
Field Resistance of Digitaria sanguinalis (L.) Scop. to Haloxyfop-P-methyl in China’s Cotton Fields


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Dose Response to Haloxyfop-P-methyl
2.3. ACCase Activity Assays
2.4. GST Activity Assays
2.5. NADPH Activity Assays
2.6. CarE Activity Assays
3. Statistical Analysis
4. Results
4.1. Dose Response to Haloxyfop-P-methyl
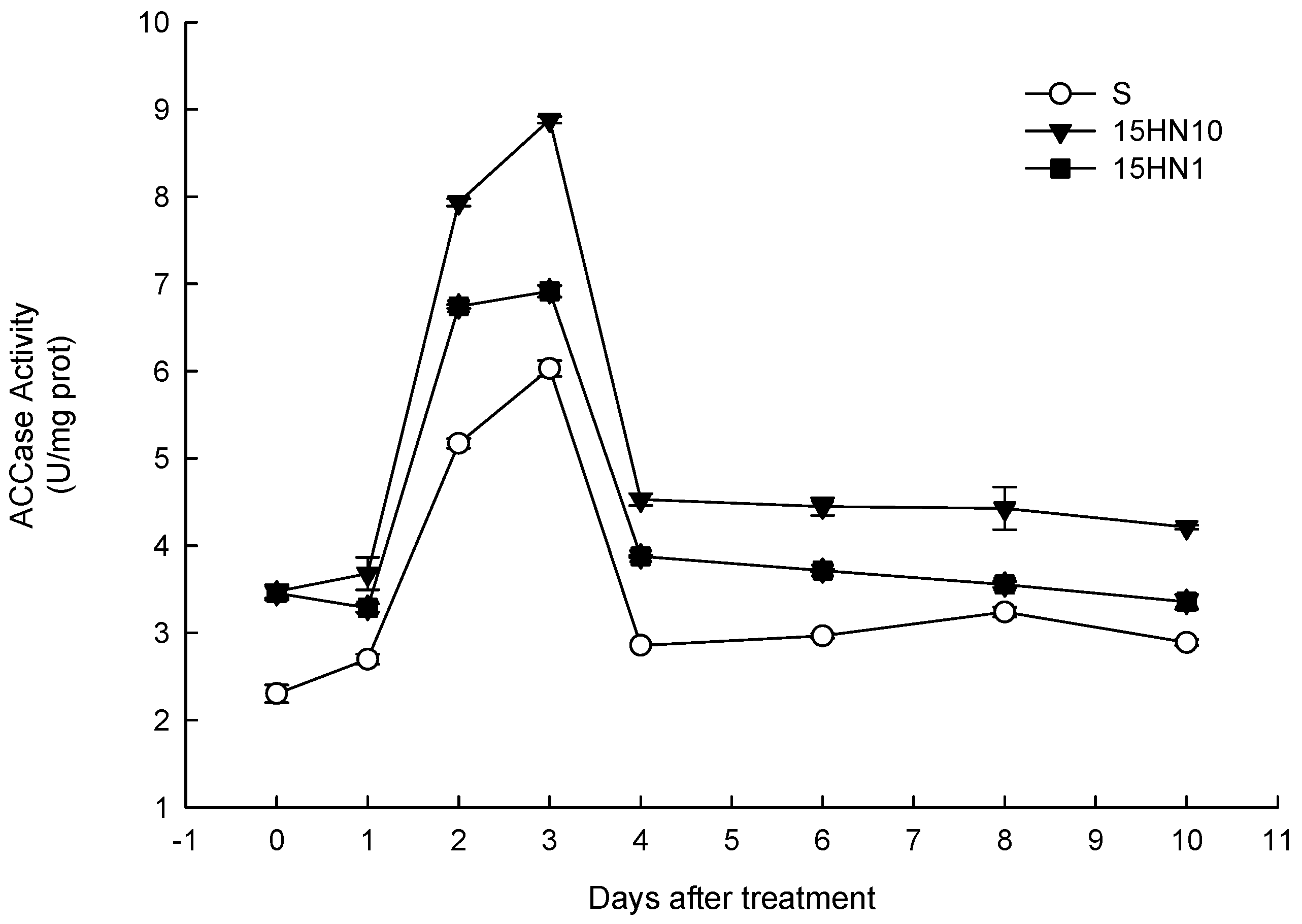
4.2. ACCase Activity
4.3. GST Activity
4.4. NADPH Activity
4.5. CarE Activity
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2022. Available online: http://www.weedscience.com (accessed on 25 April 2022).
- Dai, J.; Dong, H. Intensive cotton farming technologies in China: Achievements, challenges and countermeasures. Field Crop. Res. 2014, 155, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Wang, J.; DiTommaso, A.; Zhang, C.; Zheng, G.; Liang, W.; Islam, F.; Yang, C.; Chen, X.; Zhou, W. Weed research status, challenges, and opportunities in China. Crop Prot. 2020, 134, 104449. [Google Scholar] [CrossRef]
- Lu, D.; Bai, S.; Li, X.; Wang, G. Study on the population ecology and control of ascendant crab grass in summer grain crops in Hebei Province. Acta Phytoecol. Sin. 1995, 1, 55–63. [Google Scholar]
- Mei, Y.; Si, C.; Liu, M.; Qiu, L.; Zheng, M. Investigation of resistance levels and mechanisms to nicosulfuron conferred by non-target-site mechanisms in large crabgrass (Digitaria sanguinalis L.) from China. Pestic. Biochem. Physiol. 2017, 141, 84–89. [Google Scholar] [CrossRef]
- Editorial Committee of Flora of China. Flora Reipublicae Popularis Sinicae; Beijing Science and Technology Press: Beijing, China, 1990; Volume 10, p. 329. [Google Scholar]
- Gherekhloo, J.; Osuna, M.D.; De Prado, R. Biochemical and molecular basis of resistance to ACCase-inhibiting herbicides in Iranian Phalaris minor populations. Weed Res. 2012, 52, 367–372. [Google Scholar] [CrossRef]
- Letouzé, A.; Gasquez, J. Inheritance of fenoxaprop-P-ethyl resistance in a blackgrass (Alopecurus myosuroides Huds.) population. Theor. Appl. Genet. 2001, 103, 288–296. [Google Scholar] [CrossRef]
- Wiederholt, R.J.; Stoltenberg, D.E. Cross-resistance of a large crabgrass (Digitaria sanguinalis) accessions to aryloxyphenoxypropionate and cyclohexane-dione herbicides. Weed Technol. 1995, 9, 518–524. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in Action: Plants Resistant to Herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [Green Version]
- Hidayat, I.; Preston, C. Enhanced Metabolism of Fluazifop Acid in a Biotype of Digitaria sanguinalis Resistant to the Herbicide Fluazifop-P-Butyl. Pestic. Biochem. Physiol. 1997, 57, 137–146. [Google Scholar] [CrossRef]
- Volenberg, D.; Stoltenberg, D. Altered acetyl-coenzyme A carboxylase confers resistance to clethodim, fluazifop and sethoxydim in Setaria faberi and Digitaria sanguinalis. Weed Res. 2002, 42, 342–350. [Google Scholar] [CrossRef]
- Liu, X.; Merchant, A.; Xiang, S.; Zong, T.; Zhou, X.; Bai, L. Managing herbicide resistance in China. Weed Sci. 2021, 69, 4–17. [Google Scholar] [CrossRef]
- Cruz-Hipólito, H.; Fernández, P.; Alcántara, R.; Gherekhloo, J.; Osuna, M.D.; De Prado, R. Ile-1781-Leu and Asp-2078-Gly Mutations in ACCase Gene, Endow Cross-resistance to APP, CHD, and PPZ in Phalaris minor from Mexico. Int. J. Mol. Sci. 2015, 16, 21363–21377. [Google Scholar] [CrossRef] [Green Version]
- Shergill, L.S.; Malone, J.; Boutsalis, P.; Preston, C.; Gill, G. Basis of ACCase and ALS inhibitor resistance in Hordeum glaucum Steud. Pest Manag. Sci. 2017, 73, 1638–1647. [Google Scholar] [CrossRef]
- Zhao, N.; Ge, L.; Yan, Y.; Bai, S.; Wang, D.; Liu, W.; Wang, J. Trp-1999-Ser mutation of acetyl-CoA carboxylase and cytochrome P450s-involved metabolism confer resistance to fenoxaprop-P-ethyl in Polypogon fugax. Pest Manag. Sci. 2019, 75, 3175–3183. [Google Scholar] [CrossRef]
- Hochberg, O.; Sibony, M.; Rubin, B. The response of ACCase-resistant Phalaris paradoxa populations involves two different target site mutations. Weed Res. 2009, 49, 37–46. [Google Scholar] [CrossRef]
- Kaundun, S.S.; Hutchings, S.-J.; Dale, R.P.; McIndoe, E. Broad Resistance to ACCase Inhibiting Herbicides in a Ryegrass Population Is Due Only to a Cysteine to Arginine Mutation in the Target Enzyme. PLoS ONE 2012, 7, e39759. [Google Scholar] [CrossRef] [Green Version]
- Laforest, M.; Soufiane, B.; Simard, M.-J.; Obeid, K.; Page, E.; Nurse, R.E. Acetyl-CoA carboxylase overexpression in herbicide-resistant large crabgrass (Digitaria sanguinalis). Pest Manag. Sci. 2017, 73, 2227–2235. [Google Scholar] [CrossRef]
- Jugulam, M.; Shyam, C. Non-Target-Site Resistance to Herbicides: Recent Developments. Plants 2019, 8, 417. [Google Scholar] [CrossRef] [Green Version]
- Devine, M.D. Physiological, biochemical and molecular aspects of herbicide resistance based on altered target sites. Herbic. Act. Toxicol. Biochem. Mol. Biol. 1997, 2, 159–185. [Google Scholar]
- Preston, C. Inheritance and linkage of metabolism-based herbicide cross-resistance in rigid ryegrass (Lolium rigidum). Weed Sci. 2003, 51, 4–12. [Google Scholar] [CrossRef]
- Tal, A.; Rubin, B. Molecular characterization and inheritance of resistance to ACCase-inhibiting herbicides in Lolium rigidum. Pest Manag. Sci. 2004, 60, 1013–1018. [Google Scholar] [CrossRef]
- Délye, C. Unravelling the genetic bases of non-target-site-based resistance (NTSR) to herbicides: A major challenge for weed science in the forthcoming decade. Pest Manag. Sci. 2013, 69, 176–187. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S. Metabolism-Based Herbicide Resistance and Cross-Resistance in Crop Weeds: A Threat to Herbicide Sustainability and Global Crop Production. Plant Physiol. 2014, 166, 1106–1118. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.; Blake-Kalff, M.; Davies, T.G.E. Detoxification of xenobiotics by plants: Chemical modification and vacuolar compartmentation. Trends Plant Sci. 1997, 2, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Dixon, D.P.; Lapthorn, A.; Edwards, R. Plant glutathione transferases. Genome Biol. 2002, 3, reviews3004.1. [Google Scholar] [CrossRef] [Green Version]
- Sasan, M.; Maryam, E.; Fateme, M.; Maryam, S.; Babak, S.; Hassan, M. Plant glutathione S-transferase classification, structure and evolution. Afr. J. Biotechnol. 2011, 10, 8160–8165. [Google Scholar] [CrossRef] [Green Version]
- Werck-Reichhart, D.; Hehn, A.; Didierjean, L. Cytochromes P450 for engineering herbicide tolerance. Trends Plant Sci. 2000, 5, 116–123. [Google Scholar] [CrossRef]
- Siminszky, B. Plant cytochrome P450-mediated herbicide metabolism. Phytochem. Rev. 2006, 5, 445–458. [Google Scholar] [CrossRef]
- Barkay, D.B.; Reaney, S.H.; Langston, W.J.; Di Monte, D.A. Redox cycling of the herbicide paraquat in microglial cultures. Mol. Brain Res. 2005, 134, 52–56. [Google Scholar] [CrossRef]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Burton, J.D.; Gronwald, J.W.; Keith, R.A.; Somers, D.A.; Gengenbach, B.G.; Wyse, D.L. Kinetics of inhibition of acetyl-coenzyme A carboxylase by sethoxydim and haloxyfop. Pestic. Biochem. Physiol. 1991, 39, 100–109. [Google Scholar] [CrossRef]
- Gronwald, J.W. Lipid Biosynthesis Inhibitors. Weed Sci. 1991, 39, 435–449. [Google Scholar] [CrossRef]
- Numa, S.; Bortz, W.M.; Lynen, F. Regulation of fatty acid synthesis at the acetyl-CoA carboxylation step. Adv. Enzym. Regul. 1965, 3, 407–423. [Google Scholar] [CrossRef]
- Sasaki, Y.; Konishi, T.; Nagano, Y. The Compartmentation of Acetyl-Coenzyme A Carboxylase in Plants. Plant Physiol. 1995, 108, 445–449. [Google Scholar] [CrossRef]
- Cummins, I.; Edwards, R. Purification and cloning of an esterase from the weed black-grass (Alopecurus myosuroides), which bioactivates aryloxyphenoxypropionate herbicides. Plant J. 2004, 39, 894–904. [Google Scholar] [CrossRef]
- Haslam, R.; Raveton, M.; Cole, D.J.; Pallett, K.E.; Coleman, J.O. The Identification and Properties of Apoplastic Carboxylesterases from Wheat That Catalyse Deesterification of Herbicides. Pestic. Biochem. Physiol. 2001, 71, 178–189. [Google Scholar] [CrossRef]
- Huang, S.; Wang, Q.; Zhang, S. Study on the resistance Alopecurus aequalis Sobol. to haloxyfop-R-methyl. J. Anhui Agric. Sci. 2006, 9, 1913–1914. (In Chinese) [Google Scholar]
- Li, J.; Zong, T.; Liu, X.; Bai, L. Resistance of Eleusine indica to haloxy-R-methyl in cotton fields in part of Hunan province. Cotton Sci. 2014, 26, 279–282. (In Chinese) [Google Scholar]
- Li, J.; Li, M.; Gao, X.; Fang, F. A novel amino acid substitution Trp574Arg in acetolactate synthase (ALS) confers broad resistance to ALS-inhibiting herbicides in crabgrass (Digitaria sanguinalis). Pest Manag. Sci. 2017, 73, 2538–2543. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Tang, W.; Yu, X.; Chen, J.; Xie, L.; Lu, Y. Tolerance to Some ACCase Inhibitors in Four Common Roegneria (Roegneria kamoji) Populations from China. Front. Agron. 2020, 2. [Google Scholar] [CrossRef]
- Zong, T.; Li, J.; Liu, X.; Bai, L. Resistance of Eleusine indica to quizalofop-P-ethyl in some cotton fields in Hunan province. Plant Prot. 2015, 41, 58–63. (In Chinese) [Google Scholar]
- Zand, E.; Baghestani, M.A.; Bitarafan, M.; Shimi, P. A Guideline for Herbicides in Iran; Jahad Publication: Mashhad, Iran, 2007; p. 158. [Google Scholar]
- Cocker, K.M.; Moss, S.R.; Coleman, J.O. Multiple Mechanisms of Resistance to Fenoxaprop-P-Ethyl in United Kingdom and Other European Populations of Herbicide-Resistant Alopecurus myosuroides (Black-Grass). Pestic. Biochem. Physiol. 1999, 65, 169–180. [Google Scholar] [CrossRef]
- Devine, M.D. Mechanisms of resistance to acetyl coenzyme A carboxylase: A review. Pestic. Sci. 1997, 51, 259–264. [Google Scholar] [CrossRef]
- Leach, G.; Devine, M.; Kirkwood, R.; Marshall, G. Target Enzyme-Based Resistance to Acetyl-Coenzyme A Carboxylase Inhibitors in Eleusine indica. Pestic. Biochem. Physiol. 1995, 51, 129–136. [Google Scholar] [CrossRef]
- Guo, W.; Chi, Y.; Feng, L.; Tian, X.; Liu, W.; Wang, J. Fenoxaprop-P-ethyl and mesosulfuron-methyl resistance status of shortawn foxtail (Alopecurus aequalis Sobol.) in eastern China. Pestic. Biochem. Physiol. 2018, 148, 126–132. [Google Scholar] [CrossRef]
- Tang, H.Y.; Wang, X.E.; Shen, G.H. Study on the yield loss of wheat and barley caused by the infestation of Alopecurus japonicus and A. aequalis. J. Weed Sci. 1990, 4, 8–12. [Google Scholar]
- Han, H.; Yu, Q.; Owen, M.J.; Cawthray, G.R.; Powles, S.B. Widespread occurrence of both metabolic and target-site herbicide resistance mechanisms in Lolium rigidum populations. Pest Manag. Sci. 2016, 72, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Ahmad-Hamdani, M.S.; Yu, Q.; Han, H.; Cawthray, G.R.; Wang, S.F.; Powles, S.B. Herbicide Resistance Endowed by Enhanced Rates of Herbicide Metabolism in Wild Oat (Avena spp.). Weed Sci. 2017, 61, 55–62. [Google Scholar] [CrossRef]
- Délye, C.; Gardin, J.A.C.; Boucansaud, K.; Chauvel, B.; Petit, C. Non-target-site-based resistance should be the centre of attention for herbicide resistance research: Alopecurus myosuroides as an illustration. Weed Res. 2011, 51, 433–437. [Google Scholar] [CrossRef]
- Huan, Z.-B.; Zhang, H.-J.; Hou, Z.; Zhang, S.-Y.; Zhang, Y.; Liu, W.; Bi, Y.-L.; Wang, J.-X. Resistance Level and Metabolism of Barnyard-Grass (Echinochloa crusgalli (L.) Beauv.) Populations to Quizalofop-p-ethyl in Heilongjiang Province, China. Agric. Sci. China 2011, 10, 1914–1922. [Google Scholar] [CrossRef]
- Kaundun, S.S. An aspartate to glycine change in the carboxyl transferase domain of acetyl CoA carboxylase and non-target-site mechanism(s) confer resistance to ACCase inhibitor herbicides in a Lolium multiflorum population. Pest Manag. Sci. 2010, 66, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Shankhar, K.S.; Bailly, G.C.; Dale, R.P.; Sarah-Jane, H.; Eddie, M.I.; Ingvarsson, P.K. A novel W1999S mutation and non-target site resistance impact on acetyl-CoA carboxylase inhibiting herbicides to varying degrees in a UK Lolium multiflorum population. PLoS ONE 2013, 8, e58012. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Correlation Coefficient | GR50 (g a.i./hm2) | Resistance Index |
|---|---|---|---|
| 14HN2 | 0.9701 | 5.722 | 3.115 |
| 14HN3 | 0.9059 | 6.648 | 3.619 |
| 14HN4 | 0.9584 | 3.189 | 1.736 |
| 14HN5 | 0.9571 | 3.350 | 1.824 |
| 14HN6 | 0.9906 | 1.921 | 1.046 |
| 14HN7 | 0.9601 | 2.114 | 1.151 |
| 14HN8 | 0.9075 | 2.824 | 1.537 |
| 14HN9 | 0.9544 | 16.430 | 8.944 |
| 14HN10 | 0.9500 | 18.150 | 9.880 |
| 14HN11 | 0.9822 | 9.446 | 5.142 |
| 14HN12 | 0.9850 | 15.098 | 8.219 |
| 14HN13 | 0.9888 | 5.801 | 3.158 |
| 14JX1 | 0.9516 | 11.519 | 6.271 |
| 14HEN1 | 0.9529 | 7.575 | 4.124 |
| 14HEN2 | 0.9736 | 11.038 | 6.009 |
| 14HB1 | 0.9823 | 4.483 | 2.440 |
| 14HB2 | 0.9688 | 22.845 | 12.436 |
| 14SX1 | 0.9903 | 9.299 | 5.062 |
| 14XJ1 | 0.9755 | 21.783 | 11.858 |
| 13HEB1 | 0.9669 | 18.751 | 10.207 |
| 13HEB2 | 0.9753 | 16.001 | 8.710 |
| 13AH1 | 0.9606 | 19.492 | 10.611 |
| 13HN1(S) | 0.9925 | 1.837 | 1.000 |
| Population | Correlation Coefficient | GR50 (g a.i./hm2) | Resistance Index |
|---|---|---|---|
| 15HN1 | 0.9729 | 28.089 | 16.770 |
| 15HN2 | 0.9530 | 15.032 | 8.974 |
| 15HN3 | 0.9549 | 6.384 | 3.811 |
| 15HN5 | 0.9650 | 4.909 | 2.931 |
| 15HN6 | 0.9550 | 14.557 | 8.691 |
| 15HN8 | 0.9512 | 8.067 | 4.816 |
| 15HN9 | 0.9523 | 10.897 | 6.506 |
| 15HN10 | 0.9502 | 16.834 | 10.050 |
| 15HN12 | 0.9534 | 14.439 | 8.620 |
| 15HEN1 | 0.9693 | 4.770 | 2.848 |
| 15HEN2 | 0.9821 | 3.788 | 2.261 |
| 15JX1 | 0.9775 | 3.977 | 2.374 |
| 15JX2 | 0.9701 | 3.346 | 1.998 |
| 15JX3 | 0.9665 | 6.529 | 3.898 |
| 15JX4 | 0.9590 | 4.268 | 2.548 |
| 15JX7 | 0.9948 | 2.656 | 1.586 |
| 15HB1 | 0.9596 | 17.407 | 10.392 |
| 15HEB2 | 0.9501 | 3.681 | 2.198 |
| 14HN1(S) | 0.9586 | 1.675 | 1.000 |
| Population | Correlation Coefficient | GR50 (g a.i./hm2) | Resistance Index |
|---|---|---|---|
| 16HN2 | 0.9867 | 8.004 | 4.717 |
| 16HN3 | 0.9776 | 6.711 | 3.955 |
| 16HN4 | 0.9764 | 4.535 | 2.673 |
| 16HN5 | 0.9917 | 5.892 | 3.472 |
| 16JX1 | 0.9168 | 11.558 | 6.811 |
| 16JX4 | 0.9801 | 5.258 | 3.099 |
| 16HEN1 | 0.9559 | 4.614 | 2.791 |
| 16HEB1 | 0.9777 | 5.051 | 2.976 |
| 16HEB4 | 0.9837 | 8.607 | 5.072 |
| 16HEB5 | 0.9939 | 6.757 | 3.982 |
| 16HEB6 | 0.9896 | 5.788 | 3.411 |
| 16HEB7 | 0.9884 | 3.938 | 2.320 |
| 14HN1(S) | 0.9819 | 1.697 | 1.000 |
| Population | Correlation Coefficient | GR50 (g a.i./hm2) | Resistance Index |
|---|---|---|---|
| 17HEB1 | 0.9856 | 22.888 | 12.399 |
| 17HEB2 | 0.9911 | 21.128 | 11.446 |
| 17HEB3 | 0.9804 | 22.991 | 12.454 |
| 17HEB4 | 0.9865 | 22.204 | 12.028 |
| 17HEB5 | 0.9927 | 22.670 | 12.281 |
| 17HEB6 | 0.9922 | 30.024 | 16.264 |
| 17HEB7 | 0.9719 | 21.777 | 11.797 |
| 17HEB8 | 0.9960 | 20.974 | 11.362 |
| 17HEB9 | 0.9887 | 10.502 | 5.689 |
| 17JX1 | 0.9722 | 7.998 | 4.333 |
| 17JX2 | 0.9717 | 14.148 | 7.664 |
| 17JX3 | 0.9883 | 21.739 | 11.776 |
| 14HN1(S) | 0.9920 | 1.846 | 1.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zong, T.; Li, J.; Zhou, X.; Liu, X. Field Resistance of Digitaria sanguinalis (L.) Scop. to Haloxyfop-P-methyl in China’s Cotton Fields. Agronomy 2022, 12, 1071. https://doi.org/10.3390/agronomy12051071
Zong T, Li J, Zhou X, Liu X. Field Resistance of Digitaria sanguinalis (L.) Scop. to Haloxyfop-P-methyl in China’s Cotton Fields. Agronomy. 2022; 12(5):1071. https://doi.org/10.3390/agronomy12051071
Chicago/Turabian StyleZong, Tao, Jie Li, Xuguo Zhou, and Xiangying Liu. 2022. "Field Resistance of Digitaria sanguinalis (L.) Scop. to Haloxyfop-P-methyl in China’s Cotton Fields" Agronomy 12, no. 5: 1071. https://doi.org/10.3390/agronomy12051071