
Prospective Practice for Compound Stress Tolerance in Thyme Plants Using Nanoparticles and Biochar for Photosynthesis and Biochemical Ingredient Stability


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Location
2.2. Irrigation System
2.3. Plant Material, Transplant, and Harvest Dates
2.4. Essential Oil Analysis
2.5. Biochar Preparation
2.6. Chemical Fertilizers
2.7. Nano-Zeolite Preparation
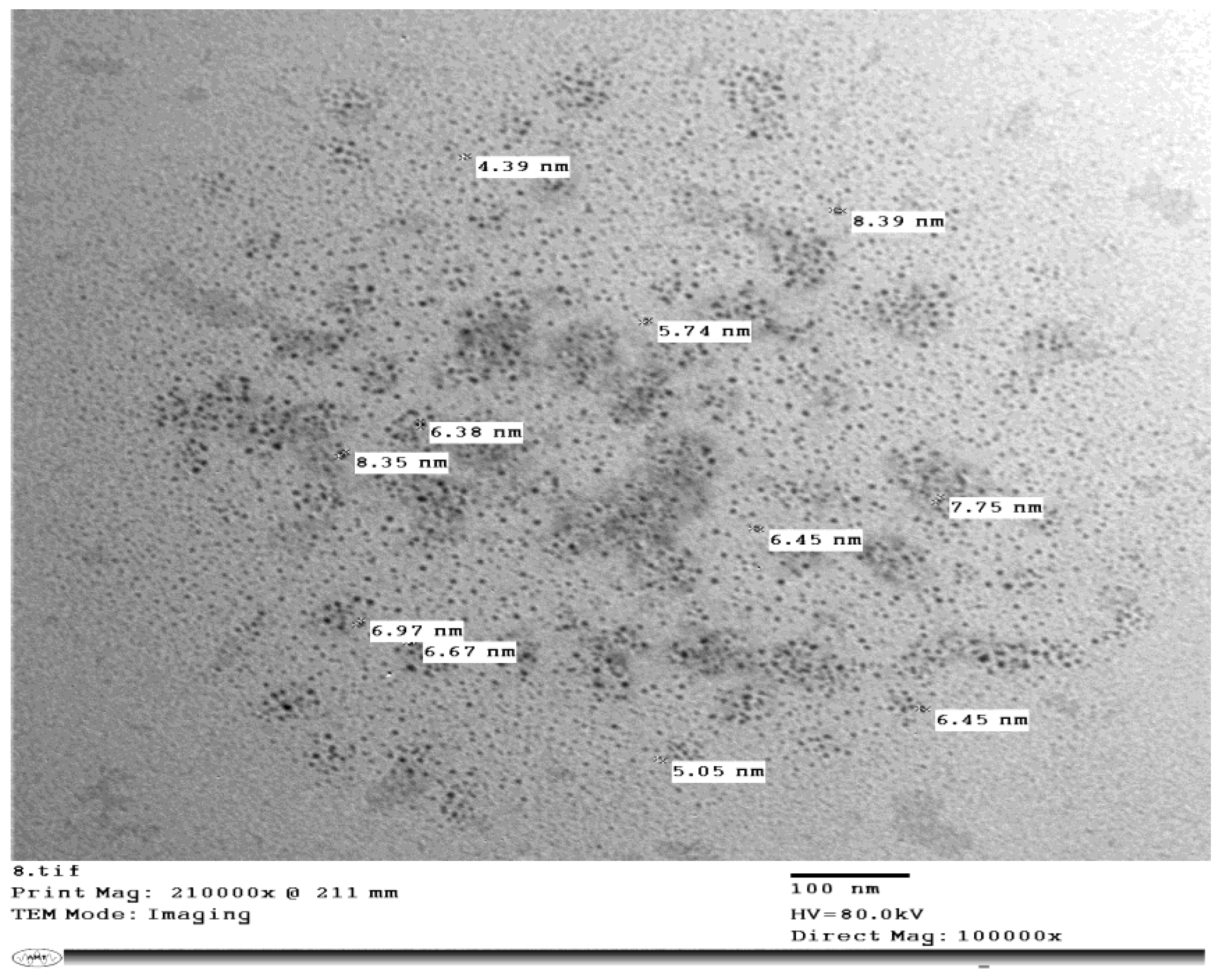
2.8. Synthesis of Silicon Nanoparticles
- -
- NPK + Organic matter (O) as a control T1
- -
- NPK + Nano Silicon (nS) + Organic matter (O) T2
- -
- Biochar (BC) + Organic matter(O) T3
- -
- Nano Silicon (nS) + Organic matter (O) T4
- -
- Nano Zeolite (nZ) + Nano Silicon (nS) + Organic matter (O) T5
- -
- Biochar (BC) + Nano Silicon (nS) + Organic matter (O) T6
- -
- Nano Zeolite (nZ) + Biochar (BC) + Nano Silicon (nS) + Organic matter (O) T7
2.9. Data Recorded
- -
- Plant height (cm)
- -
- Lateral branch numbers
- -
- Shoot fresh and dry weight (g plant−1)
- -
- Root fresh and dry weight (g plant−1)
- -
- Yield fresh weight (Kg/Fadden)
- -
- Plant Water content (%)
- -
- Photosynthetic rate (CO2 m−2 s−1)
- -
- Intercellular CO2 concentration (ppm)
- -
- Transpiration rate (mmol m−2s−1)
- -
- Plant Water use efficiency (μmol mmol−1)
- -
- Oil yield\L\Fadden.
2.10. Endogenous Phytohormones
2.11. The Activity of Antioxidant Enzymes
2.12. Statistical Analysis
3. Results and Discussion
3.1. Nanoparticles EnhancedPlant Growth, Yield, and Photosynthetic Parameters
3.2. Nanoparticles EnhancedPhotosynthetic Pigments and Biochemical Contentsof Thyme Plants
3.3. Nanoparticles Enhanced Leaf Hormones, and EnzymesActivity of Thyme Plants
3.4. Nanoparticles Enhanced Leaf Endogenous Nutrient Contents of Thyme Plants
3.5. Nanoparticles Enhanced Hydrocarbons and Oxygenated Compounds in Thyme Plant
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stahl-Biskup, E.; Venskutonis, R.P. Thyme. In Handbook of Herbs and Spices; Elsevier: Boca Raton, FL, USA, 2004; Volume 2, pp. 297–321. ISBN 9781855738355. [Google Scholar]
- Arzani, H.; Motamedi, A. Chemical compounds of Thyme as a medicinal herb in the mountainous areas of Iran. J. Nutr. Disord. Ther. 2013, 12, 003. [Google Scholar] [CrossRef] [Green Version]
- Fraire-Velázquez, S.; Balderas-Hernández, V.E. Abiotic stress in plants and metabolic responses. In Abiotic Stress-Plant Responses and Applications in Agriculture; Vahdati, K., Leslie, C., Eds.; InTech: London, UK, 2013; pp. 25–48. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Jia, W.; Yang, J.; Ismail, A.M. Role of ABA in integrating plant responses to drought and salt stresses. Field Crops Res. 2006, 97, 111–119. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrao, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Ruan, C.J.; Ja, T.D.S. Metabolomics: Creating new potentials for unraveling the mechanisms in response to salt and drought stress and for the biotechnological improvement of xerohalophytes. Crit. Rev. Biotechnol. 2011, 31, 153–169. [Google Scholar] [CrossRef]
- Jalili Marandi, R. Environmental Stress Physiologyand Mechanisms of Resistance in Horticultural Plants; Jahad University Press: Urumie, Iran, 2010; p. 636. [Google Scholar]
- De Magalhães Erismann, N.; Machado, E.C.; Tucci, M.L.S.A. Photosynthetic limitation by CO2 diffusion in drought stressed orange leaves on three rootstocks. Photosynth. Res. 2008, 96, 163–172. [Google Scholar] [CrossRef]
- Mahmoud, A.W.M.; Swaefy, H.M. Comparison between commercial and nano NPK in presence of nano zeolite on sage plant yield and its components under water stress. Agriculture 2020, 66, 24–39. [Google Scholar] [CrossRef]
- Hasani, A.; Beygi, R.O. Effects of water stress on some morphological, physiological, and metabolic characteristics of basil plant. J. Agric. Sci. 2002, 12, 47–59. [Google Scholar]
- Mahmoud, A.W.M.; Samy, M.M.; Sany, H.; Eid, R.R.; Rashad, H.M.; Abdeldaym, E.A. Nanopotassium, nanosilicon, and biochar applications improve potato salt tolerance by modulating photosynthesis, water status, and biochemical constituents. Sustainability 2022, 14, 723. [Google Scholar] [CrossRef]
- Shang, Y.; Hasan, M.; Ahammed, G.J.; Li, M.; Yin, H.; Zhou, J. Applications of nanotechnology in plant growth and crop protection: A review. Molecules 2019, 24, 2558. [Google Scholar] [CrossRef] [Green Version]
- Hassan, A.Z.A.; Mahmoud, A.W.M.; Turky, G.M.; Safwat, G. Rice husk derived biochar as smart material loading nano nutrients and microorganisms. Bulg. J. Agric. Sci. 2020, 26, 309–322. [Google Scholar]
- Adisa, I.O.; Pullagurala, V.L.R.; Peralta-Videa, J.R.; Dimkpa, C.O.; Elmer, W.H.; Gardea-Torresdey, J.; White, J. Recent advances in nano-enabled fertilizers and pesticides: A critical review of mechanisms of action. Environ. Sci. Nano 2019, 6, 2002–2030. [Google Scholar] [CrossRef]
- Manjunatha, S.B.; Biradar, D.P.; Aladakatti, Y.R. Nanotechnology and its applications in agriculture: A review. J. Farm. Sci. 2016, 29, 1–3. [Google Scholar]
- Zulfiqar, F.; Navarro, M.; Ashraf, M.; Akram, N.A.; Munné-Bosch, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Haliem, M.E.F.; Hegazy, H.S.; Hassan, N.S.; Naguib, D.M. Effect of silica ions and nano silica on rice plants under salinity stress. Ecol. Eng. 2017, 99, 282–289. [Google Scholar] [CrossRef]
- Rastogi, A.; Tripathi, D.K.; Yadav, S.; Chauhan, D.K.; Živčák, M.; Ghorbanpour, M.; El-Sheery, N.I.; Brestic, M. Application of silicon nanoparticles in agriculture. 3 Biotech 2019, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Colombani, N.; Mastrocicco, M.; Giambastiani, B.M.S. Predicting salinization trends in a lowland coastal aquifer: Comacchio (Italy). Water Resour. Manag. 2015, 29, 603–618. [Google Scholar] [CrossRef]
- Wen, J.; Dong, H.; Zeng, G. Application of zeolite in removing salinity/sodicity from wastewater: A review of mechanisms, challenges and opportunities. J. Clean. Prod. 2018, 197, 1435–1446. [Google Scholar] [CrossRef]
- Richards, L.S. Diagnosis and Improvement of Saline and Alkaline Soils; Handbook; U.S. Department of Agriculture: Washington, DC, USA, 1954; p. 60.
- Jackson, M.L. Soil Chemical Analysis; Text book; Printice-Hall of India, Privat Limited: New Delhi, India, 1973; Volume 144–197, p. 381. [Google Scholar]
- Karmeli, D.; Keller, J. Trickle Irrigation Design; Rain-Bird Sprinkler Mfg. Co.: Glendora, CA, USA, 1975. [Google Scholar] [CrossRef]
- Stenhagen, S.; Abrahamson, E.; MacLafferty, F.W. Registry of Mass Spectral Data; Wiley & Sons: New York, NY, USA, 1974. [Google Scholar]
- Sadtler, R. The Sadtler Standard Gas Chromatography Retention Index Library; Sadtler Research Laboratories: Philadelphia, PA, USA, 1986; Volume 4. [Google Scholar]
- Kinney, T.J.; Masiello, C.A.; Dugan, B.; Hockaday, W.C.; Dean, M.R.; Zygourakis, K.; Barnes, R.T. Hydrologic properties of biochars produced at different temperatures. Biomass Bioenergy 2012, 41, 34–43. [Google Scholar] [CrossRef]
- Mahmoud, A.W.M.; El-Attar, A.B.; Esmail, S.E.A.; Sany, H. Boosting biochemical composition of stevia plant using biochar loaded with beneficial microorganism strains. Bulg. J. Agric. Sci. 2020, 26, 748–760. [Google Scholar]
- Hassan, A.Z.A.; Mahmoud, A.W.M. Hydrothermal synthesis of nano crystals (AM) zeolite using variable temperature programs. J. Nanomater. Mol. Nanotechnol. 2015, 4, 7–8. [Google Scholar] [CrossRef]
- Li, J.; Wee, C.; Sohn, B. Effect of ammonium- and potassium-loaded zeolite on kale, growth and soil property. Am. J. Plant Sci. 2013, 4, 1976–1982. [Google Scholar] [CrossRef] [Green Version]
- Helrich, K. Official Methods of Analysis of the Association of Official Analytical Chemists; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.W.M.; Abdeldaym, E.A.; Abdelaziz, S.M.; El-Sawy, M.B.I.; Mottaleb, S.A. Synergetic effects of zinc, boron, silicon, and zeolite nanoparticles on confer tolerance in potato plants subjected to salinity. Agronomy 2020, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Lugojan, C.; Ciulca, S. Evaluation of relative water content in winter wheat. Hortic. For. Biotechnol. 2011, 15, 173–177. [Google Scholar]
- Moran, R. Formulae for determination of chlorophyllous pigments extracted with N,N-dimethylformamide. Plant Physiol. 1982, 69, 1376–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.K. Rapid determination of free proline for waterstress studies. Plant Soil 1973, 39, 205–208. [Google Scholar] [CrossRef]
- Sheligel, H.Q. Die verwertungorgngischersourendurch chlorella lincht. Planta J. 1986, 12, 47–51. [Google Scholar]
- Fales, H.M.; Jaouni, T.M.; Babashak, J.F. Simple device for preparing ethereal diazomethane without resorting to codistillation. Anal. Chem. 1973, 45, 2302–2303. [Google Scholar] [CrossRef]
- Vogel, A.I. A Text Book of Practical Organic Chemistry, 3rd ed.; English Language Book Society and Longman Growth Ltd.: London, UK, 1975; pp. 197–596. [Google Scholar]
- Polle, A.; Otter, T.; Mehne-Jakobs, B. Effect of magnesium deficiency on antioxidative systems in needles of Norway spruce [Piceaabies (L.) Karst.] grown with different ratios of nitrate and ammonium as nitrogen sources. New Phytol. 1994, 128, 621–628. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzym. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen-peroxide is scavenged by ascorbate-specific peroxidase in spinach-chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Duncan, P.B. New multiple range and multiple F-tests biometrics. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Mikhak, A.; Sohrabi, A.; Kassaee, M.Z.; Feizian, M. Synthetic nanozeolite/nanohydroxyapatite as a phosphorus fertilizer for German chamomile (Matricaria chamomilla L.). Ind. Crops Prod. 2017, 95, 444–452. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Gulzar, S.; Khan, M.A.; Ungar, I.A. Salt tolerance of a coastal salt marsh grass. Commun. Soil Sci. Plant Anal. 2003, 34, 2595–2605. [Google Scholar] [CrossRef]
- Rengasamy, P.; Olsson, K.A. Irrigation and sodicity. Aust. J. Soil Res. 1993, 31, 821–837. [Google Scholar] [CrossRef]
- Khoshsokhan, F.; Babalar, M.; Chaghazardi, H.R.; Fatahi Moghadam, M.R. Effect of salinity and drought stress on germination in thymeeffect of salinity and drought stress on germination indices of two Thymus species. Cercetări agronomice în Moldova 2012, 1, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Nayyar, H.; Gupta, D. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- Semerci, A.; Cicek, N.; Karahan, F.A.; Özyürek, E.; Arici, Y.K.; Ekmekci, Y. Some growth and chlorophyll fluorescence parameters of black and hybrid poplar clones under water stress. Turk. J. Agric. For. 2017, 41, 348–356. [Google Scholar] [CrossRef]
- Pedroso, F.K.J.V.; Prudente, D.A.; Carolina, A.; Bueno, R.; Machado, E.C.; Ribeiro, R.V. Drought tolerance in citrus trees is enhanced by rootstock-dependent changes in root growth and carbohydrate availability. Environ. Exp. Bot. 2014, 101, 26–35. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T. Diffusive and metabolic limitations to photosynthesis under drought and salinity in c3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Mazher, A.M.A.; El-Quesni, E.M.F.; Farahat, M.M. Responses of ornamental plants and woody trees to salinity. World J. Agric. Sci. 2007, 3, 386–395. [Google Scholar]
- Kohli, S.K.; Handa, N.; Sharma, A.; Kumar, V.; Kaur, P.; Bhardwaj, R. Synergistic effect of 24-epibrassinolide and salicylic acid on photosynthetic efficiency and gene expression in Brassica juncea L. under Pb stress. Turk. J. Biol. 2017, 41, 943–953. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Bielsa, B.; Hewitt, S.; Reyes-Chin-Wo, S.; Dhingra, A.; Rubio-Cabetas, M.J. Identification of water use efficiency related genes in ‘Garnem’ almond-peach rootstock using time-course transcriptome analysis. PLoS ONE 2018, 13, e0205493. [Google Scholar] [CrossRef]
- Rodríguez-Gamir, J.; Primo-Millo, E.; Forner, J.B.; Forner-Giner, M.A. Citrus rootstock responses to water stress. Sci. Hortic. 2010, 126, 95–102. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Thirunavukkarasu, M.; Subramanian, K. Surface modified nano-zeolite used as carrier for slow release of sulphur. J. Nat. Appl. Sci. 2014, 6, 19–26. [Google Scholar] [CrossRef]
- Moreno, J.L.; Ondoño, S.; Torres, I.; Bastida, F. Compost, leonardite, and zeolite impacts on soil microbial community under barley crops. J. Soil Sci. Plant Nutr. 2017, 17, 214–230. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Chi, D.; Xia, G.; Chen, T.; Sun, Y.; Song, Y. Effects of zeolite on drought resistance and water–nitrogen use efficiency in paddy rice. J. Irrig. Drain. Eng. 2019, 145, 04019024. [Google Scholar] [CrossRef]
- De Smedt, C.; Steppe, K.; Spanoghe, P. Beneficial effects of zeolites on plant photosynthesis. Adv. Mater. 2017, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.K.; Singh, V.P.; Kumar, D.; Chauhan, D.K. Impact of exogenous silicon addition on chromium uptake, growth, mineral elements, oxidative stress, antioxidant capacity, and leaf and root structures in rice seedlings exposed to hexavalent chromium. Acta Physiol. Plant. 2012, 34, 279–289. [Google Scholar] [CrossRef]
- Jullok, N.; Van Hooghten, R.; Luis, P.; Volodin, A.; Van Haesendonck, C.; Vermant, J.; Van der Bruggen, B. Effect of silica nanoparticles in mixed matrix membranes for pervaporation dehydration of acetic acid aqueous solution: Plant-inspired dewatering systems. J. Clean. Prod. 2016, 112, 4879–4889. [Google Scholar] [CrossRef]
- Kalteh, M.; Alipour, Z.T.; Ashraf, S.; Aliabadi, M.M.; Nosratabadi, A.F. Effect of silica nanoparticles on basil (Ocimumbasilicum) under salinity stress. J. Chem. Health Risks 2014, 4, 49–55. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon decreases transpiration rate and conductance from stomata of maize plants. J. Plant. Nutr. 2006, 29, 1637–1647. [Google Scholar] [CrossRef]
- DeRosa, M.C.; Monreal, C.; Schnitzer, M.; Walsh, R.; Sultan, Y. Nanotechnology in fertilizers. Nat. Nanotechnol. 2010, 5, 91. [Google Scholar] [CrossRef]
- Xiao, R.; Awasthi, M.K.; Li, R.; Park, J.; Pensky, S.M.; Wang, Q.; Wang, J.J.; Zhang, Z. Recent developments in biochar utilization as an additive in organic solid waste composting: A review. Bioresour. Technol. 2017, 246, 203–213. [Google Scholar] [CrossRef]
- Sánchez-García, M.; Alburquerque, J.A.; Sánchez-Monedero, M.A.; Roig, A.; Cayuela, M.L. Biochar accelerates organic matter degradation and enhances N mineralisation during composting of poultry manure without a relevant impact on gas emissions. Bioresour. Technol. 2015, 192, 272–279. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Wang, M.; Pandey, A.; Chen, H.; Awasthi, S.K.; Wang, Q.; Ren, X.; Lahori, A.H.; Li, D.S.; Li, R.; et al. Heterogeneity of zeolite combined with biochar properties as a function of sewage sludge composting and production of nutrient-rich compost. Waste Manag. 2017, 68, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, M.H.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.S.; Liu, X.; Gao, D.; Clevenger, J.; Dash, S.; et al. The genome sequences of Arachis duranensis and Arachis ipaensis, the diploid ancestors of cultivated peanut. Nat. Genet. 2016, 48, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Demmig-Adams, B.; Jared, J.; Stewart, I.D.; Christopher, R.; Adams, W. Optimization of photosynthetic productivity in contrasting environments by regulons controlling plant form and function. Int. J. Mol. Sci. 2018, 19, 872. [Google Scholar] [CrossRef] [Green Version]
- Cha-Um, S.; Kirdmanee, C. Effect of salt stress on proline accumulation, photosynthetic ability and growth characters in two maize cultivars. Pak. J. Bot. 2009, 41, 87–98. [Google Scholar]
- Turner, N.C.; Abbo, S.; Berger, J.D.; Chaturvedi, S.K.; French, R.J.; Ludwig, C.; Mannur, D.M.; Singh, S.J.; Yadava, H.S. Osmotic adjustment in chickpea (Cicer arietinum L.) results in no yield benefit under terminal drought. J. Exp. Bot. 2007, 58, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Babaee, K.; Dehaghi, M.A.; ModaresSanavi, S.A.M.; Jabbari, R. Water deficit effect on morphology, proline content and thymol percentage of Thyme (Thymus vulgaris L.). Iran. J. Med. Aromat. Plants Res. 2010, 26, 239–251. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Al Hassan, M.; Fuertes, M.M.; Ramos Sánchez, F.J.; Vicente, O.; Boscaiu, M. Effects of salt and water stress on plant growth and on accumulation of osmolytes and antioxidant compounds in cherry tomato. Not. Bot. Horti Agrobot. 2015, 43, 1–11. [Google Scholar] [CrossRef]
- Treml, J.; Šmejkal, K. Flavonoids as potent scavengers of hydroxyl radicals. Compr. Rev. Food Sci. Food Saf. 2016, 15, 720–738. [Google Scholar] [CrossRef]
- Song, A.; Li, P.; Fan, F.; Li, Z.; Liang, Y. The effect of silicon on photosynthesis and expression of its relevant genes in rice (Oryza sativa L.) under high-zinc stress. PLoS ONE 2014, 9, e113782. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Wang, M.; Bi, D. Selenium modulates the activities of antioxidant enzymes, osmotic homeostasis and promotes the growth of sorrel seedlings under salt stress. Plant Growth Regul. 2005, 45, 155–163. [Google Scholar] [CrossRef]
- Abou-Baker, N.H.; Ibrahim, E.A.; Abd-Eladl, M.M. Biozeolite for improving bean production under abiotic stress conditions. Bull. Transilv. Univ. Brasov. For. Wood Ind. Agric. Food Eng. Ser. II 2017, 10, 31–46. [Google Scholar]
- Cetinkaya, H.; Kulak, M.; Karaman, M.; Karaman, H.S.; Kocer, F. Flavonoid accumulation behavior in response to the abiotic stress: Can a uniform mechanism be illustrated for all plants. In Flavonoids—From Biosynthesis to Human Health; Intechopen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, K.; Ito, Y.; Yamaguchi-Shinozaki, K. Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol. 2009, 149, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Wang, B.; Tang, K.; Hsu, C.C.; Xie, S.; Du, H.; Yang, Y.; Tao, W.A.; Zhu, J.K. An Arabidopsis nucleoporin NUP85 modulates plant responses to ABA and salt stress. PLoS Genet. 2017, 13, e1007124. [Google Scholar] [CrossRef] [Green Version]
- Sharma, E.; Sharma, R.; Borah, P.; Jain, M.; Khurana, J.P. Emerging roles of auxin in abiotic stress responses. In Elucidation of Abiotic Stress Signaling in Plants: Functional Genomics Perspectives; Pandey, G.K., Ed.; Springer: New York, NY, USA, 2015; pp. 299–328. [Google Scholar] [CrossRef]
- Song, Y.; Wang, L.; Xiong, L. Comprehensive expression profiling analysis of OsIAA gene family in developmental processes and in response to phytohormone and stress treatments. Planta 2009, 229, 577–591. [Google Scholar] [CrossRef]
- Yuan, H.; Zhao, K.; Lei, H.; Shen, X.; Liu, Y.; Liao, X.; Li, T. Genome-wide analysis of the GH3 family in apple (Malus× domestica). BMC Genom. 2013, 14, 297. [Google Scholar] [CrossRef] [Green Version]
- Korver, R.A.; Koevoets, I.T.; Testerink, C. Out of shape during stress: A key role for auxin. Trends Plant Sci. 2018, 23, 783–793. [Google Scholar] [CrossRef]
- Novák, O.; Napier, R.; Ljung, K. Zooming in on plant hormone analysis: Tissue- and cell-specific approaches. Annu. Rev. Entomol. 2017, 68, 323–348. [Google Scholar] [CrossRef]
- Shan, X.Y.; Yan, J.B.; Xie, D.X. Comparison of phytohormone signaling mechanisms. Curr. Opin. Plant Biol. 2012, 15, 84–91. [Google Scholar] [CrossRef]
- Shu, K.; Chen, Q.; Wu, Y.; Liu, R.; Zhang, H.; Wang, P.; Li, Y.; Wang, S.; Tang, S.; Liu, C.; et al. ABI4 mediates antagonistic effects of abscisic acid and gibberellins at transcript and protein levels. Plant J. 2016, 85, 348–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urano, K.; Maruyama, K.; Jikumaru, Y.; Kamiya, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Analysis of plant hormone profiles in response to moderate dehydration stress. Plant J. 2017, 90, 17–36. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sanchez, F.; Syvertsen, J.P.; Gimeno, V.; Botia, P.; Perez-Perez, J.G. Responses to flooding and drought stress by two citrus rootstock seedlings with different water-use efficiency. Biol. Plant 2007, 130, 532–542. [Google Scholar] [CrossRef]
- Hussain, S.; Khalid, M.F.; Saqib, M.; Ahmad, S.; Zafar, W.; Rao, M.J.; Morillon, R.; Anjum, M.A. Drought tolerance in citrus rootstocks is associated with better antioxidant defense mechanism. Acta Physiol. Plant. 2018, 40, 135. [Google Scholar] [CrossRef]
- Kim, Y.H.; Khan, A.L.; Kim, D.H.; Lee, S.Y.; Kim, K.M.; Waqas, M.; Jung, H.Y.; Shin, J.H.; Kim, J.G.; Lee, I.J. Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, oryza sativa low silicon genes, and endogenous phytohormones. BMC Plant Biol. 2014, 14, 13. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought; salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Lisar, S.Y.S.; Motafakkerazad, R.; Hossain, M.M.; Rahman, I.M.M. Water stress in plants: Causes, effects and responses. In Water Stress; Rahman, M., Hasegawa, H., Eds.; InTech: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.J.; Randall, D.P.; Flowers, T.J. Silicon deposition in the root reduces sodium uptake in rice (Oryza sativa L.) seedlings by reducing bypass flow. Plant Cell Environ. 2006, 29, 1970–1979. [Google Scholar] [CrossRef]
- Abdi, G.; Salehi, H.; Eshghi, S. Effect of natural zeolite and paclobutrazol on reducing salt stress in Kentucky bluegrass (Poa pratensis L.). Hortic. Environ. Biotechnol. 2010, 51, 159–166. [Google Scholar]
- Morales-Díaz, A.B.; Ortega-Ortíz, H.; Juárez-Maldonado, A.; Cadenas-Pliego, G.; González-Morales, S.; Benavides-Mendoza, A. Application of nanoelements in plant nutrition and its impact in ecosystems. Adv. Nat. Sci. Nanosci. Nanotechnol. 2017, 8, 013001. [Google Scholar] [CrossRef]
- Mahmoud, A.W.M.; Abdelaziz, S.M.; El-mogy, M.M.; Abdeldaym, E.A. Effect of foliar ZnO and FeOnanoparticles application on growth and nutritional quality of red radish and assessment of their accumulation on human health. Agriculture 2019, 65, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Mehrabanjoubani, P.; Abdolzadeh, A.; Sadeghipour, H.R.; Aghdasi, M. Impacts of silicon nutrition on growth and nutrient status of rice plants grown under varying zinc regimes. Theor. Exp. Plant Physiol. 2015, 27, 19–29. [Google Scholar] [CrossRef]
- Bolanos, L.; Lukaszewski, K.; Bonilla, I.; Blevins, D. Why boron? Plant Physiol. Biochem. 2004, 42, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.C.; Ding, R.X.; Liu, Q. Effects of silicon on salt tolerance of barley and its mechanism. Sci. Agric. Sin. 1999, 32, 75–83. [Google Scholar]
- Noori, M.; Zendehdel, M.; Ahmadi, A. Using natural zeolite for the improvement of soil salinity and crop yield. Toxicol. Environ. Chem. 2006, 88, 77–84. [Google Scholar] [CrossRef]
- Al-Busaidi, A.; Yamamoto, T.; Inoue, M.; Eneji, A.E.; Mori, Y.; Irshad, M. Effects of zeolite on soil nutrients and growth of barley following irrigation with saline water. J. Plant. Nutr. 2008, 31, 1159–1173. [Google Scholar] [CrossRef]
- Kulevanova, S.; Ristić, M.; Stafilov, T.; Dorevski, K.; Ristov, T. Essential oil analysis of some taxa of genera Thymus L.—Environment influences. Bull. Chem. Technol. Maced. 1996, 15, 33–38. [Google Scholar]
- Hay, R.K.M. Phenology and ontogeny. In Volatile Oil Crops: Their Biology, Biochemistry and Production; Longman Scientific and Technical: London, UK, 1993. [Google Scholar]
- Farahani, H.A.; Valadabadi, S.A.; Daneshian, J.; Khalvati, M.A. Evaluation changing of essential oil of balm (Melissa officinalis L.) under water deficit stress conditions. J. Med. Plants Res. 2009, 3, 329–333. [Google Scholar]
- Bahreininejad, B.; Razmjoo, J.; Mirza, M. Effect of water stress on productivity and essential oil content and composition of Thymus carmanicus. J. Essent. Oil Bear. Plants 2014, 17, 717–725. [Google Scholar] [CrossRef]
- Leicach, S.R.; Garau, A.M.; Guarnaschelli, A.B.; Yaber Grass, M.A.; Sztarker, N.D.; Dato, A. Changes in Eucalyptus camaldulensis essential oil composition as response to drought preconditioning. J. Plant Interact. 2010, 5, 205–210. [Google Scholar] [CrossRef]
- Ozturk, A.; Unlukara, A.; Ipek, A.; Gurbuz, B. Effects of salt stress and water deficit on plant growth and essential oil content of lemon balm (Melissa officinalis L.). Pak. J. Bot. 2004, 36, 787–792. [Google Scholar]
- Sharafzadeh, S.; Zare, M. Effect of drought stress on qualitative and quantitative characteristics of some medicinal plants from Lamiaceae family: A review. Adv. Environ. Biol. 2011, 5, 2058–2062. [Google Scholar]



{kind=link}
{kind=link}
| Parameters | Soil Depth [cm] | |
|---|---|---|
| 0–30 | 30–60 | |
| Particle-size distribution [%] | ||
| Sand | 90.10 | 90.00 |
| Silt | 6.90 | 6.50 |
| Clay | 3.00 | 3.50 |
| Textural class | Sand | Sand |
| Saturation water content [cm3 cm−3] | 0.385 | 0.396 |
| Field capacity [cm3cm−3] | 0.213 | 0.218 |
| Permanent wilting point [cm3cm−3] | 0.057 | 0.057 |
| Available water [cm3.cm−3] | 0.156 | 0.161 |
| Bulk density [mg m−3] | 1.64 | 1.65 |
| Saturated hydraulic conductivity, [m day−1] | 2.40 | 2.34 |
| Organic matter [%] | 0.31 | 0.25 |
| Calcium carbonates [%] | 4.80 | 3.71 |
| pH (1:1 soil: water suspension) | 7.70 | 7.81 |
| EC(1:1 soil: water extract) [dSm−1] | 4.02 | 4.13 |
| Soluble ions [meq/100] | ||
| Ca2+ | 13.85 | 13.41 |
| Mg2+ | 12.15 | 10.59 |
| Na+ | 8.10 | 10.25 |
| K+ | 6.00 | 6.05 |
| CO32− | - | - |
| HCO3− | 11.92 | 9.75 |
| Cl− | 14.00 | 10.50 |
| SO42− | 15.08 | 21.30 |
| Available nutrients [mg Kg−1 soil] | ||
| N | 16.21 | 13.12 |
| P | 7.78 | 6.21 |
| K | 46.50 | 45.89 |
| Fe | 9.20 | 12.00 |
| Mn | 1.63 | 1.50 |
| Cu | 2.10 | 1.15 |
| Zn | 2.00 | 1.61 |
| B | 0.23 | 0.21 |
| Property | Value |
|---|---|
| Moisture content [%] | 25 |
| pH [1:5] | 7.5 |
| EC (1:5 extract) [dS m−1] | 3.1 |
| Organic matter [%] | 70 |
| Organic-C [%] | 33.11 |
| Total-N [%] | 1.82 |
| C/N ratio | 14:1 |
| Total-K [%] | 1.25 |
| Total-P [%] | 1.29 |
| Fe [ppm] | 1019 |
| Mn [ppm] | 111 |
| Cu [ppm] | 180 |
| Zn [ppm] | 280 |
| Total content of Bacteria [CFU.g−1] | 2.5 × 107 |
| Phosphate-dissolving Bacteria [CFU.g−1] | 2.5 × 106 |
| Weed seeds | 0 |
| Property | Rice Husk-Derived Biochar |
|---|---|
| Si (mg/kg) | 179 |
| Ca (mg/kg) | 213 |
| K (mg/kg) | 199 |
| Mg (mg/kg) | 179 |
| Moisture content (%) | 3.88 |
| Ash (%) | 47.90 |
| pH | 7.65 |
| Fixed C (mg/kg) | 46.35 |
| H (mg/kg) | 2.64 |
| N (mg/kg) as N2O after inoculation | 2.4 |
| N (mg/kg) after soaking in ammonium sulfate | 3.65 |
| Sulfate (mg/kg) | 0.22 |
| Oxygen (mg/kg) | 2.74 |
| H:C | 0.05 |
| C:N (after inoculation by a microorganism) | 18.85 |
| C:N (after soaking in ammonium sulfate) | 12.92 |
| EC (dS/m) | 0.14 |
| Zeta potential (mV) | −26.6 |
| Chemical Composition (%) | SiO2 | TiO2 | Al2O3 | Fe2O3 | FeO | MnO | MgO | CaO | Na2O | K2O | SrO | P2O3 | N |
| 45.50 | 2.81 | 13.30 | 5.40 | 8.31 | 0.51 | 6.30 | 9.52 | 2.83 | 0.87 | 0.22 | 0.67 | 2.70 | |
| Trace Elements (ppm) | Ba | Co | Cr | Se | Cu | Zn | Zr | Nb | Ni | Rb | Y | ||
| 10 | 1.2 | 35 | 0.8 | 19 | 64 | 257 | 13 | 55 | 15 | 22 |
| Treatment | Plant Height (cm) | Lateral Branches Number/Plant | Shoot F.W. (g.) | Shoot D.W. (g.) | Root F.W. (g.) | Root D.W. (g.) | YieldF.W. (kg\fed.) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | |
| T1 (NPK+ O) | 17.7 e | 18.8 d | 13.0 c | 14.7 d | 34.5 d | 35.8 d | 10.6 d | 11.5 e | 23.5 d | 25.0 d | 8.7 f | 10.6 e | 418.5 e | 442.5 e |
| T2(NPK+nS+O) | 19.8 d | 20.3 c | 15.2 c | 18.2 c | 36.8 c | 38.6 c | 12.8 c | 13.5 d | 26.8 c | 28.8 c | 12.5 d | 15.2 c | 521.1 d | 561.6 d |
| T3 (BC+O) | 19.5 d | 18.8 d | 14.9 c | 16.6 c | 36.1 c | 38.1 c | 12.1 c | 12.7 d | 25.6 d | 26.2 d | 10.8 e | 12.4 d | 473.6 e | 500.7 d |
| T4 (nS+O) | 18.6 d | 17.8 d | 17.8 b | 20.0 b | 37.5 c | 39.2 c | 12.1 c | 13.5 d | 28.1 c | 29.6 c | 15.1 c | 15.4 c | 548.0 d | 577.3 d |
| T5(nZ+nS+O) | 23.1 b | 25.7 b | 20.5 a | 23.3 b | 43.8 b | 45.2 b | 16.2 a | 16.6 b | 30.1 b | 33.5 b | 17.5 b | 18.1 b | 708.5 b | 783.2 b |
| T6(BC+nS +O) | 21.2 c | 23.0 c | 18.8 b | 21.5 b | 40.5 b | 43.7 b | 14.2 b | 15.0 c | 28.7 c | 30.3 c | 15.3 c | 16.7 c | 670.4 c | 695.8 c |
| T7(nZ+BC+nS+O) | 25.6 a | 29.4 a | 22.4 a | 25.5 a | 47.3 a | 50.5 a | 17.8 a | 19.7 a | 35.6 a | 38.3 a | 19.4 a | 21.3 a | 875.3 a | 921.5 a |
| Treatment | Leaf Water Content (%) | Photosynthesis Rate (µmol m−2s−1) | Intercellular CO2 Concentration (ppm) | Transpiration Rate (mmol m−2s−1) | Water Use Efficiency (μmol mmol−1) | Oil Yield (L\fed.) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | |
| T1 (NPK+O) | 65.96 c | 66.11 b | 15.87 d | 17.82 d | 92.5 f | 89.6 g | 4.39 a | 5.11 a | 3.61 d | 3.48 d | 2.3 d | 3.6 d |
| T2(NPK+nS+O) | 67.55 b | 67.32 b | 17.08 d | 19.13 c | 98.3e | 100.2 f | 3.65 b | 3.72 c | 4.67 c | 5.14 b | 4.8 c | 5.4 c |
| T3(BC+O) | 66.78 c | 66.13 b | 18.25 c | 20.16 b | 112.5 d | 114.1 e | 4.17 a | 4.52 b | 4.37 c | 4.46 c | 4.3 c | 5.0 c |
| T4 (nS+O) | 67.95 b | 66.51 b | 18.29 c | 20.07 b | 115.3 d | 118.5 d | 3.38 b | 3.72 c | 5.41 b | 5.39 b | 4.5 c | 5.0 c |
| T5(nZ+nS+O) | 70.89 b | 70.12 a | 19.48 b | 21.14 b | 126.7 b | 130.25 b | 3.26 b | 3.29 c | 5.97 b | 6.42 a | 5.6 b | 6.1 b |
| T6(BC+nS+O) | 68.81 b | 66.93 b | 19.09 b | 20.15 b | 119.4 c | 122.6 c | 4.11 a | 4.17 b | 4.64 c | 4.83 c | 5.2 b | 5.8 c |
| T7(nZ+BC+nS+O) | 74.53 a | 71.31 a | 21.61 a | 23.11 a | 131.5 a | 136.8 a | 3.12 b | 3.14 c | 6.92 a | 7.35 a | 6.4 a | 7.3 a |
| Treatment | Total Chlorophyll (mg/g FW.) | Carotenoids (mg/g FW.) | Soluble Sugar (mg/g DW.) | Total Phenols (mg Catechin/g DW.) | Total Flavonoids (μg CE/g.) | Proline Content Leaves (mg g−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | |
| T1 (NPK+O) | 3.01 c | 3.14 d | 2.87 c | 3.05 b | 66.42 a | 68.37 a | 36.2 b | 37.2 c | 0.31 c | 0.33 c | 3.89 c | 3.95 c |
| T2(NPK+nS+O) | 3.71 c | 4.08 b | 3.58 b | 3.66 b | 55.11 b | 54.83 c | 38.1 b | 40.3 b | 0.35 b | 0.37 b | 4.22 b | 4.25 b |
| T3 (BC+O) | 3.15 c | 3.42 c | 3.06 b | 3.28 b | 61.25 a | 62.04 b | 36.8 b | 38.5 b | 0.33 c | 0.36 b | 4.06 b | 4.12 b |
| T4 (nS+O) | 4.11 b | 4.47 b | 3.56 b | 3.72 b | 50.18 c | 51.09 c | 37.0 b | 38.7 b | 0.38 b | 0.37 b | 4.51 b | 4.58 b |
| T5(nZ+nS+O) | 5.03 a | 5.18 a | 4.25 a | 4.62 a | 47.31 d | 58.45 b | 40.3 a | 41.7 b | 0.43 a | 0.42 a | 5.09 a | 5.13 a |
| T6(BC+nS+O) | 4.24 b | 4.50 b | 4.11 a | 4.31 a | 51.36 c | 52.26 c | 37.2 b | 39.5 b | 0.41 a | 0.40 a | 4.68 b | 4.77 b |
| T7(nZ+BC+nS+O) | 5.21 a | 5.32 a | 4.29 a | 4.68 a | 43.69 d | 45.81 d | 42.5 a | 44.7 a | 0.45 a | 0.44 a | 5.36 a | 5.12 a |
| Treatment | IAA (µg/g F.W.) | GA3 (µg/g F.W.) | ABA (µg/g F.W.) | Peroxidase (Unit/mg Protein) | Catalase (Unit/mg Protein) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | |
| T1 (NPK+O) | 15.272 d | 14.829 d | 45.717 c | 48.096 c | 14.019 a | 14.704 a | 0.118 a | 0.124 a | 57.46 a | 56.96 a |
| T2 (NPK+nS+O) | 17.309 c | 15.616 d | 52.801 b | 51.903 c | 13.022 a | 10.801 c | 0.061 b | 0.060 b | 48.81 b | 50.39 b |
| T3 (BC+O) | 18.371 c | 18.552 c | 53.078 b | 49.661 c | 11.077 b | 10.969 c | 0.109 a | 0.116 a | 54.27 a | 56.18 a |
| T4 (nS+O) | 20.771 b | 22.407 b | 55.802 b | 59.470 b | 13.791 a | 12.303 b | 0.043 c | 0.045 c | 38.28 c | 40.08 c |
| T5 (nZ+nS+O) | 26.113 a | 27.491 a | 61.507 a | 66.418 a | 9.460 c | 9.710 c | 0.033 d | 0.030 d | 27.84 d | 30.11 d |
| T6 (BC+nS+O) | 22.758 b | 22.918 b | 57.416 b | 60.382 b | 11.502 b | 11.779 b | 0.042 c | 0.040 c | 37.61 c | 36.44 c |
| T7 (nZ+BC+nS+O) | 26.417 a | 28.682 a | 65.827 a | 68.339 a | 8.413 c | 8.863 d | 0.025 e | 0.023 e | 22.53 e | 20.71 e |
| Treatment | N (%) | P (%) | K (%) | Ca (%) | Mg (%) | Na (%) | Zn (ppm) | Fe (ppm) | Mn (ppm) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | |
| T1 | 1.05 d | 1.13 e | 0.19 e | 0.22 e | 2.18 d | 2.20 d | 1.33 e | 1.35 e | 0.27 d | 0.30 d | 1.75 a | 1.82 a | 15.41 d | 16.05 c | 146.7 d | 148.5 d | 12.57 d | 14.19 c |
| T2 | 1.18 c | 1.26 d | 0.25 c | 0.29 c | 2.23 b | 2.26 c | 1.45 d | 1.49 d | 0.31 c | 0.33 c | 1.34 c | 1.51 c | 17.88 c | 17.92 c | 151.2 c | 150.3 d | 13.17 c | 15.05 b |
| T3 | 1.17 c | 1.25 d | 0.20 d | 0.24 d | 2.20 c | 2.22 d | 1.46 d | 1.48 d | 0.28 d | 0.31 c | 1.62 b | 1.66 b | 17.05 c | 15.14 d | 153.5 c | 155.4 c | 13.06 c | 14.11 c |
| T4 | 1.20 b | 1.54 c | 0.25 c | 0.28 c | 2.22 c | 2.25 c | 1.45 d | 1.51 c | 0.33 c | 0.36 b | 0.70 d | 0.75 d | 18.87 b | 20.11 b | 160.3 b | 164.6 b | 14.56 c | 16.27 b |
| T5 | 1.35 b | 1.92 b | 0.31 b | 0.35 b | 2.25 b | 2.31 b | 1.48 c | 1.53 c | 0.37 b | 0.40 a | 0.59 e | 0.63 e | 22.06 a | 23.10 a | 170.1 a | 173.4 a | 17.53 a | 18.21 a |
| T6 | 1.23 b | 1.56 c | 0.28 c | 0.32 b | 2.24 b | 2.29 b | 1.50 b | 1.59 b | 0.35 c | 0.38 b | 0.71 d | 0.76 d | 19.12 b | 21.15 b | 165.5 b | 167.8 b | 16.44 b | 18.15 a |
| T7 | 2.11 a | 2.50 a | 0.46 a | 0.49 a | 2.39 a | 2.45 a | 1.66 a | 1.78 a | 0.39 a | 0.42 a | 0.42 f | 0.55 f | 22.51 a | 25.31 a | 174.6 a | 177.3 a | 18.78 a | 18.92 a |
| Treatment | T1 | T2 | T3 | T4 | T5 | T6 | T7 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | |
| Hydrocarbons | ||||||||||||||
| Cineol | 0.51 | 0.46 | 0.38 | 0.34 | 0.37 | 0.41 | 0.62 | 0.57 | 0.33 | 0.36 | 0.36 | 0.32 | 0.35 | 0.33 |
| Limonene | 18.07 | 15.39 | 20.41 | 19.22 | 21.16 | 19.44 | 16.73 | 17.29 | 22.18 | 24.62 | 22.04 | 23.71 | 25.41 | 27.33 |
| α-Pinene | 1.14 | 1.09 | 1.32 | 1.20 | 1.11 | 1.35 | 1.15 | 1.11 | 1.41 | 1.40 | 1.32 | 1.36 | 1.51 | 1.55 |
| ß-Pinene | 1.19 | 1.16 | 1.05 | 0.97 | 0.52 | 0.61 | 0.33 | 0.30 | 0.64 | 0.70 | 0.32 | 0.29 | 0.85 | 0.81 |
| Total | 20.91 | 18.10 | 23.16 | 21.73 | 23.16 | 21.81 | 18.83 | 19.27 | 24.56 | 27.08 | 24.04 | 25.68 | 28.12 | 30.02 |
| Oxygenated compound | ||||||||||||||
| Linalool | 1.64 | 1.7 | 1.24 | 1.22 | 0.72 | 0.77 | 0.68 | 0.65 | 1.71 | 1.69 | 1.45 | 1.37 | 1.88 | 2.09 |
| Boreniol | 0.24 | 0.27 | 0.33 | 0.39 | 0.26 | 0.30 | 0.28 | 0.36 | 0.73 | 0.85 | 1.01 | 0.87 | 1.28 | 1.34 |
| Methyl-chavicol | 2.11 | 2.63 | 2.22 | 2.37 | 4.02 | 2.86 | 4.11 | 4.16 | 2.54 | 2.61 | 3.10 | 2.55 | 2.78 | 2.74 |
| Thymol | 32.71 | 35.11 | 39.08 | 41.13 | 37.26 | 40.51 | 26.88 | 29.65 | 42.31 | 41.14 | 37.28 | 39.48 | 50.89 | 53.27 |
| α-Terpineole | 0.33 | 0.38 | 0.49 | 0.52 | 2.19 | 2.43 | 1.03 | 1.11 | 0.89 | 0.82 | 1.12 | 1.16 | 1.09 | 1.18 |
| Carvacrol | 0.81 | 0.92 | 0.88 | 0.97 | 1.02 | 1.11 | 1.27 | 1.31 | 1.42 | 1.45 | 1.14 | 1.17 | 1.79 | 1.83 |
| Total | 37.84 | 41.01 | 44.24 | 46.6 | 45.47 | 47.98 | 34.25 | 37.24 | 49.60 | 48.56 | 45.10 | 46.06 | 59.71 | 62.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoud, A.W.M.; Esmail, S.E.A.; El-Attar, A.B.; Othman, E.Z.; El-Bahbohy, R.M. Prospective Practice for Compound Stress Tolerance in Thyme Plants Using Nanoparticles and Biochar for Photosynthesis and Biochemical Ingredient Stability. Agronomy 2022, 12, 1069. https://doi.org/10.3390/agronomy12051069
Mahmoud AWM, Esmail SEA, El-Attar AB, Othman EZ, El-Bahbohy RM. Prospective Practice for Compound Stress Tolerance in Thyme Plants Using Nanoparticles and Biochar for Photosynthesis and Biochemical Ingredient Stability. Agronomy. 2022; 12(5):1069. https://doi.org/10.3390/agronomy12051069
Chicago/Turabian StyleMahmoud, Abdel Wahab M., Sanaa E. A. Esmail, Asmaa B. El-Attar, Eman Z. Othman, and Reham M. El-Bahbohy. 2022. "Prospective Practice for Compound Stress Tolerance in Thyme Plants Using Nanoparticles and Biochar for Photosynthesis and Biochemical Ingredient Stability" Agronomy 12, no. 5: 1069. https://doi.org/10.3390/agronomy12051069