theobromae

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Inoculum Preparation
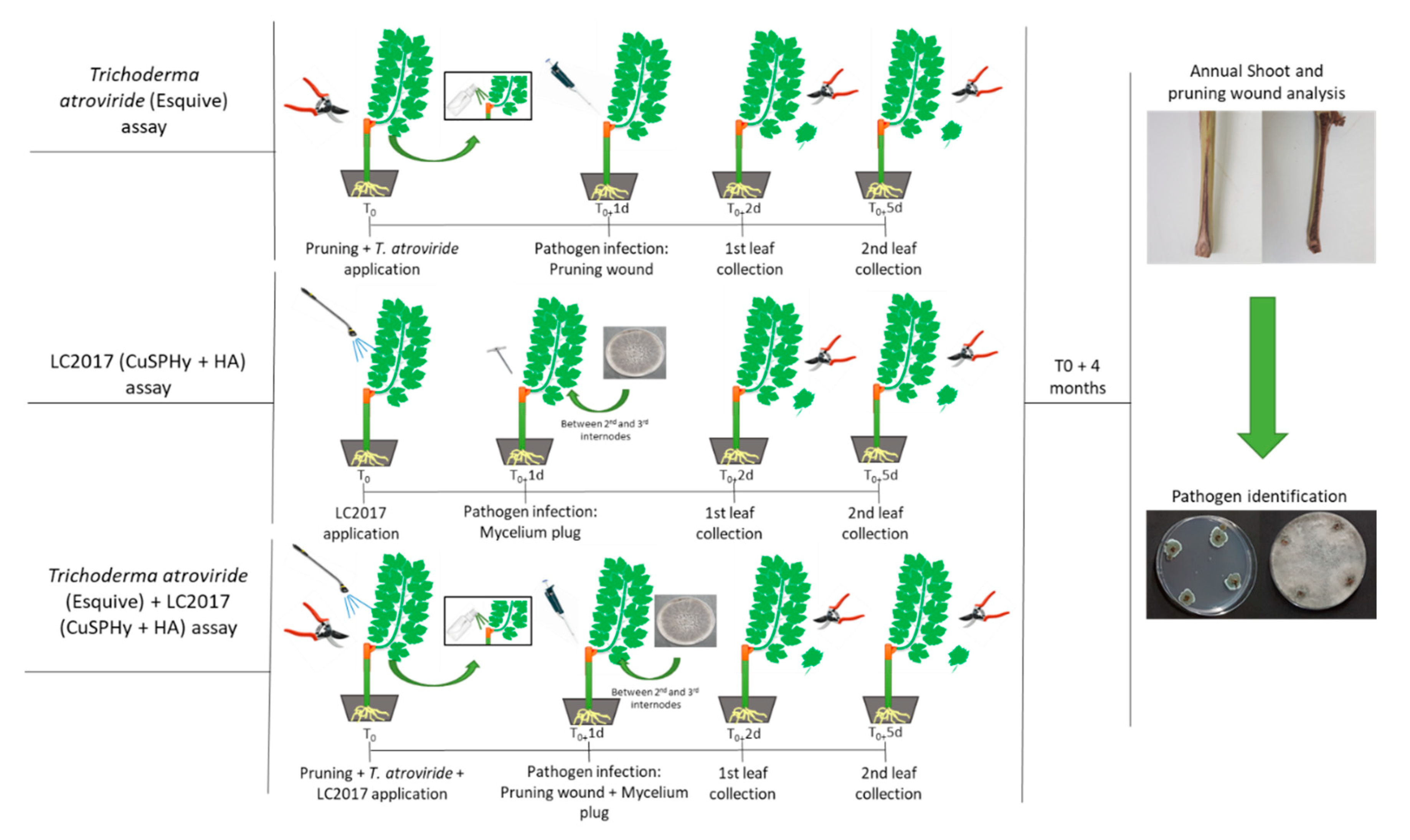
2.2. Plant Material, Experimental Design and Treatments
2.2.1. Plant Material
2.2.2. Tested Products
2.2.3. Product Application and Pathogen Inoculation
2.3. Gene Expression Analysis
2.4. Determination of Internal Lesions and Pathogen Recovery
2.5. Statistical Analysis
3. Results
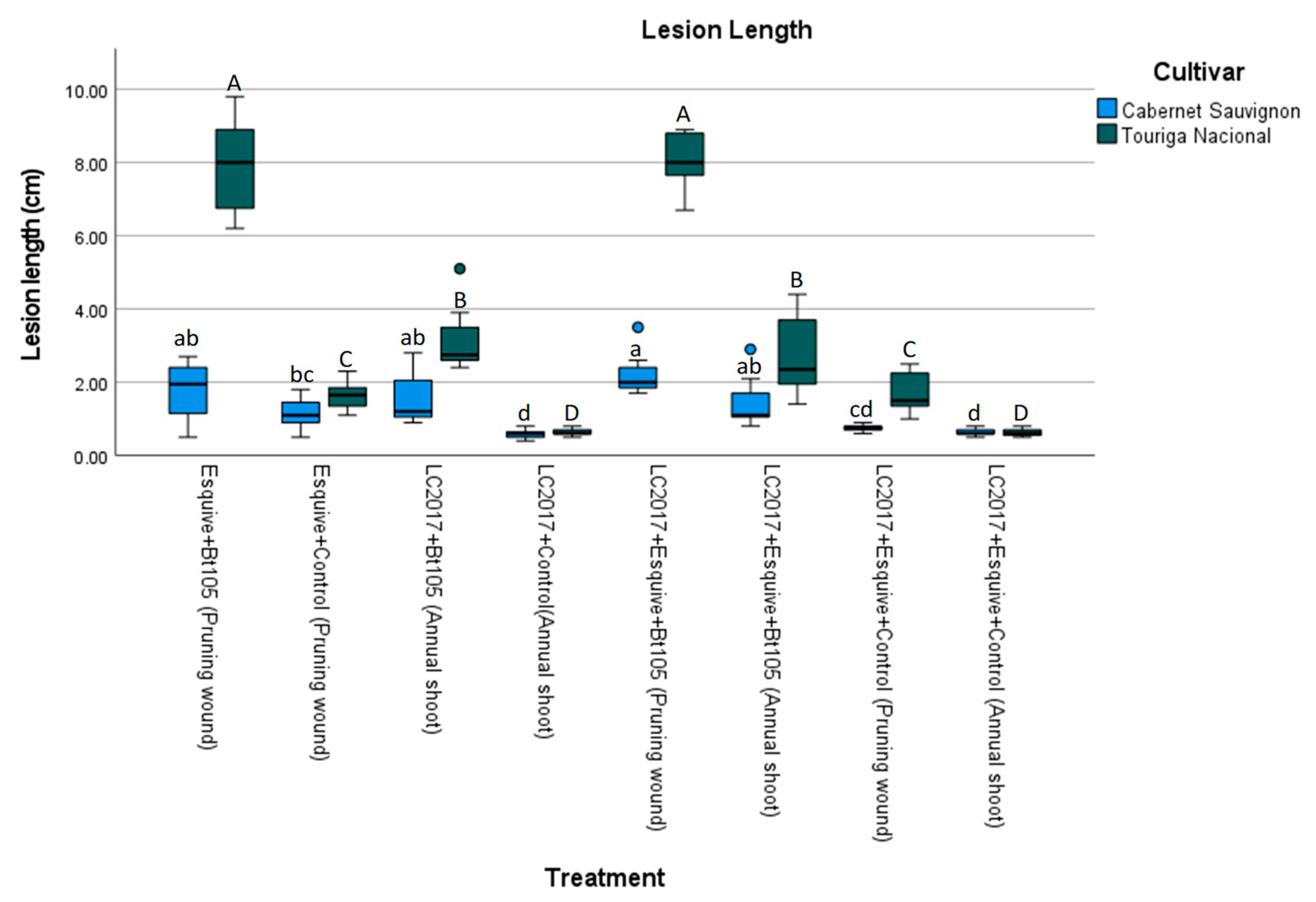
3.1. Relationship between the Lesion Length Induced by L. theobromae and Treatments
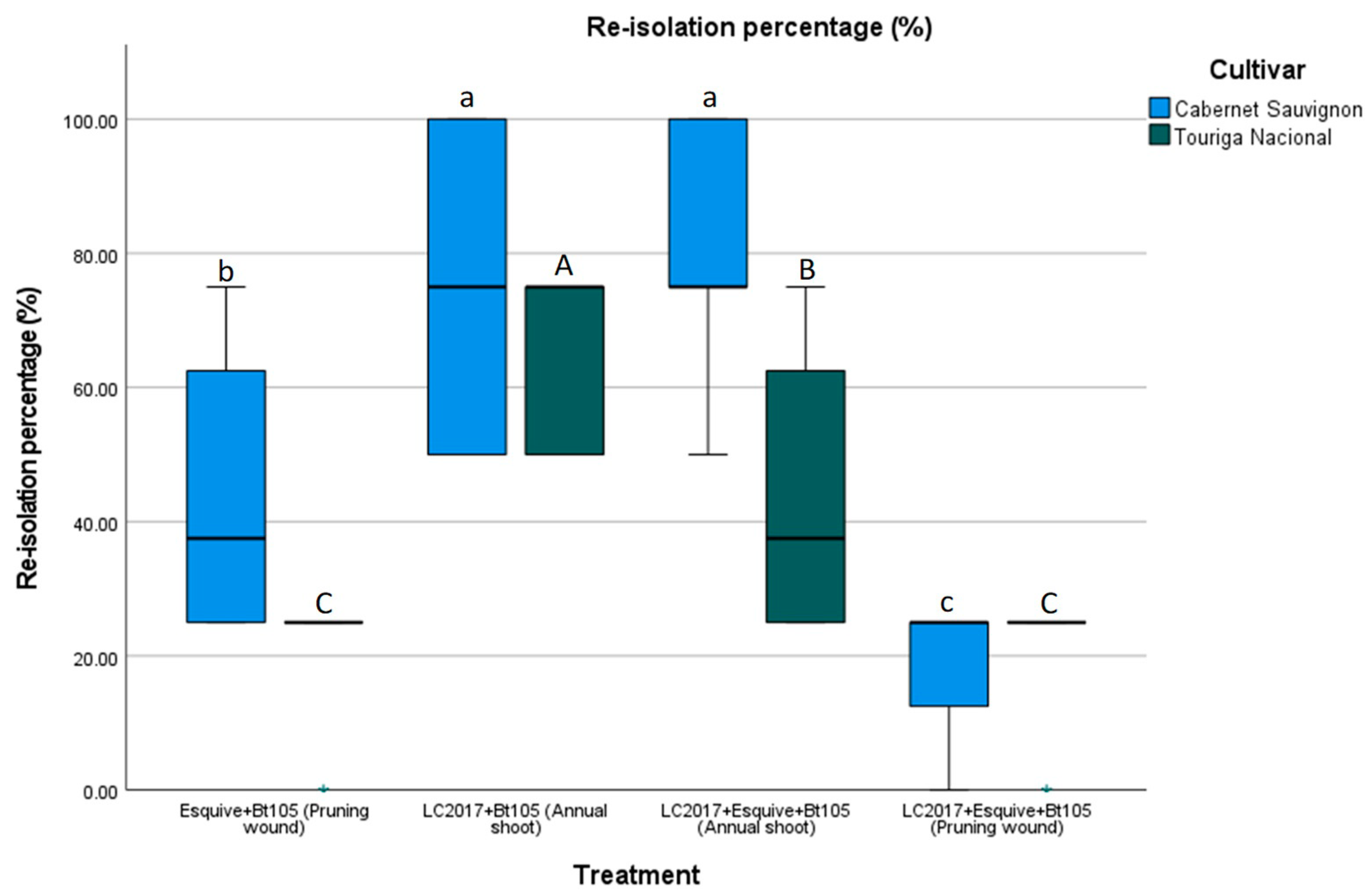
3.2. Pathogen Recovery
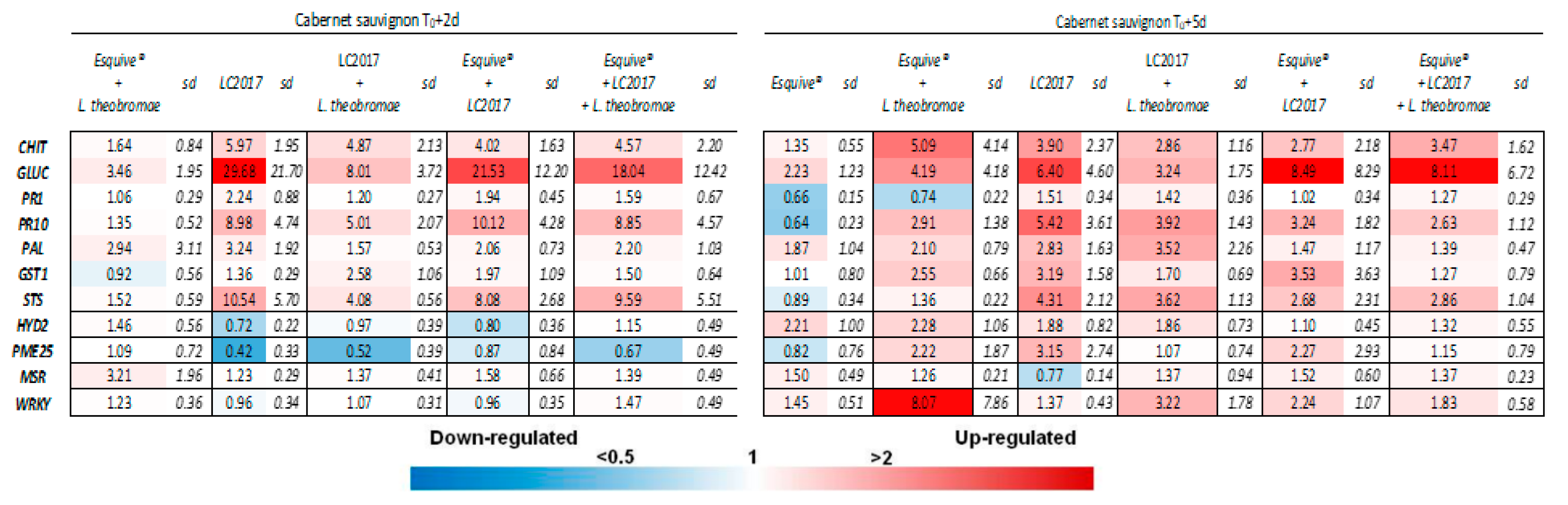
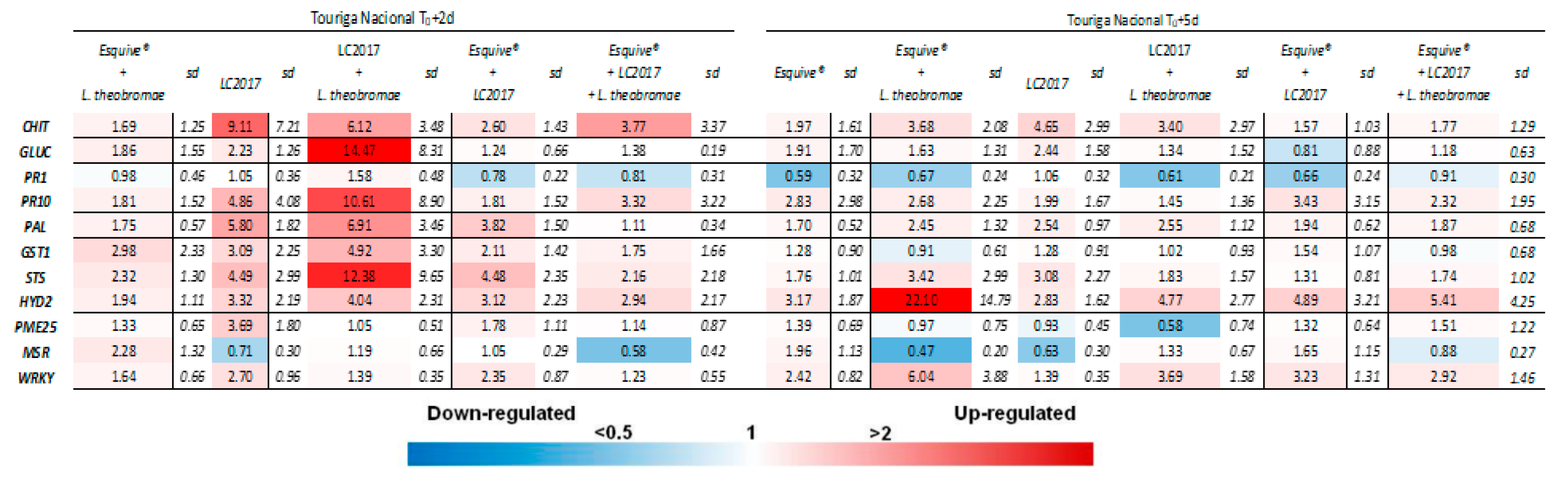
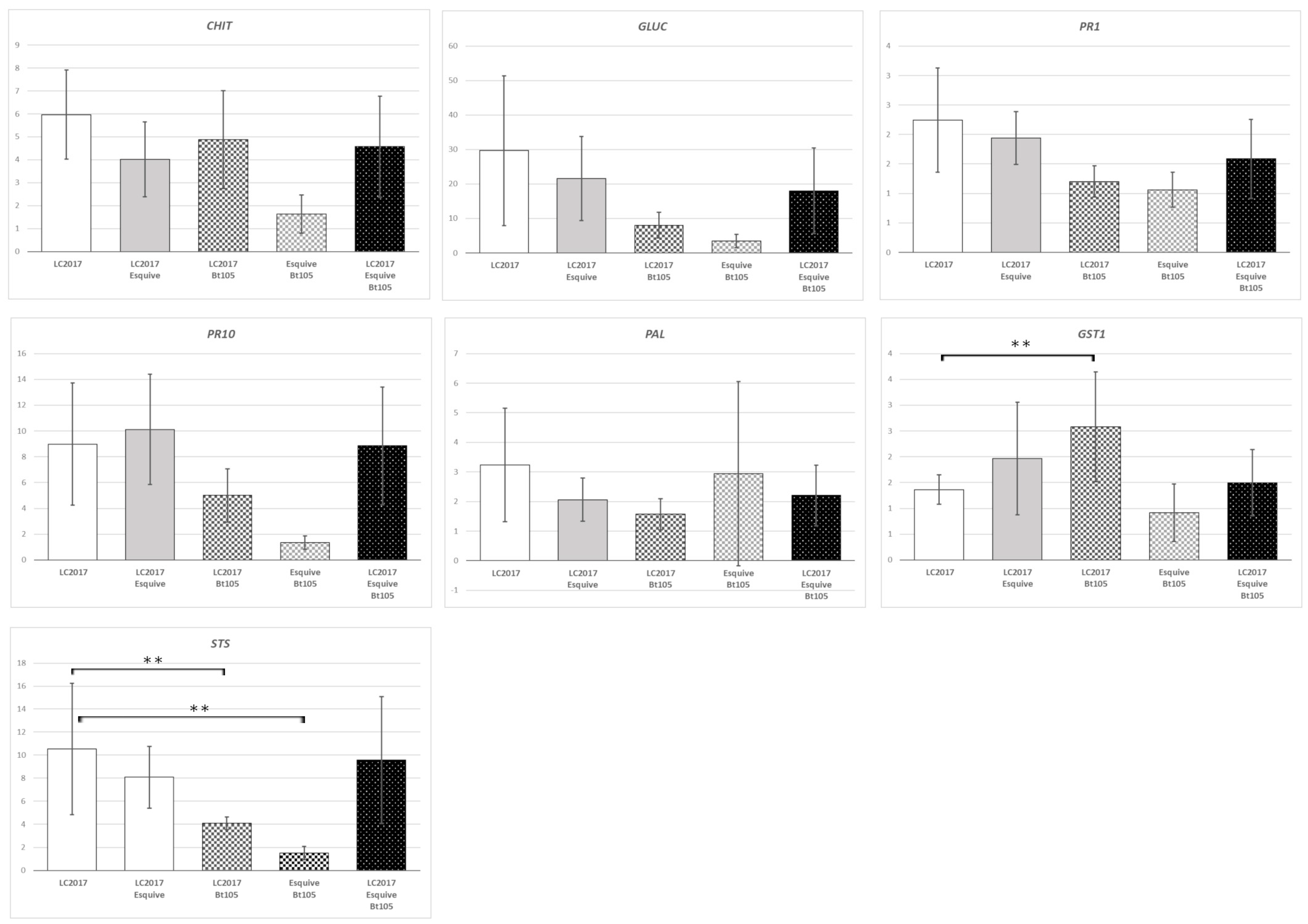
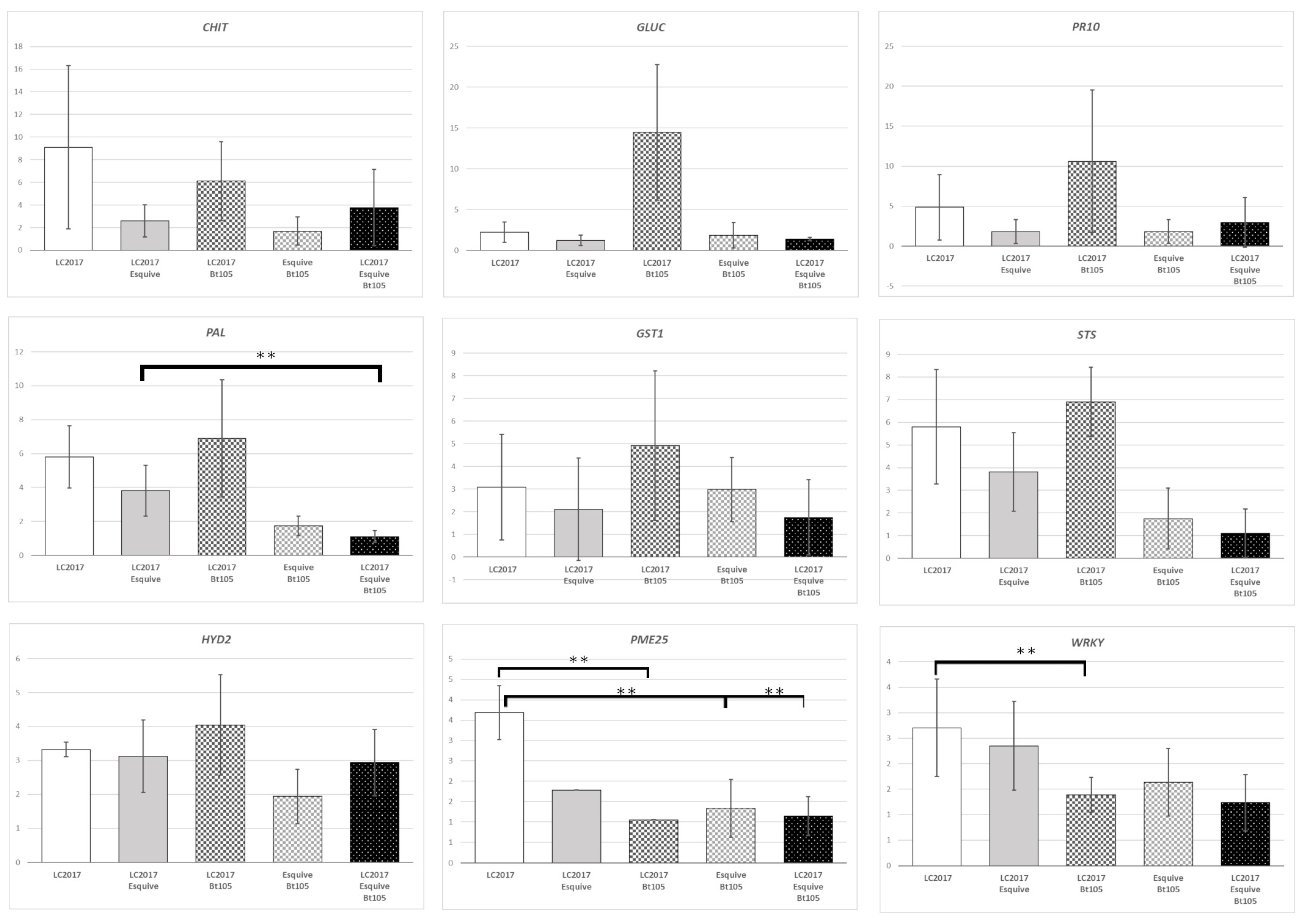
3.3. Plant Responses at Gene Expression Levels According to the Treatments
3.3.1. Effect of LC2017 Treatment Alone
3.3.2. Effect of the Double Treatment Esquive® and LC2017
3.3.3. Influence of Cultivar on Defense Responses
4. Discussion
4.1. Effect of Treatments on Lesion Length Induced by L. theobromae
4.2. The Combination of Esquive® with LC2017 Does Not Appear to Improve Plant Defense Response
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bertsch, C.; Ramírez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, C.; Fontaine, F. Grapevine trunk diseases: Complex and still poorly understood. Plant Pathol. 2013, 62, 243–265. [Google Scholar] [CrossRef] [Green Version]
- Gramaje, D.; Úrbez-Torres, J.R.; Sosnowski, M. Managing grapevine trunk diseases with respect to etiology and epidemiology: Current strategies and future prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente, M.; Fontaine, F.; Gramaje, D.; Armengol, J.; Smart, R.; Nagy, Z.A.; Rego, C.; Corio-Costet, M. Grapevine Trunk Diseases. A Review, 1st ed.; OIV Publications: Paris, France, 2016; pp. 4–7. [Google Scholar]
- Mondello, V.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food Quality Protection Act. Food Quality Protection Act of 1996, P.L. 104–170, Title II, Section 303, Enacted August 3, 1996. Codified in: Title 7, U.S. Code, Section 136r-1. Integrated Pest Management. 1998. Integrated Pest Management. Available online: https://www.epa.gov/laws-regulations/summary-food-quality-protection-act (accessed on 14 January 2022).
- Kogan, M. Integrated Pest Management: Historical Perspectives and Contemporary Developments. Annu. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Berbegal, M.; Ramón-Albalat, A.; León, M.; Armengol, J. Evaluation of long-term protection from nursery to vineyard provided by Trichoderma atroviride SC1 against fungal grapevine trunk pathogens. Pest Manag. Sci. 2020, 76, 967–977. [Google Scholar] [CrossRef]
- Leal, C.; Richet, N.; Guise, J.F.; Gramaje, D.; Armengol, J.; Fontaine, F.; Trotel-Aziz, P. Cultivar contributes to the beneficial effects of Bacillus subtilis PTA-271 and Trichoderma atroviride SC1 to protect grapevine against Neofusicoccum parvum. Front. Microbiol. 2021, 12, 726132. [Google Scholar] [CrossRef]
- Martínez-Diz, M.D.P.; Díaz-Losada, E.; Díaz-Fernández, Á.; Bouzas-Cid, Y.; Gramaje, D. Protection of grapevine pruning wounds against Phaeomoniella chlamydospora and Diplodia seriata by commercial biological and chemical methods. Crop Prot. 2021, 143, 105465. [Google Scholar] [CrossRef]
- Mutawila, C.; Fourie, P.H.; Halleen, F.; Mostert, L. Grapevine cultivar variation to pruning wound protection by Trichoderma species against trunk pathogens. Phytopathol. Mediterr. 2011, 50, S264–S276. [Google Scholar]
- Mutawila, C.; Halleen, F.; Mostert, L. Development of benzimidazole resistant Trichoderma strains for the integration of chemical and biocontrol methods of grapevine pruning wound protection. BioControl 2015, 60, 387–399. [Google Scholar] [CrossRef]
- Mutawila, C.; Halleen, F.; Mostert, L. Optimisation of time of application of Trichoderma biocontrol agents for protection of grapevine pruning wounds. Aust. J. Grape Wine Res. 2016, 22, 279–287. [Google Scholar] [CrossRef]
- Mounier, E.; Boulisset, F.; Cortes, F.; Cadiou, M. Esquive® WP limits development of grapevine trunk diseases and safeguards the production potential of vineyards. In Biocontrol of Major Grapevine Diseases. Leading Research; Compant, S., Mathieu, F., Eds.; CABI: Wallingford, UK, 2016; pp. 160–170. [Google Scholar]
- Mounier, E.; Cortes, F.; Cadious, M.; Pajot, E. The benefits of Trichoderma atroviride strain I-1237 for the protection of grapevines against trunk diseases: From the nursery to the vineyard. Phytopathol. Mediterr. 2014, 53, 591–592. [Google Scholar]
- Reis, P.; Letousey, P.; Rego, C. Trichoderma atroviride strain I-1237 protects pruning wounds against grapevine wood pathogens. Phytopathol. Mediterr. 2017, 56, 580. [Google Scholar]
- Reis, P.; Gaspar, A.; Alves, A.; Fontaine, F.; Rego, C. Combining an HA + Cu (II) Site-Targeted Copper-Based Product with a Pruning Wound Protection Program to Prevent Infection with Lasiodiplodia spp. in Grapevine. Plants 2021, 10, 2376. [Google Scholar] [CrossRef] [PubMed]
- Úrbez-Torres, J.R.; Tomaselli, E.; Pollard-Flamand, J.; Boulé, J.; Gerin, D.; Pollastro, S. Characterization of Trichoderma isolates from southern Italy, and their potential biocontrol activity against grapevine trunk disease fungi. Phytopathol. Mediterr. 2020, 59, 425–439. [Google Scholar]
- Harman, G.E. Overview of mechanisms and uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieterse, J.; Zamioudis, C.; Berendsen, L.; Weller, M.; Van Wees, M.; Bakker, M. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Van Wees, S.C.; Van der Ent, S.; Pieterse, C.M. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Barbetti, M.J.; Li, H.; Lorito, M. A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol. Mol. Plant Pathol. 2008, 72, 80–86. [Google Scholar] [CrossRef]
- Waghunde, R.R.; Shelake, R.M.; Sabalpara, A.N. Trichoderma: A significant fungus for agriculture and environment. Afr. J. Agric. Res. 2016, 11, 1952–1965. [Google Scholar]
- De la Fuente, M.; Fernandez-Calvino, D.; Tylkowski, B.; Montornes, J.M.; Olkiewics, M.; Pereira, R.; Cachada, A.; Caffi, T.; Fedele, G.; De Herralde, F. Alternatives to Cu applications in viticulture. How R&D projects can provide applied solutions, helping to establish legislation limits. In Grapes and Wine; Morata, A., Loira, I., González, C., Eds.; IntechOpen: London, UK, 2021. [Google Scholar]
- Perez-Rodriguez, P.; Soto-Gómez, D.; De La Calle, I.; López-Periago, J.E.; Paradelo, M. Rainfall-induced removal of copper-based spray residues from vines. Ecotoxicol. Environ. Saf. 2016, 132, 304–310. [Google Scholar] [CrossRef]
- Available online: https://ec.europa.eu/food/plant/pesticides/eu-pesticides-database/active-substances/?event=as.details&as_id=1221 (accessed on 4 April 2022).
- Berkelmann-Löhnertz, B.; Klärner, S.; Baus, O.; Herrmann, G.; Flemming, B.; Keicher, R.; Schwarz, H.P.; Pfliehinger, M.; Löhnertz, O. Kupferreduzierung und Kupfersubstitution im Weinbau–zum Stand der Dinge aus Sicht der Forschung. In Reports from the Julius Kühn-Institut–Plant Protection in Organic Farming, Proceedings of the 11th Workshop in Berlin, Germany, 1 December 2011; Berichte aus dem Julius Kühn-Institut: Berlin, Germany, 2012; Volume 164, pp. 70–95. [Google Scholar]
- Battiston, E.; Salvatici, M.C.; Lavacchi, A.; Gatti, A.; Di Marco, S.; Mugnai, L. Functionalisation of a nano-structured hydroxyapatite with copper (II) compounds as pesticide: In situ TEM and ESEM observations of treated Vitis vinifera L. leaves. Pest Manag. Sci. 2018, 74, 1903–1915. [Google Scholar] [CrossRef] [PubMed]
- Battiston, E.; Compant, S.; Antonelli, L.; Mondello, V.; Clément, C.; Simoni, A.; Di Marco, S.; Mugnai, L.; Fontaine, F. In planta activity of novel copper(II) based formulations to inhibit the Esca-associated fungus Phaeoacremonium minimum in grapevine propagation material. Front. Plant Sci. 2021, 12, 312. [Google Scholar] [CrossRef] [PubMed]
- Mondello, V.; Fernandez, O.; Guise, J.-F.; Trotel-Aziz, P.; Fontaine, F. In planta Activity of the Novel Copper Product HA + Cu(II) Based on a Biocompatible Drug Delivery System on Vine Physiology and Trials for the Control of Botryosphaeria Dieback. Front. Plant Sci. 2021, 12, 693995. [Google Scholar] [CrossRef]
- Mondello, V.; Lemaître-Guillier, C.; Trotel-Aziz, P.; Gougeon, R.; Acedo, A.; Schmitt-Kopplin, P.; Adrian, M.; Pinto, C.; Fernandez, O.; Fontaine, F. Assessment of a new copper-based formulation to control esca disease in field and study on its impact on the vine microbiome, vine physiology and enological parameters of the juice. J. Fungi 2022, 8, 151. [Google Scholar] [CrossRef] [PubMed]
- Úrbez-Torres, J.R. The status of Botryosphaeriaceae species infecting grapevines. Phytopathol. Mediterr. 2011, 50, S5–S45. [Google Scholar]
- Reis, P.; Gaspar, A.; Alves, A.; Fontaine, F.; Rego, C. Response of different grapevine cultivars to infection by Lasiodiplodia theobromae and Lasiodiplodia mediterranea. Plant Dis. 2022. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Slippers, M.J.; Rheeder, J.; Marasas, W.F.O.; Phillips, A.J.L.; Alves, A.; Burguess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.J.L. Botryosphaeria species associated with diseases of grapevines in Portugal. Phytopathol. Mediterr. 2002, 41, 3–18. [Google Scholar]
- Úrbez-Torres, J.R.; Leavitt, G.M.; Voegel, T.; Gubler, W.D. Identification and distribution of Botryosphaeria species associated with grapevine cankers in California. Plant Dis. 2006, 90, 1490–1503. [Google Scholar] [CrossRef] [Green Version]
- Reis, P.; Pierron, R.; Larignon, P.; Lecomte, P.; Abou-Mansour, E.; Farine, S.; Bertsch, C.; Jacques, A.; Trotel-Aziz, P.; Rego, C.; et al. Vitis methods to understand and develop strategies for diagnosis and sustainable control of grapevine trunk diseases. Phytopathology 2019, 109, 916–931. [Google Scholar] [CrossRef]
- Magnin-Robert, M.; Spagnolo, A.; Dilezitoko, A.; Cilindre, C.; Mercier, L.; Schaeffer-Reiss, C.; van Dorsselaer, A.; Clément, C.; Fontaine, F. Proteomic insights into changes in grapevine wood in response to Esca proper and apoplexy. Phytopathol. Mediterr. 2014, 53, 168–187. [Google Scholar]
- Spagnolo, A.; Mondello, V.; Larignon, P.; Villaume, S.; Rabenoelina, F.; Clément, C.; Fontaine, F. Defense responses in grapevine (cv. Mourvèdre) after Inoculation with the Botryosphaeria dieback pathogens Neofusicoccum parvum and Diplodia seriata and their relationship with flowering. Int. J. Mol. Sci. 2017, 18, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spagnolo, A.; Larignon, P.; Magnin-Robert, M.; Hovasse, A.; Cilindre, C.; Van Dorsselaer, A.; Clément, C.; Schaeffer-Reiss, C.; Fontaine, F. Flowering as the most highly sensitive period of grapevine (Vitis vinifera L. cv Mourvèdre) to the Botryosphaeria dieback agents Neofusicoccum parvum and Diplodia seriata infection. Int. J. Mol. Sci. 2014, 15, 9644–9669. [Google Scholar] [CrossRef] [PubMed]
- Magnin-Robert, M.; Letousey, P.; Spagnolo, A.; Rabenoelina, F.; Jacquens, L.; Mercier, L.; Clément, C.; Fontaine, F. Leaf stripe of esca induces alteration of photosynthesis and defence reactions in presymptomatic leaves. Funct. Plant Biol. 2011, 38, 856–866. [Google Scholar] [CrossRef]
- Magnin-Robert, M.; Spagnolo, A.; Boulanger, A.; Joyeux, C.; Clément, C.; Abou-Mansour, E.; Fontaine, F. Changes in plant metabolism and accumulation of fungal metabolites in response to Esca proper and apoplexy expression in the whole grapevine. Phytopathology 2016, 106, 541–553. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Abou-Mansour, E.; Courteaux, B.; Rabenoelina, F.; Clément, C.; Fontaine, F.; Aziz, A. Bacillus subtilis PTA-271 Counteracts Botryosphaeria Dieback in Grapevine, Triggering Immune Responses and Detoxification of Fungal Phytotoxins. Front. Plant Sci. 2019, 10, 25. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Gubler, W.D. Pathogenicity of Botryosphaeriaceae Species Isolated from Grapevine Cankers in California. Plant Dis. 2009, 93, 584–592. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and roles of stilbenes in plants. Plant Sci. 2009, 177, 143–155. [Google Scholar] [CrossRef]
- Stempien, E.; Jean, R.; Pierron, G.; Adendorff, I.; Van Jaarsveld, W.J.; Halleen, F.; Mostert, L. Host defence activation and root colonization of grapevine rootstocks by the biological control fungus Trichoderma atroviride. Phytopathol. Mediterr. 2020, 59, 615–626. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Product | Inoculation |
|---|---|---|
| 1 | LC2017 | Water (control) |
| 2 | LC2017 | L. theobromae (Bt105) |
| 3 | Esquive® | Water (control) |
| 4 | Esquive® | L. theobromae (Bt105) |
| 5 | LC2017 + Esquive® | Water (control) |
| 6 | LC2017 + Esquive® | L. theobromae (Bt105) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reis, P.; Mondello, V.; Diniz, I.; Alves, A.; Rego, C.; Fontaine, F. Effect of the Combined Treatments with LC2017 and Trichoderma atroviride Strain I-1237 on Disease Development and Defense Responses in Vines Infected by Lasiodiplodia theobromae. Agronomy 2022, 12, 996. https://doi.org/10.3390/agronomy12050996
Reis P, Mondello V, Diniz I, Alves A, Rego C, Fontaine F. Effect of the Combined Treatments with LC2017 and Trichoderma atroviride Strain I-1237 on Disease Development and Defense Responses in Vines Infected by Lasiodiplodia theobromae. Agronomy. 2022; 12(5):996. https://doi.org/10.3390/agronomy12050996
Chicago/Turabian StyleReis, Pedro, Vincenzo Mondello, Inês Diniz, Artur Alves, Cecília Rego, and Florence Fontaine. 2022. "Effect of the Combined Treatments with LC2017 and Trichoderma atroviride Strain I-1237 on Disease Development and Defense Responses in Vines Infected by Lasiodiplodia theobromae" Agronomy 12, no. 5: 996. https://doi.org/10.3390/agronomy12050996