
Fermentation Quality and Bacterial Diversity of Broussonetia papyrifera Leaves Ensiled with Lactobacillus plantarum and Stored at Different Temperatures

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Silage Preparation
2.2. Gas Production Analysis
2.3. Analysis of Microbial Counting and Chemical Composition
2.4. Bacterial Community Analysis
2.5. Nutritional Component Analysis
2.6. Statistical Analysis
3. Results
3.1. Characteristics of Raw B. papyrifera Leaves before Ensiling
3.2. Fermentation Quality and Microbial Populations of B. papyrifera Leaves Silage
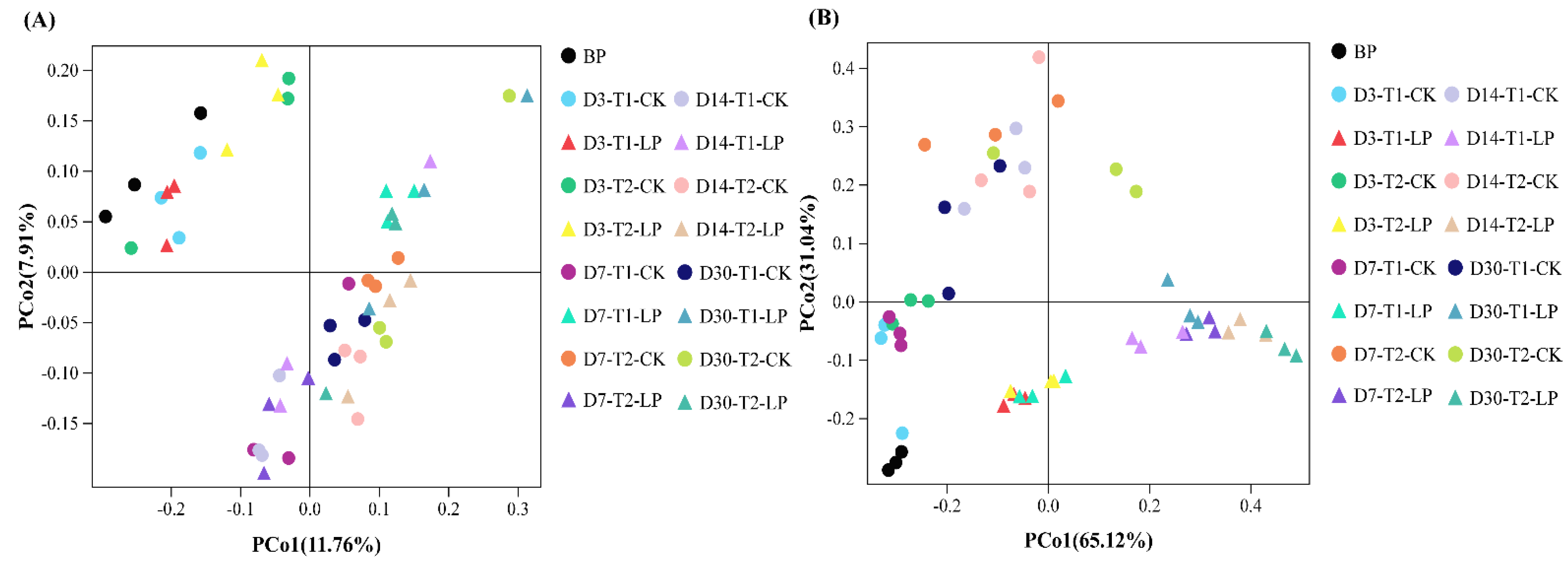
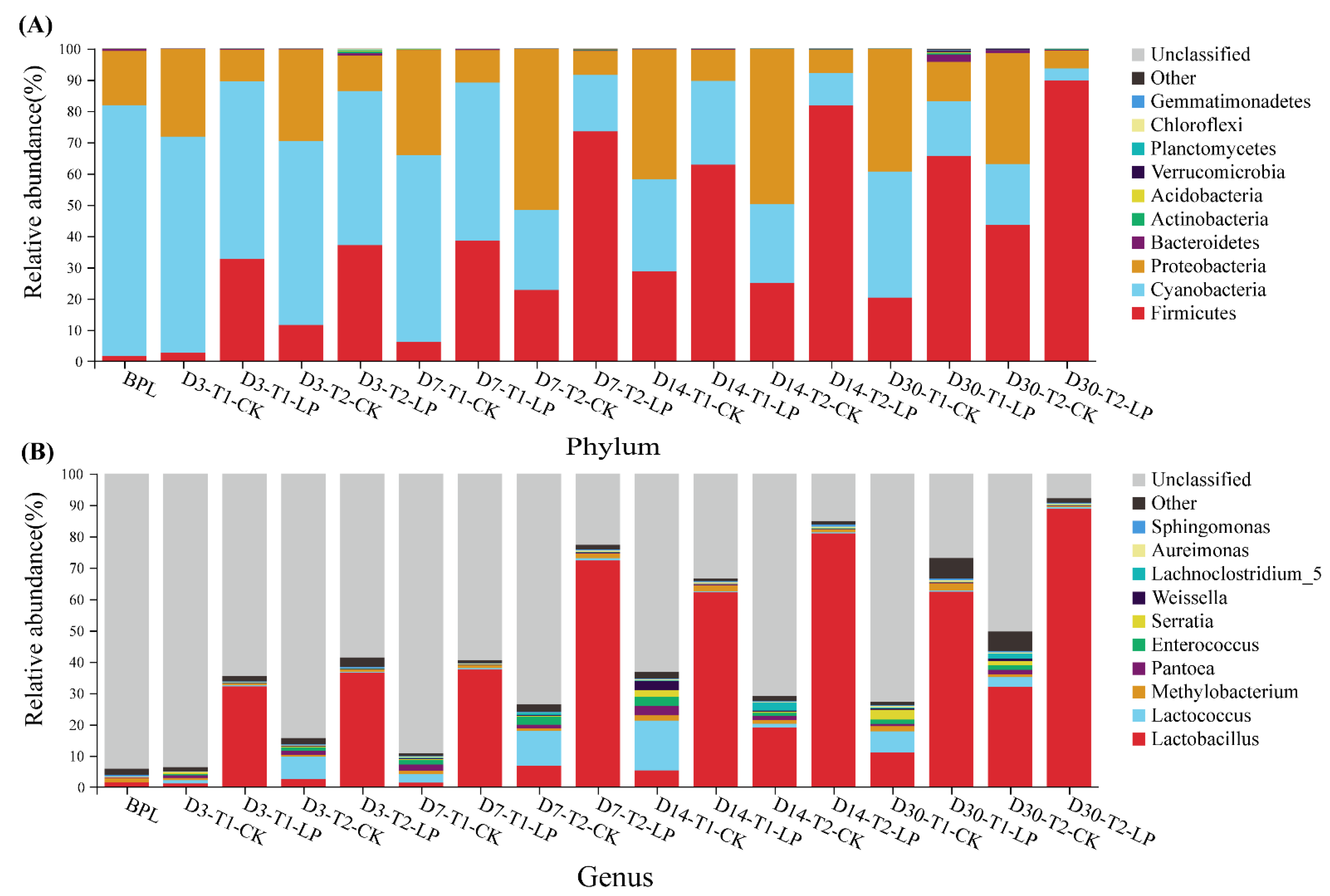
3.3. Bacterial Community of B. papyrifera Leaves Silage
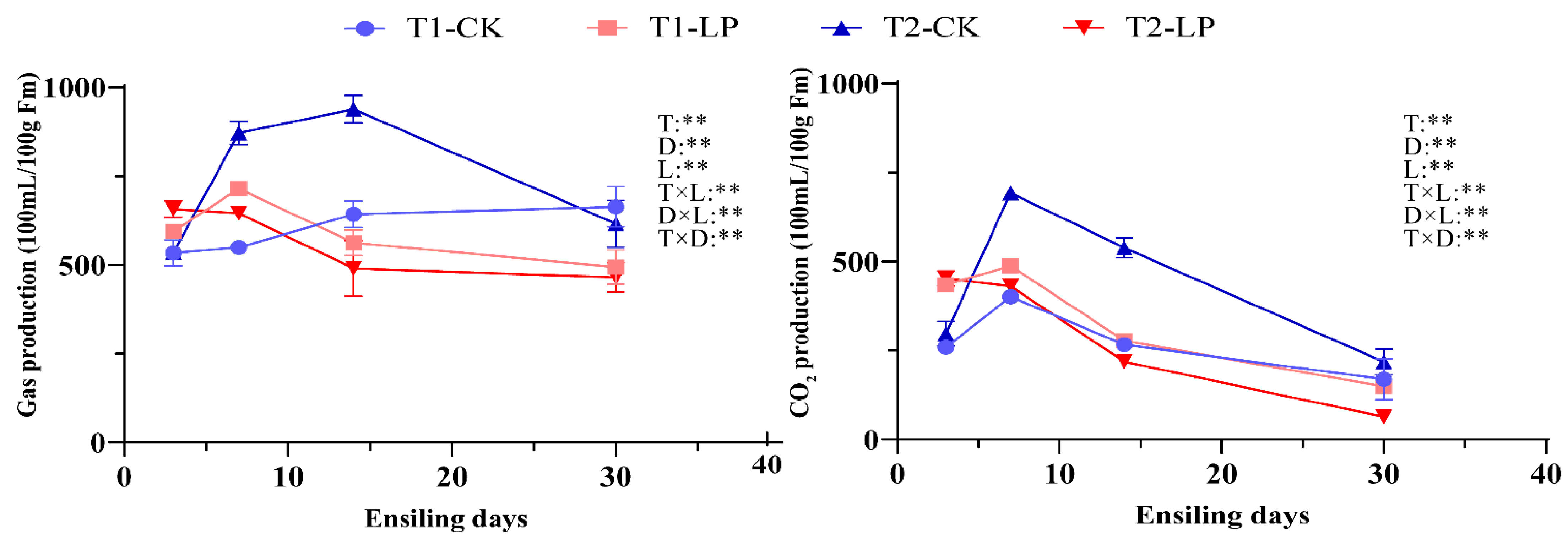
3.4. Gas Production of B. papyrifera Leaves Silage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, D.; Bhat, K.P.; Fong, H.H.; Farnsworth, N.R.; Pezzuto, J.M.; Kinghorn, A.D. Aromatase inhibitors from Broussonetia papyrifera. J. Nat. Prod. 2001, 64, 1286–1293. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Yang, H.; Meng, X.; Yoo, C.G.; Li, M.; Pu, Y.; Ragauskas, A.J.; Sykes, R.W. Adsorption of cellobiohydrolases I onto lignin fractions from dilute acid pretreated Broussonetia papyrifera. Bioresour. Technol. 2017, 244, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Chen, H.; Wu, C.; Liao, P.; Lin, Y.; Hsieh, M.; Ching, H. Comparison with various parts of Broussonetia papyrifera as to the antinociceptive and anti-inflammatory activities in rodents. J. Agric. Chem. Soc. Jpn. 2008, 72, 2377–2384. [Google Scholar]
- Dong, L.; Zhang, H.; Gao, Y.; Diao, Q. Dynamic profiles of fermentation characteristics and bacterial community composition of Broussonetia papyrifera ensiled with perennial ryegrass. Bioresour. Technol. 2020, 310, 123396. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Xu, T.; Shen, Q.; Liu, Y.; Huang, G.; Rao, D.; Song, C.; Wang, J. Productive and metabolic increments of the inclusion of Broussonetia papyrifera to replace maize silage in growing goats. Czech J. Anim. Sci. 2020, 65, 303–310. [Google Scholar] [CrossRef]
- Peng, X.; Liu, H.; Chen, P.; Tang, F.; Hu, Y.; Wang, F.; Pi, Z.; Zhao, M.; Chen, N.; Chen, H.; et al. A chromosome-scale genome assembly of paper mulberry (Broussonetia papyrifera) provides new insights into its forage and papermaking usage. Mol. Plant 2019, 12, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Z.; Yang, P.L.; Gao, X.H.; Wen, Z.G.; Dai, S.F.; Zhu, M.X.; Wang, L.X. Effects of replacement of alfalfa by big-leaf mulberry on growth performance, digestion and meat quality in growing rabbits. World Rabbit Sci. 2019, 27, 199–205. [Google Scholar] [CrossRef]
- Obour, R.; Oppong, S.K. Forage palatability of Broussonetia papyrifera an invasive species in Ghana: Relative preference and palatability by sheep and goats. J. Energy Nat. Resour. Manag. 2015, 2, 63–70. [Google Scholar] [CrossRef]
- Du, Z.; Sun, L.; Chen, C.; Lin, J.; Yang, F.; Cai, Y. Exploring microbial community structure and metabolic gene clusters during silage fermentation of paper mulberry, a high-protein woody plant. Anim. Feed Sci. Technol. 2020, 275, 114766. [Google Scholar] [CrossRef]
- Mcdonald, P. The Biochemistry of Silage; John Wiley & Sons Ltd.: Chichester, UK, 1991. [Google Scholar]
- Muck, R.E. Recent advances in silage microbiology. Agric. Food Sci. 2013, 22, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Wang, Y.; Yang, F.; Wang, Y.; Zhang, H. Screening a Lactobacillus plantarum strain for good adaption in alfalfa ensiling and demonstrating its improvement of alfalfa silage quality. J. Appl. Microbiol. 2020, 129, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Li, X.; Guan, H.; Huang, L.; Ma, X.; Peng, Y.; Li, Z.; Nie, G.; Zhou, J.; Yang, W.; et al. Microbial community and fermentation characteristic of Italian ryegrass silage prepared with corn stover and lactic acid bacteria. Bioresour. Technol. 2019, 279, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, X.; Zhao, M.; Yu, Z. Lactic acid bacteria strains for enhancing the fermentation quality and aerobic stability of Leymus chinensis silage. Grass Forage Sci. 2016, 71, 472–481. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, Z.; Wang, X.; Tian, J. Effects of inoculants and environmental temperature on fermentation quality and bacterial diversity of alfalfa silage. Anim. Sci. J. 2018, 89, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Olstorpe, M.; Lyberg, K.; Lindberg, J.E.; Schnurer, J.; Passoth, V. Population diversity of yeasts and lactic acid bacteria in pig feed fermented with whey, wet wheat distillers’ grains, or water at different temperatures. Appl. Environ. Microbiol. 2008, 74, 1696–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, Z.G.; Szakacs, G.; Ashbell, G.; Hen, Y. The effect of temperature on the ensiling process of corn and wheat. J. Appl. Microbiol. 2010, 90, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.C.; Adesogan, A.T. Influence of ensiling temperature, simulated rainfall, and delayed sealing on fermentation characteristics and aerobic stability of corn silage. J. Dairy Sci. 2006, 89, 3122–3132. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.; Zhou, W.; Yang, F.Y.; Chen, X.Y.; Zhang, Q. Effects of wilting and Lactobacillus plantarum Addition on the fermentation quality and microbial community of Moringa oleifera Leaf Silage. Front. Microbiol. 2018, 9, 1817. [Google Scholar] [CrossRef] [Green Version]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- He, L.; Zhou, W.; Wang, Y.; Wang, C.; Chen, X.; Zhang, Q. Improving the quality of rice straw silage with Moringa oleifera leaves and propionic acid: Fermentation, nutrition, aerobic stability and microbial communities. Bioresour. Technol. 2020, 299, 122579. [Google Scholar] [CrossRef]
- Thijs, S.; Op, D.B.M.; Beckers, B.; Truyens, S.; Stevens, V.; Van Hamme, J.D.; Nele, W.; Jaco, V. Comparative evaluation of four bacteria-specific primer pairs for 16s rRNA gene surveys. Front. Microbiol. 2017, 8, 494. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Lv, H.; Xing, Y.; Chen, X.; Zhang, Q. Intrinsic tannins affect ensiling characteristics and proteolysis of Neolamarckia cadamba leaf silage by largelyaltering bacterial community. Bioresour. Technol. 2020, 311, 123496. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Murphy, R.P. A method for the extraction of plant samples and the determination of total soluble carbohydrates. J. Sci. Food Agric. 1958, 9, 714–717. [Google Scholar] [CrossRef]
- Wang, C.; He, L.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Effects of mixing Neolamarckia cadamba leaves on fermentation quality, microbial community of high moisture alfalfa and stylo silage. Microb. Biotechnol. 2019, 12, 869–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyader, J.; Baron, V.; Beauchemin, K. Corn forage yield and quality for silage in short growing season areas of the Canadian prairies. Agronomy 2018, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.H. Theoretical carbohydrates requirement for alfalfa silage production. Agron. J. 1962, 54, 291–293. [Google Scholar] [CrossRef]
- Cai, Y.M.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Nakase, T. Influence of Lactobacillus spp. from an Inoculant and of Weissella and Leuconostoc spp. from forage crops on silage fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [Green Version]
- Kung, L. Potential factors that may limit the effectiveness of silage additives. In Proceedings of the 15th International Silage Conference, Madison, WI, USA, 27–29 July 2009; pp. 37–45. [Google Scholar]
- Muck, R.E. Silage microbiology and its control through additives. Rev. Bras. Zootec. 2010, 39, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Tian, J.; Zhang, Q.; Jiang, Y.; Wu, Z.; Yu, Z. Effects of mixing red clover withalfalfa at different ratios on dynamics of proteolysis and protease activities duringensiling. J. Dairy Sci. 2018, 101, 8954–8964. [Google Scholar] [CrossRef]
- Parvin, S.; Nishino, N. Bacterial community associated with ensilage process of wilted guinea grass. J. Appl. Microbiol. 2009, 107, 2029–2036. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.M.; Jiang, Y.; Pech, C.A.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157:H7 and silage additives. J. Dairy Sci. 2018, 101, 2048–2059. [Google Scholar] [CrossRef] [PubMed]
- Kung, L.J.; Taylor, C.C.; Lynch, M.P.; Neylon, J.M. The effect of treating alfalfa with Lactobacillus buchneri 40788 on silage fermentation, aerobic stability, and nutritive value for lactating dairy cows. J. Dairy Sci. 2003, 86, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Driehuis, F.S.; Elferink, S.; Wikselaar, P. Fermentation characteristics and aerobic stability of grass silage inoculated with Lactobacillus buchneri, with or without homofermentative lactic acid bacteria. Grass Forage Sci. 2010, 57, 330–343. [Google Scholar] [CrossRef]
- Pahlow, G.; Muck, R.E.; Driehuis, F.; Oude Elferink, S.J.W.H. Microbiology of Ensiling. Silage Sci. Technol. 2003, 42, 31–93. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Wang, C.; He, L.; Zhou, W.; Yang, F.; Zhang, Q. The bacterial community and fermentation quality of mulberry (Morus alba) leaf silage with or without Lactobacillus casei and sucrose. Bioresour. Technol. 2019, 293, 122059. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef]
- Muck, R.E. Inoculation of silage and its effects on silage quality. In Proceedings of the Informational Conference with Dairy and Forage Industries; Dairy Research Centre: Rosemont, IL, USA, 1996; pp. 43–51. [Google Scholar]
- Pang, D.G.; Yang, H.J.; Cao, B.B.; Wu, T.T.; Wang, J.Q. The beneficial effect of enterococcus faecium on the in vitro ruminal fermentation rate and extent of three typical total mixed rations in northern china. Livest. Sci. 2014, 167, 154–160. [Google Scholar] [CrossRef]
- Upendar, G.; Mistry, A.N.; Das, R.; Thakurata, S.G.; Chakrabarty, J.; Ghanta, K.T.C.; Dutta, S. Carbon dioxide biofixation using microorganisms and assessment of biofuel production. Environ. Prog. Sustain. Energy 2017, 37, 1594–1600. [Google Scholar] [CrossRef]
- Stefan, F.; Lukas, Y.W.; Matthias, K.; Stefan, R.K.; Michael, S.; Hans, H.R. Differences of heterotrophic 13CO2 assimilation by Pseudomonas knackmussii strain B13 and Rhodococcus opacus 1CP and potential impact on biomarker stable isotope probing. Environ. Microbiol. 2008, 10, 1641–1651. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Means ± SD |
|---|---|
| Dry Matter (g/kg) | 354 ± 1.53 |
| Crude protein (g/kg DM) | 139 ± 1.93 |
| Neutral detergent fiber (g/kg DM) | 350 ± 17.2 |
| Acid detergent fiber (g/kg DM) | 122 ± 6.83 |
| Water-soluble carbohydrates (g/kg DM) | 57.1 ± 2.47 |
| Lactic acid bacteria (log10 CFU/g FM) | 3.92 ± 0.14 |
| Coliform bacteria (log10 CFU/g FM) | 5.15 ± 0.26 |
| Yeasts (log10 CFU/g FM) | 3.78 ± 0.16 |
| Molds (log10 CFU/g FM) | <2.00 |
| Item | Days | 15 °C | 30 °C | SEM | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | LP | CK | LP | T | D | L | T × D | T × L | D × L | |||
| Ensiling Characteristics | ||||||||||||
| Dry Matter Loss (g/kg DM) | 3 | 0.80 b,B | 1.09 a,A | 0.92 a,b,B | 1.04 a | 0.041 | 0.780 | <0.01 | <0.01 | 0.676 | <0.01 | <0.01 |
| 7 | 0.76 B | 0.75 B | 0.94 B | 0.71 | ||||||||
| 14 | 1.06 a,b,A,B | 1.03 a,b,A,B | 1.21 a,A,B | 0.75 b | ||||||||
| 30 | 1.32 a,b,A | 1.00 b,c,A,B | 1.50 a,A | 0.62 c | ||||||||
| pH | 3 | 7.28 a,A | 4.83 b,A | 6.97 a,A | 5.16 b,A | 0.146 | <0.01 | <0.01 | <0.01 | 0.01 | <0.01 | <0.01 |
| 7 | 6.87 a,B | 4.69 c,A,B | 6.55 b,B | 4.56 c,B | ||||||||
| 14 | 6.47 a,C | 4.62 c,B | 5.92 bC | 4.54 d,B | ||||||||
| 30 | 6.35 a,D | 4.57 c,B | 5.50 b,D | 4.49 c,B | ||||||||
| Lactic acid (g/kg DM) | 3 | ND | ND | ND | ND | 1.29 | 0.04 | 0.152 | <0.01 | 0.56 | 0.14 | 0.50 |
| 7 | 19.6 b | 37.8 a | 29.1 a,b | 39.0 a,A,B | ||||||||
| 14 | 25.6 b | 36.8 a | 27.3 b | 37.3 a,B | ||||||||
| 30 | 27.0 b | 39.0 a | 33.9 a,b | 40.6 a,A | ||||||||
| Acetic acid (g/kg DM) | 3 | ND | 11.0 | 9.65 B | 10.6 | 0.50 | 0.17 | <0.01 | 0.69 | 0.12 | <0.01 | <0.01 |
| 7 | 9.51 | 11.5 | 10.4 B | 10.7 | ||||||||
| 14 | 10.5 | 11.5 | 12.3 B | 11.6 | ||||||||
| 30 | 13.2 b | 13.6 b | 21.3 a,A | 12.8 b | ||||||||
| Butyric acid (g/kg DM) | 3 | 1.72 b,c,D | 1.53 c,B | 2.89 a,C | 2.00 b | 1.69 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| 7 | 2.30 b,C | 1.71 c,B | 4.33 a,B | 2.08 b,c | ||||||||
| 14 | 3.22 b,B | 2.07 c,A | 4.89 a,A | 2.17 c | ||||||||
| 30 | 4.19 b,A | 2.11 c,A | 5.01 a,A | 2.16 c | ||||||||
| Ammonium nitrogen (g/kg DM) | 3 | 0.31 b,C | 0.29 b,A,B | 0.49 a,D | 0.32 b,C | 0.037 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| 7 | 0.44 b,B | 0.27 c,A | 0.78 a,C | 0.36bBC | ||||||||
| 14 | 0.51 b,B | 0.32 c,B | 0.93 a,B | 0.43 b,c,B | ||||||||
| 30 | 0.69 b,A | 0.38 c,A | 1.19 a,A | 0.63 b,A | ||||||||
| Microbial populations | ||||||||||||
| Lactic acid bacteria (log10 CFU/g FM) | 3 | 7.63 c,B | 9.73 a,A | 8.27 b,A | 9.59 a,A | 0.141 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| 7 | 8.26 d,A | 9.49 a,A | 8.52 c,A | 8.89 b,B | ||||||||
| 14 | 7.44 c,B | 9.28 a,A | 7.36 c,B | 8.27 b,C | ||||||||
| 30 | 8.58 a,A | 8.44 a,B | 7.23 b,B | 6.36 b,D | ||||||||
| Coliform bacteria (log10 CFU/g FM) | 3 | 7.72 a | 6.26 b,A | 8.24 a,A | 5.80 b,A | 0.181 | <0.01 | <0.01 | <0.01 | <0.01 | 0.05 | 0.34 |
| 7 | 7.89 b | 5.97 c,A | 8.35 a,A | <2.00 d,B | ||||||||
| 14 | 7.88 a | <2.00 b,B | 6.56 a,B | <2.00 b,B | ||||||||
| 30 | 7.37 a | <2.00 b,B | <2.00 b,C | <2.00 b,B | ||||||||
| Yeasts (log10 CFU/g FM) | 3 | <2.00 | <2.00 | <2.00 | <2.00 | - | - | - | - | - | - | - |
| 7 | <2.00 | <2.00 | <2.00 | <2.00 | ||||||||
| 14 | <2.00 | <2.00 | <2.00 | <2.00 | ||||||||
| 30 | <2.00 | <2.00 | <2.00 | <2.00 | ||||||||
| Molds (log10 CFU/g FM) | 3 | <2.00 | <2.00 | <2.00 | <2.00 | - | - | - | - | - | - | - |
| 7 | <2.00 | <2.00 | <2.00 | <2.00 | ||||||||
| 14 | <2.00 | <2.00 | <2.00 | <2.00 | ||||||||
| 30 | <2.00 | <2.00 | <2.00 | <2.00 | ||||||||
| Item | Days | 15 °C | 30 °C | SEM | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | LP | CK | LP | T | D | L | T × D | T × L | D × L | |||
| Sobs | 3 | 236 A | 241 B,C | 258 A | 297 A | 6.62 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | 0.245 |
| 7 | 178 d,B | 314 a,A | 247 b,B | 208 c,B | ||||||||
| 14 | 176 c,B | 214 b,c,C | 241 a,b,B | 276 a,A,B | ||||||||
| 30 | 230 b,A | 281 a,A,B | 256 a,b,A | 257 a,A,B | ||||||||
| Shannon | 3 | 1.77 b,B | 1.72 b | 2.28 a,B | 2.09 a,b,A | 0.100 | 0.681 | 0.066 | <0.01 | <0.01 | <0.01 | <0.01 |
| 7 | 2.16 b,B | 1.78 b,c | 2.71 a,A,B | 1.57 c,B | ||||||||
| 14 | 3.16 a,A | 1.70 c | 2.62 b,A,B | 1.29 c,B | ||||||||
| 30 | 2.84 a,A | 1.63 b | 3.26 a,A | 0.98 c,C | ||||||||
| Simpson | 3 | 0.54 b,B | 0.57 a,b,A | 0.61 a,B | 0.62 a,A | 0.0253 | <0.01 | 0.023 | <0.01 | <0.01 | <0.01 | <0.01 |
| 7 | 0.60 b,B | 0.60 b,A | 0.77 a,A | 0.45 c,B | ||||||||
| 14 | 0.80aA | 0.54bAB | 0.77aA | 0.34cC | ||||||||
| 30 | 0.75 a,A | 0.50 b,B | 0.79 a,A | 0.17 c,D | ||||||||
| Chao | 3 | 342 a,B | 296 b,C | 353 a | 367 a,A | 9.54 | 0.584 | <0.01 | 0.037 | 0.191 | <0.01 | <0.01 |
| 7 | 244 b,C | 395 a,A,B | 353 a | 266 b,B | ||||||||
| 14 | 259 b,C | 451 a,A | 369 a | 371 a,A | ||||||||
| 30 | 415 a,A | 361 b,B,C | 357 b | 373 b,A | ||||||||
| Ace | 3 | 351 a,B | 307 b,B | 346 a,b | 372 a,A,B | 9.23 | 0.018 | <0.01 | <0.01 | <0.01 | 0.027 | <0.01 |
| 7 | 241 b,C | 386 a,A | 354a | 251 b,C | ||||||||
| 14 | 235 b,C | 332 a,A,B | 353 a | 406 a,A | ||||||||
| 30 | 405 a,A | 355 b,A,B | 360 b | 358 b,B | ||||||||
| Goods_coverage | 3 | 0.999 | 0.999 | 0.999 | 0.999 | 0 | 0.783 | 0.252 | 0.455 | 0.204 | 0.020 | <0.01 |
| 7 | 0.999 | 0.999 | 0.999 | 1.000 | ||||||||
| 14 | 1.000 | 0.999 | 0.999 | 0.999 | ||||||||
| 30 | 0.999 | 0.999 | 0.999 | 0.999 | ||||||||
| Genus | Gas Production | CO2 Production | ||
|---|---|---|---|---|
| Correlation | Significance | Correlation | Significance | |
| Lactobacillus | −0.355 | <0.05 | −0.297 | <0.05 |
| Lactococcus | 0.177 | 0.229 | 0.12 | 0.426 |
| Methylobacterium | 0.102 | 0.492 | −0.235 | 0.116 |
| Pantoea | 0.178 | 0.225 | 0.035 | 0.819 |
| Enterococcus | 0.331 | <0.05 | 0.189 | 0.209 |
| Serratia | 0.189 | 0.199 | −0.306 | <0.05 |
| Weissella | 0.092 | 0.533 | −0.091 | 0.546 |
| Lachnoclostridium_5 | 0.533 | <0.01 | 0.136 | 0.366 |
| Aureimonas | −0.224 | 0.125 | −0.37 | <0.05 |
| Sphingomonas | −0.139 | 0.345 | −0.277 | 0.062 |
| Clostridium_sensu_stricto_1 | 0.319 | <0.05 | 0.005 | 0.974 |
| Escherichia-Shigella | 0.397 | <0.01 | 0.251 | 0.092 |
| Ralstonia | −0.22 | 0.133 | 0.072 | 0.633 |
| Hydrogenophaga | −0.157 | 0.285 | 0.174 | 0.246 |
| Aerococcus | −0.152 | 0.304 | −0.065 | 0.67 |
| Roseomonas | −0.078 | 0.596 | −0.097 | 0.521 |
| Acinetobacter | −0.197 | 0.18 | −0.277 | 0.063 |
| Pseudomonas | −0.119 | 0.419 | −0.299 | <0.05 |
| Bacillus | 0.182 | 0.216 | 0.029 | 0.85 |
| Prevotella_7 | −0.031 | 0.836 | 0.089 | 0.556 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, M.; Zhou, W.; Zou, X.; Wu, S.; Chen, X.; Zhang, Q. Fermentation Quality and Bacterial Diversity of Broussonetia papyrifera Leaves Ensiled with Lactobacillus plantarum and Stored at Different Temperatures. Agronomy 2022, 12, 986. https://doi.org/10.3390/agronomy12050986
Zheng M, Zhou W, Zou X, Wu S, Chen X, Zhang Q. Fermentation Quality and Bacterial Diversity of Broussonetia papyrifera Leaves Ensiled with Lactobacillus plantarum and Stored at Different Temperatures. Agronomy. 2022; 12(5):986. https://doi.org/10.3390/agronomy12050986
Chicago/Turabian StyleZheng, Mingyang, Wei Zhou, Xuan Zou, Shuo Wu, Xiaoyang Chen, and Qing Zhang. 2022. "Fermentation Quality and Bacterial Diversity of Broussonetia papyrifera Leaves Ensiled with Lactobacillus plantarum and Stored at Different Temperatures" Agronomy 12, no. 5: 986. https://doi.org/10.3390/agronomy12050986