
Topping Inhibited Potassium Uptake via Regulating Potassium Flux and Channel Gene Expression in Tobacco

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Treatments
2.2. Determination of K+ Concentration
2.3. Measurement of Root K+ Flux
2.4. Measurement of Hormone IAA Concentration
2.5. Expression Analysis of K+ Channel Genes
2.6. Statistical Analysis
3. Results
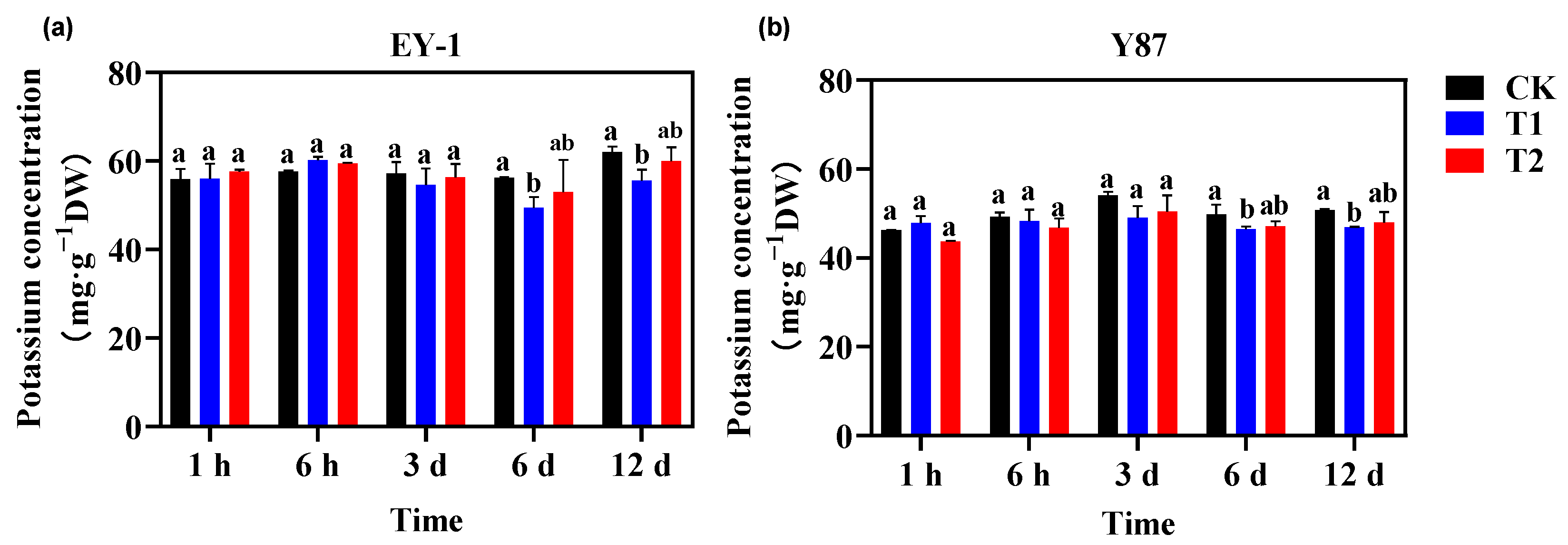
3.1. Effect of Topping on the K+ Concentration of Tobacco
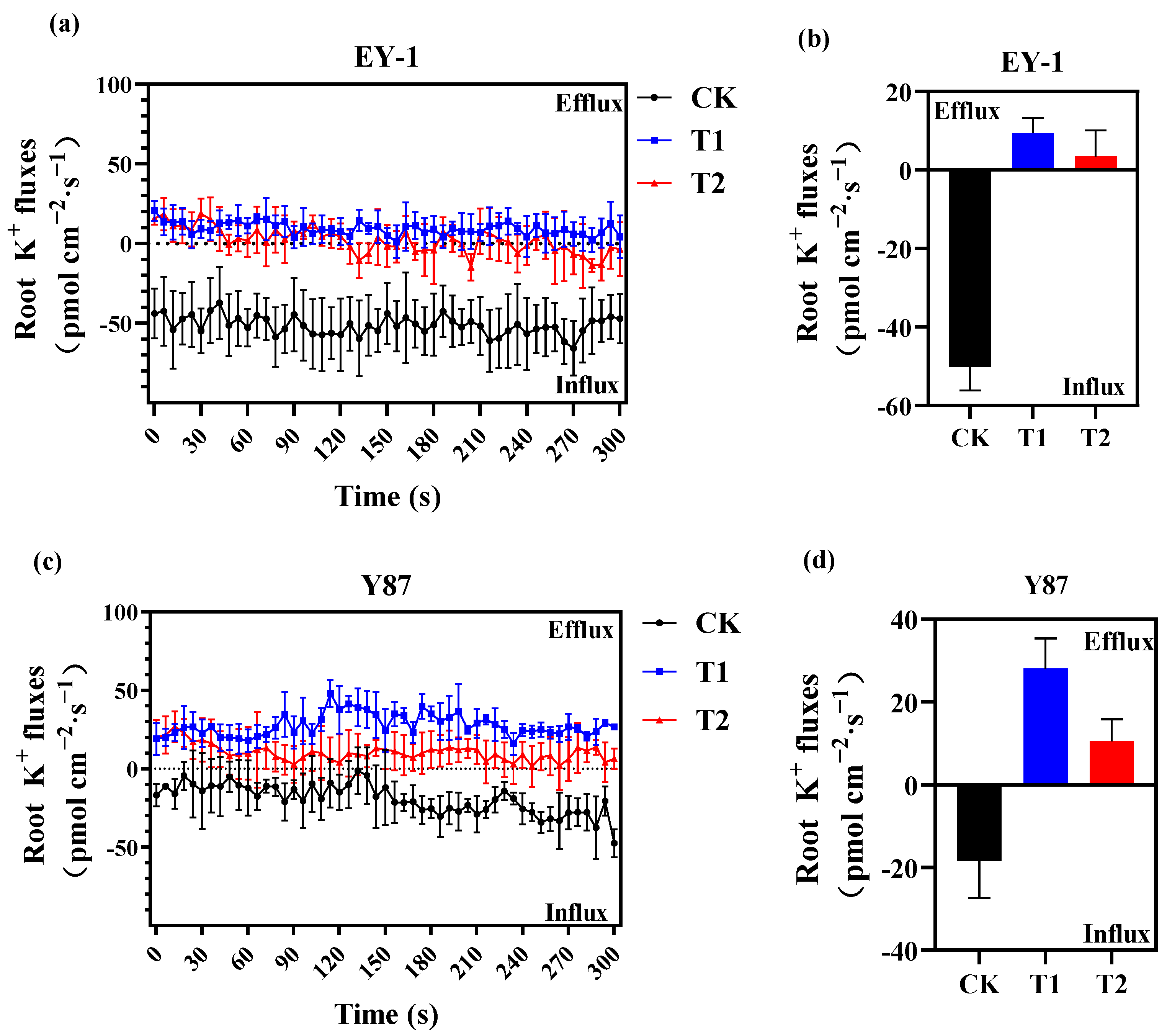
3.2. Effects of Topping on Root K+ Flux of Tobacco
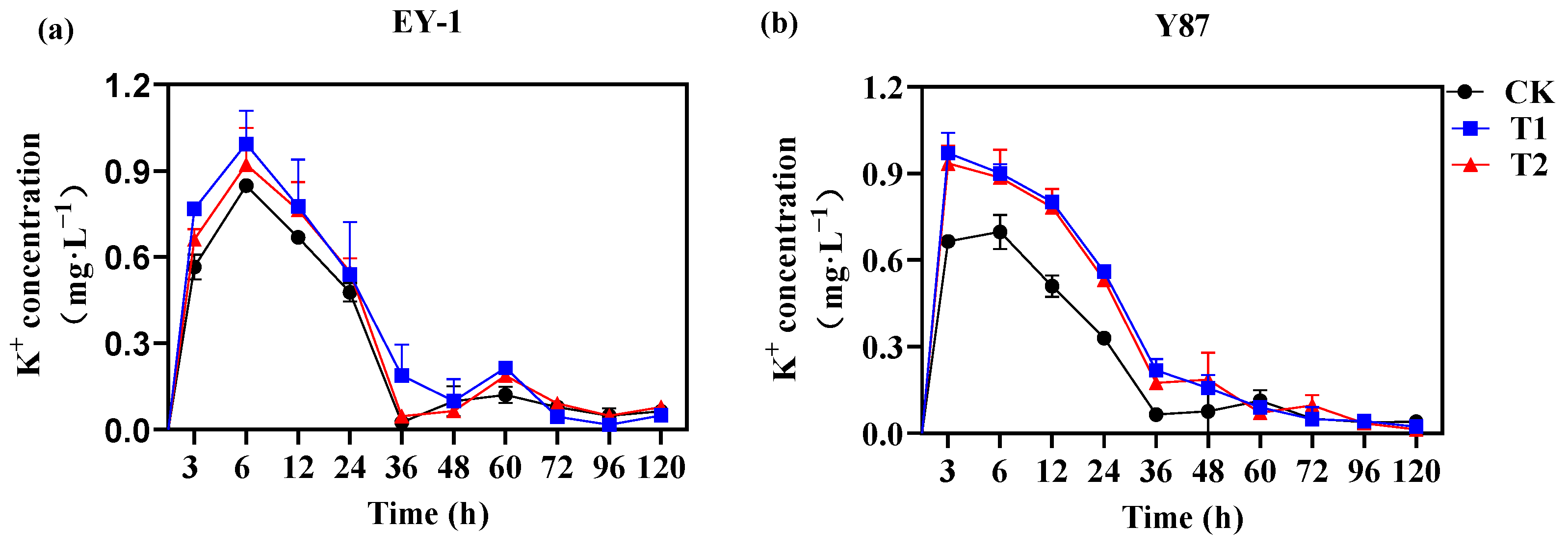
3.3. Effects of Topping on K+ Loss of Tobacco
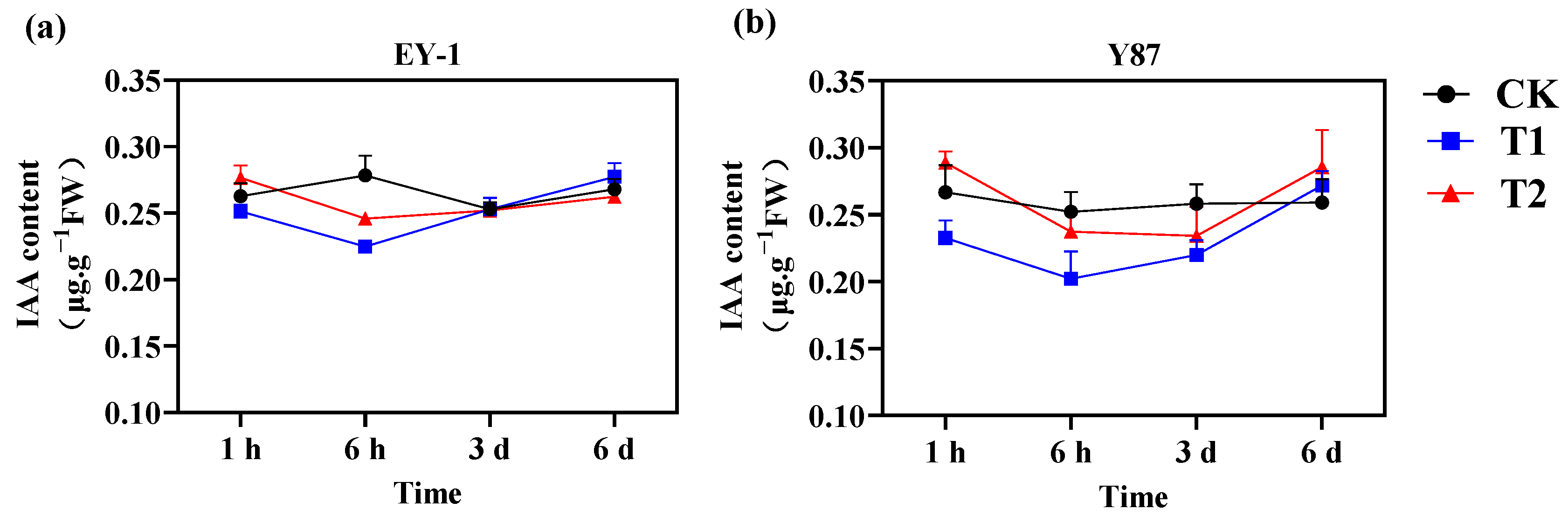
3.4. Effects of Topping on Tobacco Endogenous Hormone IAA Content
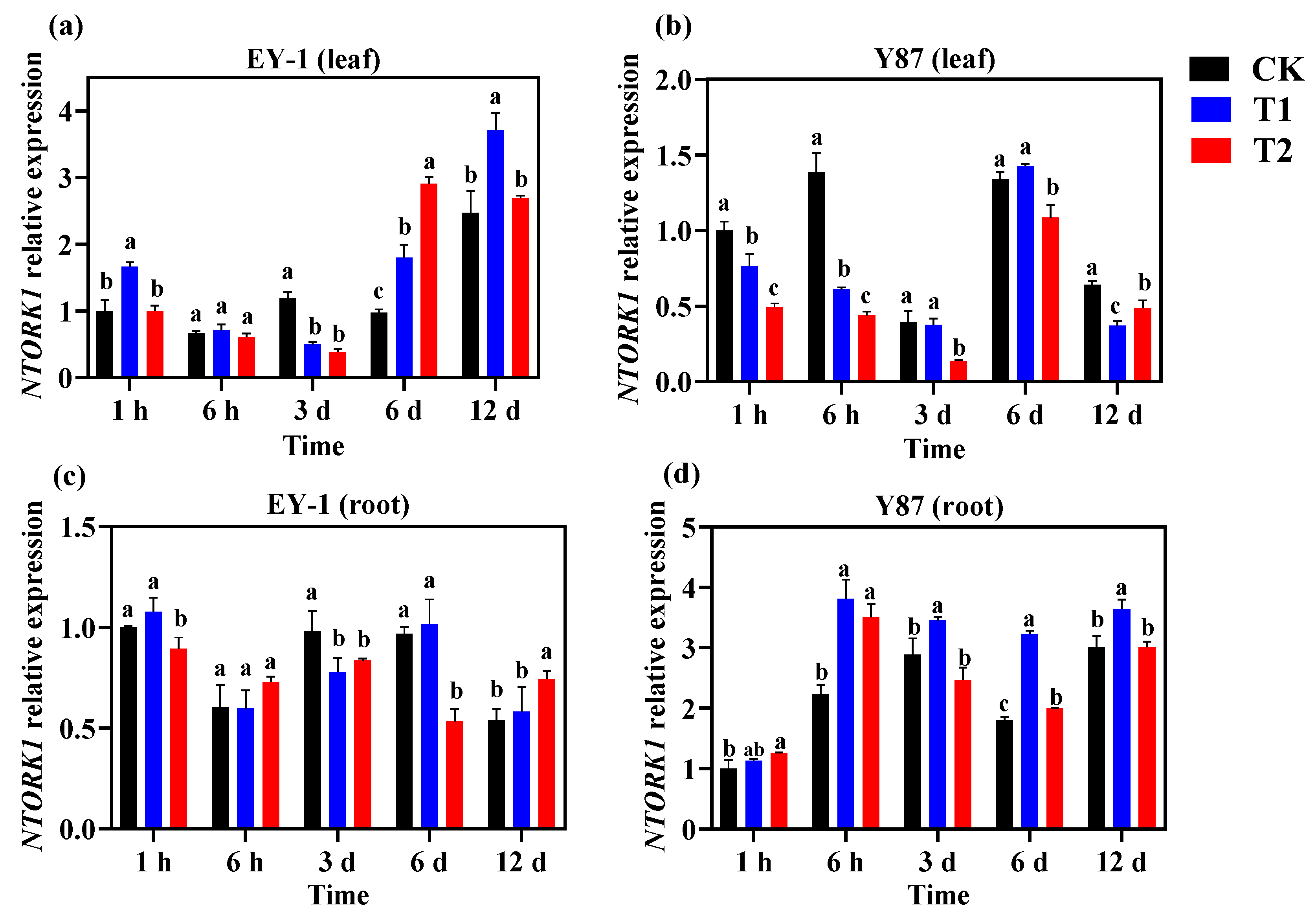
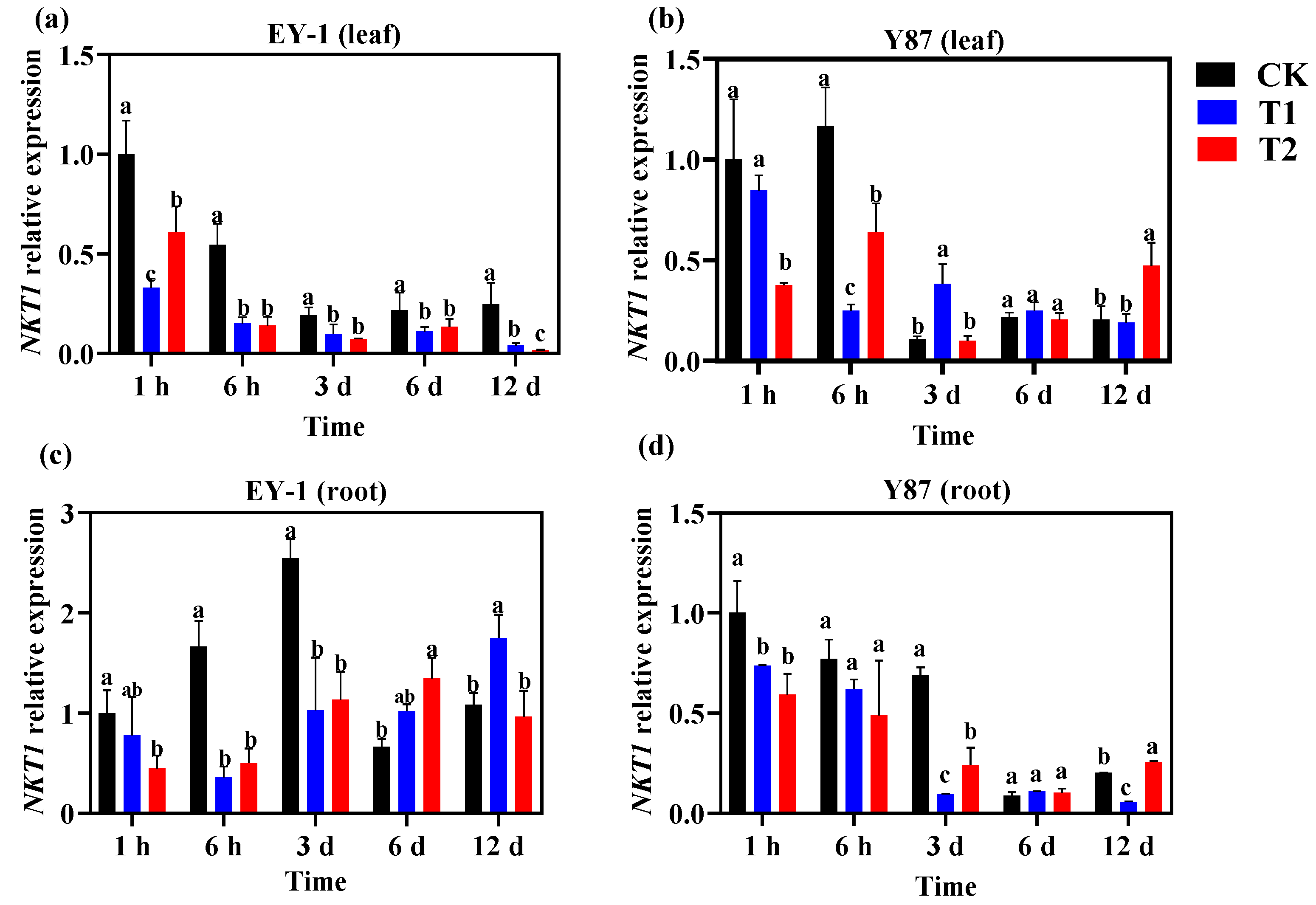
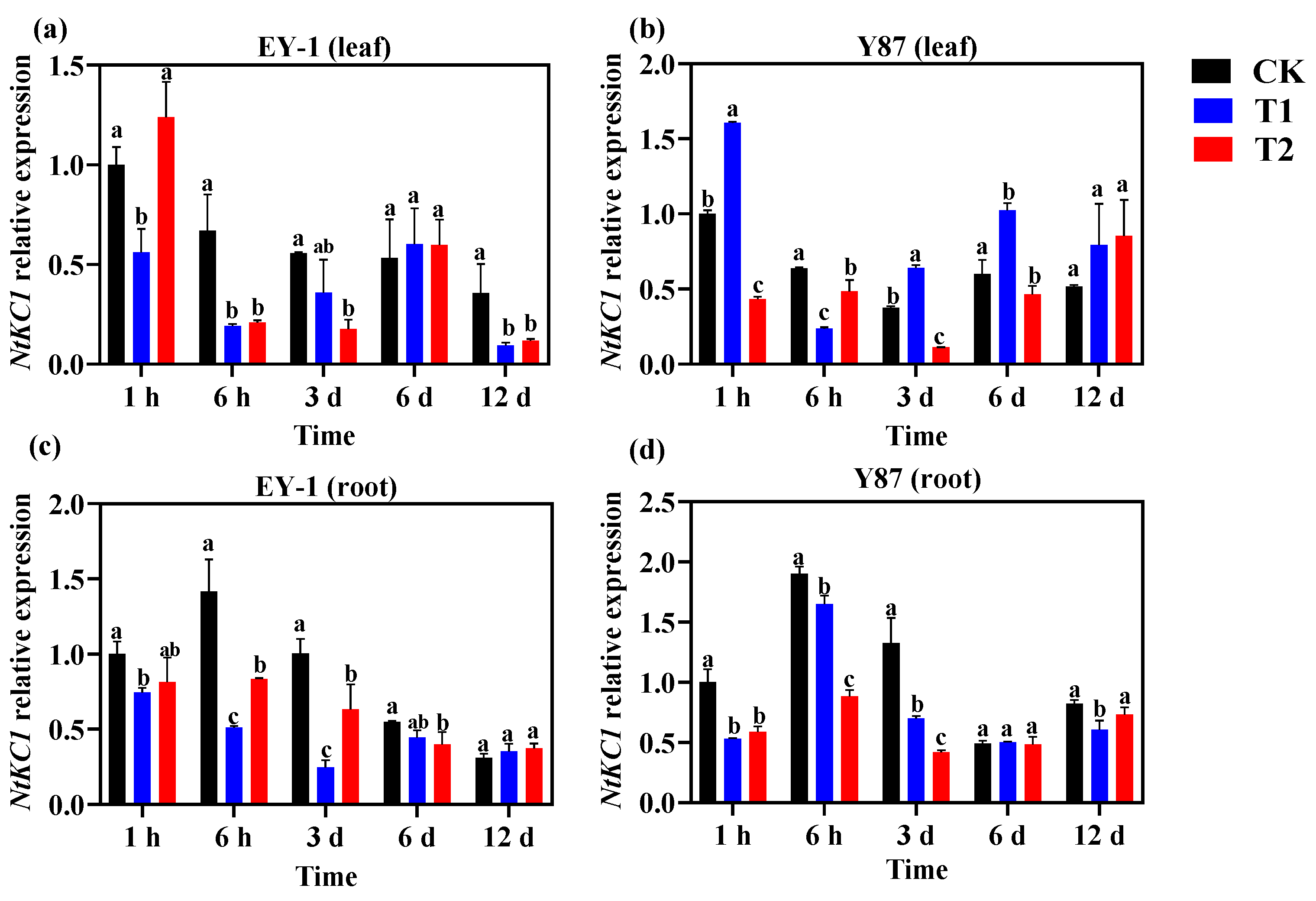
3.5. Effects of Topping on the Expression of K+ Channel Genes in Tobacco
3.5.1. Expression Analysis of K+ Efflux Channel Gene NTORK1
3.5.2. Expression Analysis of K+ Influx Channel Gene NKT1
3.5.3. Expression Analysis of K+ Influx Channel Gene NtKC1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dai, X.Y.; Sun, Y.S.; Wei, W.X.; Wu, J.S.; Fan, Y.K. Effects of top excision on the potassium accumulation and expression of potassium channel genes in tobacco. J. Exp. Bot. 2009, 60, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarkson, D.T.; Hanson, J.B. The mineral nutrition of higher plants. Annu. Rev. Plant Physiol. 2003, 31, 239–298. [Google Scholar] [CrossRef]
- Ul-Allah, S.; Ijaz, M.; Nawaz, A.; Sattar, A.; Sher, A.; Naeem, M.; Shahzad, U.; Nawaz, F.; Mahmood, K. Potassium application improves grain yield and alleviates drought susceptibility in diverse maize hybrids. Plants 2020, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieves-Cordones, M.; Alemán, F.; Martínez, V.; Rubio, F. K+ uptake in plant roots. The systems involved, their regulation and parallels in other organisms. J. Plant. Physiol. 2014, 171, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Battie-Laclau, P.; Laclau, J.P.; Beri, C.; Mietton, L.; Muniz, M.R.A.; Arenque, B.C.; Marisa, D.C.P.; Jordan-Meille, L.; Bouillet, J.P.; Nouvellon, Y. Photosynthetic and anatomical responses of eucalyptus grandis leaves to potassium and sodium supply in a field experiment. Plant. Cell. Environ. 2013, 37, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Peñuelas, J. Potassium Control of Plant Functions: Ecological and Agricultural Implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef]
- Monder, H.; Maillard, M.; Chérel, I.; Zimmermann, S.; Paris, N.; Cuéllar, T.; Gaillard, I. Adjustment of K+ fluxes and grapevine defense in the face of climate change. Int. J. Mol. Sci. 2021, 22, 10398. [Google Scholar] [CrossRef]
- Erel, R.; Yermiyahu, U.; Ben-Gal, A.; Dag, A.; Shapira, O.; Schwartz, A. Modification of non-stomatal limitation and photoprotection due to K and Na nutrition of olive trees. J. Plant. Physiol. 2015, 177, 1–10. [Google Scholar] [CrossRef]
- Lei, B.; Chan, W.; Zhao, H.; Zhang, K.; Yu, J.; Yu, S.; Cai, K.; Zhang, J.; Lu, K. Nitrogen application and differences in leaf number retained after topping affect the tobacco (Nicotiana tabacum) transcriptome and metabolome. BMC Plant. Biol. 2022, 22, 38. [Google Scholar] [CrossRef]
- Leggett, J.E.; Sims, J.L.; Gossett, D.R.; Pal, U.R.; Benner, J.F. Potassium and magnesium nutrition effects on yield and chemical composition of burley tobacco leaves and smoke. Can. J. Plant. Sci. 1977, 57, 159–166. [Google Scholar] [CrossRef]
- Su, Y.; Wei, W.; Chen, F.; Long, W.; Fan, Y.; Chen, X. Effects of topping on potassium metabolism and expression of potassium channels in tobacco plants. Sci. Agric. Sin. 2009, 42, 854–861. [Google Scholar]
- Hong, L.; Fu, L.; Su, F.; Wang, Y.; Zhai, X.; Tian, Y.; Zhu, D.; Liu, W. Effect of auxin on the potassium distribution and accumulation in flue-cured tobacco plants. Acta Agron. Sin. 2003, 29, 457–461. [Google Scholar]
- Zioni, A.; Vaadia, Y.; Lips, S. Nitrate uptake by roots as regulated by nitrate reduction products of the shoot. Physiol. Plant. 1971, 24, 288–290. [Google Scholar] [CrossRef]
- Dijkshoorn, W. Nitrate accumulation, nitrogen balance and cation-anion ratio during the regrowth of perennial ryegrass. Neth. J. Agric. Sci. 1958, 6, 211–221. [Google Scholar] [CrossRef]
- Kirkby, A. Estimation of potassium recirculation in tomato plants by comparison of the rates of potassium and calcium accumulation in the tops with their fluxes in the xylem stream. Plant Physiol. 1979, 63, 1143–1148. [Google Scholar]
- Jeschke, W.; Klagges, S.; Hilpert, A.; Bhatti, A.; Sarwar, G. Partitioning and flows of ions and nutrients in salt-treated plants of Leptochloa fusca L. Kunth. New Phytol. 1995, 130, 23–35. [Google Scholar] [CrossRef]
- Wang, X.; Wang, B.; Song, Z.; Zhao, L.; Ruan, W.; Gao, Y.; Jia, X.; Yi, K. A spatial–temporal understanding of gene regulatory networks and NtARF-mediated regulation of potassium accumulation in tobacco. Planta 2021, 255, 9. [Google Scholar] [CrossRef] [PubMed]
- Jie, L.; Miao, L.; Cai, R.; Wu, H.; Xu, H.W.; Zhou, X.F. Application of non-invasive micro-test technique in study of plant roots growth and development. Biotechnology 2013, 13, 89–93. [Google Scholar]
- Thor, K.; Jiang, S.; Michard, E.; George, J.; Scherzer, S.; Huang, S.; Dindas, J.; Derbyshire, P.; Leitão, N.; DeFalco, T.A.; et al. The calcium-permeable channel OSCA1.3 regulates plant stomatal immunity. Nature 2020, 585, 569–573. [Google Scholar] [CrossRef]
- Li, C.; Zhang, T.P.; Feng, P.W.; Li, D.X.; Bresticb, M.; Liu, Y.; Yang, X.H. Genetic engineering of glycinebetaine synthesis enhances cadmium tolerance in BADH-transgenic tobacco plants via reducing cadmium uptake and alleviating cadmium stress damage. Environ. Exp. Bot. 2021, 191, 104602. [Google Scholar] [CrossRef]
- Meng, X.; Liang, T.; Liu, F.; Yu, H.; Hu, L.; Zhang, Y.; Zhou, H.; Zhang, S.; Yin, Q. Potassium absorption and kinetics by different tobacco varieties at different potassium efficiency. Tob. Sci. Technol. 2019, 1, 1–7. [Google Scholar]
- Shu, H.Y.; Changm, S.H.; Yangm, T.Z. Preliminary study on potassium efflux from tobacco and its prevention. Crops 2005, 2, 16–17. [Google Scholar]
- Wang, A.G.; Zhang, S.X.; Dai, H.X.; Wang, G.S.; Zhang, X.J.; Zhang, Y.L. Effects of cement production on surrounding environment and heavy metal accumulation in tobacco leaves. Tob. Sci. Technol. 2018, 51, 7–14. [Google Scholar]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000; p. 270. [Google Scholar]
- Yang, T.Y.; Zhang, S.; Hu, Y.B.; Wu, F.C.; Hu, Q.D.; Chen, G.; Jiang, C. The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels. Plant Physiol. 2014, 166, 945–959. [Google Scholar] [CrossRef] [Green Version]
- Meulebroek, L.V.; Bussche, J.V.; Steppe, K.; Vanhaecke, L. Ultra-high performance liquid chromatography coupled to high resolution Orbitrap mass spectrometry for metabolomic profiling of the endogenous phytohormonal status of the tomato plant. J. Chromatogr. A 2012, 1260, 67–80. [Google Scholar] [CrossRef]
- Guo, Y.D.; Meng, X.Y.; Di, H.X.; Chen, Q.S.; Zhai, Z.; Wang, A.G.; Liang, T.B.; Yin, Q.S. Effects of nano-carbon sol on expression of K+ channel in tobacco and K+ fluxes in tobacco roots. Tob. Sci. Technol. 2021, 4, 1–7. [Google Scholar]
- Coskun, D.; Britto, D.; Kronzucker, H. The nitrogen–potassium intersection: Membranes, metabolism, and mechanism. Plant. Cell. Environ. 2017, 40, 2029–2041. [Google Scholar] [CrossRef]
- Akhtar, M.N.; Ul-Haq, T.; Ahmad, F.; Imran, M.; Ahmed, W.; Ghaffar, A.; Shahid, M.; Saleem, M.H.; Alshaya, H.; Okla, M.K.; et al. Application of potassium along with nitrogen under varied moisture regimes improves performance and mitrogen-use efficiency of high- andlow-potassium efficiency cotton cultivars. Agronomy 2022, 12, 502. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, Y.; Chen, X.; Xu, F.; Ding, M.; Ye, W.; Kawai, Y.; Toda, Y.; Hayashi, Y.; Suzuki, T.; et al. Plasma membrane H+-ATPase overexpression increases rice yield via simultaneous enhancement of nutrient uptake and photosynthesis. Nat. Commun. 2021, 12, 735. [Google Scholar] [CrossRef]
- Gaymard, F.; Pilot, G.; Lacombe, B.; Bouchez, D.; Bruneau, D.; Boucherez, J.; Michaux-Ferrière, N.; Thibaud, J.B.; Sentenac, H. Identification and disruption of a plant shaker-like outward channel involved in K+ release into the xylem sap. Cell 1998, 94, 647–655. [Google Scholar] [CrossRef] [Green Version]
- Basset, M.; Conejero, G.; Lepetit, M.; Fourcroy, P.; Sentenac, H. Organization and expression of the gene coding for the potassium transport system AKT1 of Arabidopsis thaliana. Plant Mol. Biol. 1995, 29, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, D.; Basset, M.; Lepetit, M.; Conejero, G.; Gaymard, F.; Astruc, S.; Grignon, C. Tissue-specific expression of Arabidopsis AKT1 gene is consistent with a role in K+ nutrition. Plant J. 1996, 9, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Vishwakarma, K.; Hossen, M.; Shahadat, K.V.; Shackira, A.M.; Puthur, J.T.; Abdi, G.; Sarraf, M.; Hasanuzzaman, M. Potassium in plants: Growth regulation, signaling, and environmental stress tolerance. Plant Physiol. Bioch. 2022, 172, 56–69. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Meng, L.; Tang, L.N.; Qi, W.C.; Hu, F.Q.; Lv, Y.D.; Song, J.W. The landscape of alternative splicing regulating potassium use efficiency in Nicotiana tabacum. Front. Plant Sci. 2021, 12, 774829. [Google Scholar] [CrossRef]
- Jin, W.H.; Zhou, Q.; Wei, Y.F.; Jinmiao, Y.; Hao, F.S.; Cheng, Z.P.; Guo, H.X.; Liu, W.Q. NtWRKY-R1, a novel transcription factor, integrates IAA and JA signal pathway under topping damage stress in Nicotiana tabacum. Front. Plant Sci. 2018, 8, 2263. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.M.; Yang, T.Z. Cloning and expression profile analysis of a putative potassium transporter gene NTHAK1 in tobacco. J. Nucl. Agric. Sci. 2011, 25, 469–476. [Google Scholar]
- Sun, J.; Chen, S.L.; Dai, S.X.; Wang, R.G.; Li, N.Y.; Shen, X.; Zhou, X.Y. NaCl-induced alternations of cellular and tissue ion fluxes in roots of salt-resistant and salt-sensitive poplar species. Plant Physiol. 2009, 149, 1141–1153. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.X.; Li, C.J.; Yang, Y.H.; Zhang, F.S. Why does potassium concentration in flue-cured tobacco leaves decrease after apex excision? Field. Crop. Res. 2010, 116, 86–91. [Google Scholar]
- Skalický, V.; Vojtková, T.; Pěnčík, A.; Vrána, J.; Juzoń, K.; Koláčková, V.; Sedlářová, M.; Kubeš, M.F.; Novák, O. Auxin metabolite profiling in isolated and intact plant nuclei. Int. J. Mol. Sci. 2021, 22, 12369. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Transcript ID | Primer Sequence (5′–3′) |
|---|---|---|
| NKT1 | AB196790 | F: GGCTCGTCTAACGGCAGATT |
| R: CAAGCACAACCCTTCCACCT | ||
| NtKC1 | AB196791 | F: CACTATTGTCATGGCGGATG |
| R: TCTTCGGTACATCCGTTTCTG | ||
| NTORK1 | AB196792 | F: AGTGAAACAACTTGAGAGTACCTC |
| R: GAGAAGCATAAACTGCTACAGTGG | ||
| ACTIN | AB158612 | F: AACAGTTTGGTTGGAGTTCTGG |
| R: CATGAAGATTAAAGGCGGAGTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, T.; Dai, H.; Khan, W.A.; Guo, Y.; Meng, X.; Wang, G.; Zhang, Y. Topping Inhibited Potassium Uptake via Regulating Potassium Flux and Channel Gene Expression in Tobacco. Agronomy 2022, 12, 1166. https://doi.org/10.3390/agronomy12051166
Liang T, Dai H, Khan WA, Guo Y, Meng X, Wang G, Zhang Y. Topping Inhibited Potassium Uptake via Regulating Potassium Flux and Channel Gene Expression in Tobacco. Agronomy. 2022; 12(5):1166. https://doi.org/10.3390/agronomy12051166
Chicago/Turabian StyleLiang, Taibo, Huaxin Dai, Waleed Amjad Khan, Yadi Guo, Xiangyu Meng, Guiyao Wang, and Yanling Zhang. 2022. "Topping Inhibited Potassium Uptake via Regulating Potassium Flux and Channel Gene Expression in Tobacco" Agronomy 12, no. 5: 1166. https://doi.org/10.3390/agronomy12051166